
大穗型小麦叶片性状、养分含量及氮素分配特征
2013-09-07 07:23王丽芳王德轩上官周平
生态学报 2013年17期
王丽芳,王德轩,上官周平
(西北农林科技大学水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,杨陵 712100)
随着我国小麦需求量的持续增长和耕地面积的不断减少,大穗型小麦品种在农田作物栽培与育种实践中日益受到重视[1]。众多植物表现性状中,叶片性状特征直接影响到植物的生长发育状况,且小麦叶片是与环境接触面积较大的器官,又是籽粒碳水化合物的主要来源,不同的叶片结构性状对产量的影响不一,作为形状和生理特征占优势的大穗型小麦综合有机体,揭示大穗小麦对营养元素资源的利用和对环境的适应策略,探讨大穗小麦叶片结构性状、养分含量与各器官氮素的吸收和累积及其向籽粒的输运特性,对提高大穗小麦产量、改善小麦品质和提高氮素利用效率具有积极意义。
作物比叶面积(SLA)是评价生长发育与高产育种的重要指标[2],能反映植物对碳获取与利用的平衡关系,低的SLA植物具有较好的资源保留能力、适应于资源匮乏的生境条件[3],叶干物质含量(LDMC)主要反映植物养分元素保有能力,植物的SLA和LDMC等性状具体表现为植物的水分利用效率和氮利用效率之间的权衡[4]。在小麦叶片结构性状研究中,Watson[5]指出叶片大小是导致产量差异的一个重要因素,旗叶面积应作为育种的主要目标之一。傅兆麟等[6]研究表明,旗叶面积与穗粒重具有极显著正相关关系,且开花后绿叶面积的大小影响籽粒的灌浆速率[7]。小麦开花前营养体的氮养分调运是籽粒氮的主要来源[8],叶片的结构性状和养分组成可以反映植物对环境条件的适应性,并随土壤和气候条件而变化[9],叶片较大的SLA可能暗示植物具有较高的N、P、K含量[10]。遗传特性和栽培条件尤其是施用氮肥是影响小麦对氮素吸收、累积及转运的关键因素[11-12]。目前针对大穗型小麦叶片研究单集中在叶片面积大小和产量性状方面较多,然而关于大穗型小麦叶片性状与养分含量和氮素转运的研究亟待加强。
本试验选用具有较大产量潜力的8个大穗型小麦新品系和多穗型对照品种——西农979,研究大穗型小麦新品系和多穗型品种的叶性状与养分含量及氮素转运的差异性,为大穗型小麦品种群体的合理构建和养分利用的优化调控提供理论依据。
1 材料与方法
1.1 试验设计
试验于2010—2011年在陕西省武功县苏坊镇苏东村进行,该试验区属暖温带半湿润季风气候区。北纬34°17',经度108°04',海拔577 m,年日照1887.8h,年平均气温13.2℃,年蒸发量1302.5 mm,降水量630 mm,主要集中在7—9月份。土壤为红油土,土层深厚,地势平坦,地力水平较高,保水保肥性好。
试验于2010年10月7—10日播种,播种量为187.5 kg/hm2,平均行距20 cm。试验设置9个处理,小区面积为20 m2,重复3次,2011年6月15日收获。小麦播种时,基肥施尿素375 kg/hm2、磷酸二铵525 kg/hm2、硫酸钾112.5 kg/hm2,冬灌施尿素112.5 kg/hm2,其他栽培管理措施同当地大田,无追肥。
1.2 试验材料
大田试验选用参加2009年陕西省小麦品种预备试验、2010年小麦区域试验和其他性状表现良好的8个具有高产潜力的新品系,是从多种类型中经多年多代筛选的、有望在优质高产上有所突破的一批大穗型小麦新品系(表1)。以黄准冬麦区大面积推广种植的西农979作为对照品种。
1.3 测定项目与方法
1.3.1 比叶面积与干物质含量测定
小麦抽穗期、灌浆前期和灌浆中期选取生长良好、无病虫害且完整的旗叶,每小区选9个叶片,叶片装入自封袋后立即带回实验室,用BenQ Scabber5560 series#4扫描仪扫描,利用图像分析软件(Motic images advanced 3.0)测定其叶面积,然后将叶片挂好标签后完全置于装水的大培养皿中,在暗光下浸泡24h进行复水,至饱和恒重,将饱和的叶片用纸擦至叶表面无多余水分滴落后称重;最后将饱和后的叶片装入信封并在70℃下烘干至恒重(干重,mg)。计算比叶面积(SLA,m2/kg)=叶片面积/叶片干重,叶干物质含量(LDMC,mg/g)= 叶片干重/叶片饱和鲜重[13]。
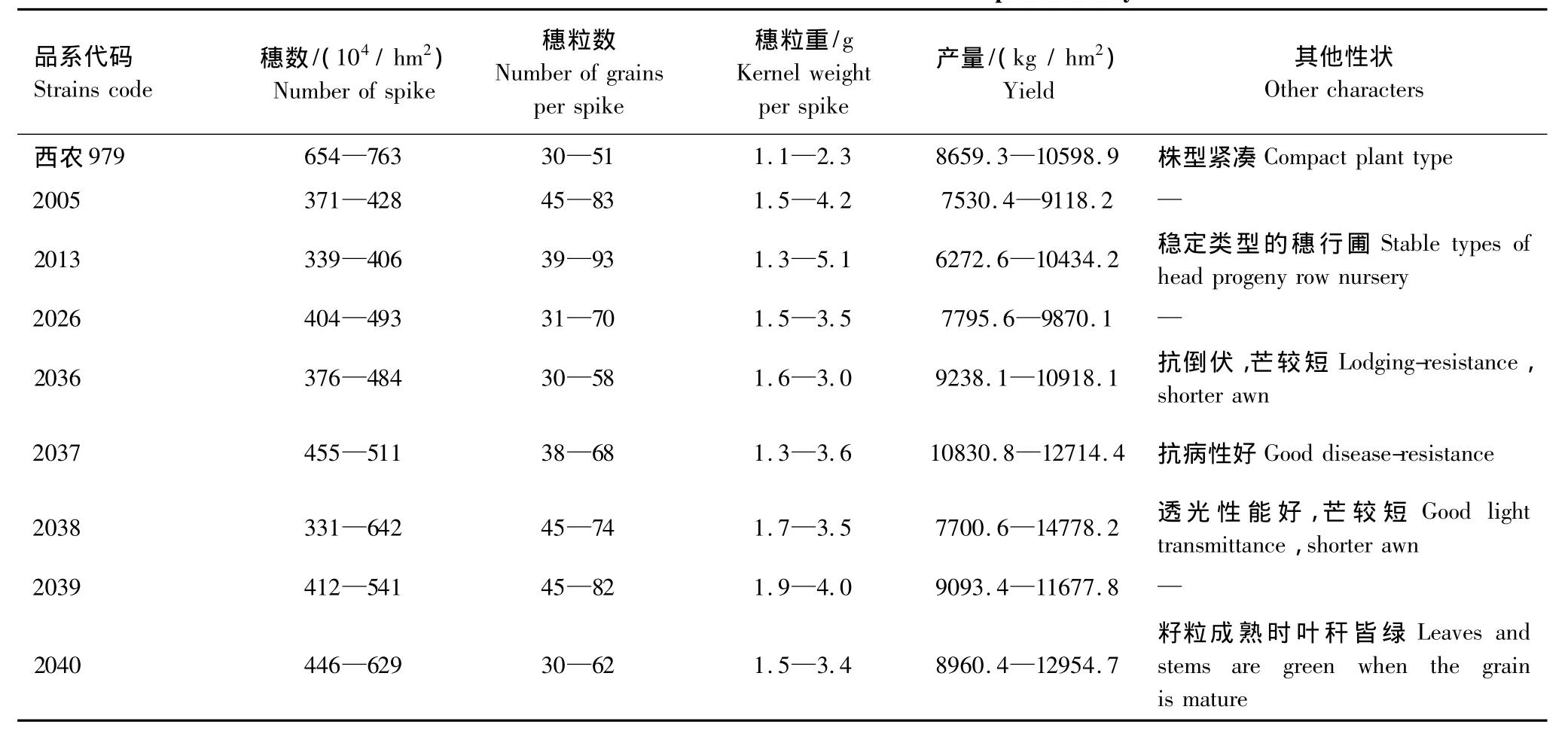
表1 供试小麦品系的主要性状特征Table1 Main characteristics of wheat lines used in present study
1.3.2 叶片养分含量测定
将小麦拔节期、抽穗期和灌浆期采集的叶片样品于105℃下杀青10min,70℃下烘干至恒重,叶片粉碎后过100目筛制成供试样品。植物样品经浓硫酸-过氧化氢溶液消煮后的消煮液,用于植物全氮(N)、全磷(P)、全钾(K)的测定[14]。全氮N用Kjeldahl定氮法(2300全自动定氮仪,Sweden)测定;全磷(P)用钼锑钪比色法(6505紫外分光光度计,UK)测定;全钾(K)用火焰光度法 (ZL-5100原子吸收分光光度计,USA)测定。叶片养分含量单位为质量百分含量,每一样品养分指标的测定重复3次。
1.3.3 氮素分配及转运的采样与测定
于小麦开花期按旗叶、倒二叶、余叶、茎+叶鞘和穗分解,成熟期按旗叶、倒二叶、余叶、茎+叶鞘、穗草(穗轴+颖壳)和籽粒分解,在105℃下杀青10min,在70℃烘至恒重并称重。植物样品粉碎过100目筛制成供试样品,采用H2SO4-H2O2消煮,用Kjeldahl定氮法(2300全自动定氮仪,Sweden)测定氮素含量。参考赵满兴等[15]公式计算小麦氮素累积和转移等指标:
各器官的氮素分配比例(%)=各器官的氮素积累量/单茎氮素含量×100
营养器官氮素转移量=开花期营养器官氮素积累量-成熟期营养器官氮素积累量
营养器官氮素转移率(%)=营养器官氮素转移量/开花期营养器官氮素积累量×100
营养器官转移氮素对籽粒氮素的贡献率(%)=营养器官氮素转移量/成熟期籽粒氮素积累量×100
1.3.3 数据分析
采用Excel 2003进行数据处理,DPS7.05进行统计分析,Duncan's新复极差法进行多重比较。
2 结果与分析
2.1 小麦不同生育期旗叶比叶面积(SLA)的变化
小麦不同生育期、不同品系的SLA不同,同一品种SLA在不同生育期的变化范围也各不相同(表2),8个品系和对照品种的 SLA 变化幅度从大到小依次为2005、2039、2037、2040、2026、CK、2036、2013、2038。对于不同小麦品种同一生育期比较得知,抽穗期叶片SLA以2013品系最大,灌浆期前期以2005品系最大,而灌浆中期以CK品种最大。各生育期大穗型小麦品系平均值均小于西农979,2040品系平均较小的SLA具有较好的资源保留能力,2013品系平均较大的SLA表明其具有较高的资源获取能力。总之,在相同管理条件下,不同小麦品种在同一生育期内SLA均存在明显差异,其叶片性状特征反映了小麦不同品种对生境异质性具有不同的适应对策,大穗型小麦品系小的SLA值表明其可能会更好的适应资源贫瘠和干旱的生境条件。
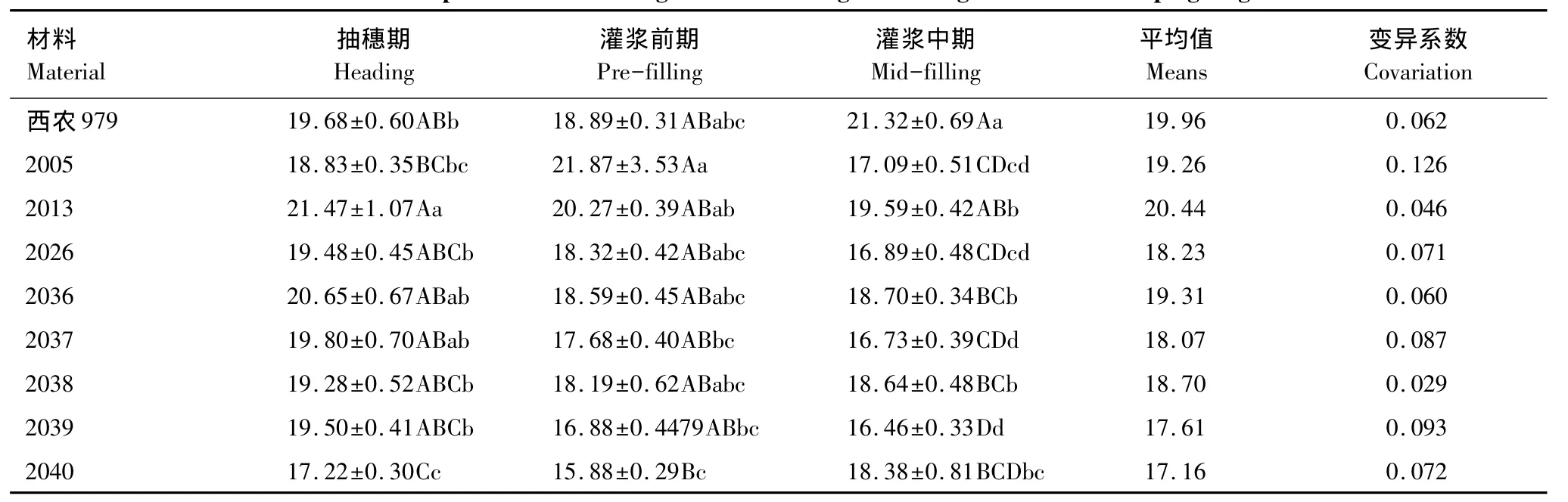
表2 小麦不同生育期旗叶SLA比较/(m2/kg)Table2 Comparison of wheat flag leaf SLA during different growth and developing stages
2.2 小麦不同生育期旗叶干物质含量(LDMC)的变化
各小麦品系(种)LDMC在生育期内存在差异,LDMC和SLA变化趋势基本一致,LDMC在小麦抽穗期后增大(表3)。2040品系平均LDMC较大,2013品系的LDMC较小,而2040品系平均SLA较小,2013品系的SLA 较大。LDMC 的变化幅度从大到小依次为 2039、2036、2037、2038、2013、CK、2040、2026、2005。小麦不同品系LDMC在不同的生育时期变化有所不同,LDMC随SLA减小而变大,表明小麦不同生长发育阶段,各品系对生境条件具有一定的适应性。
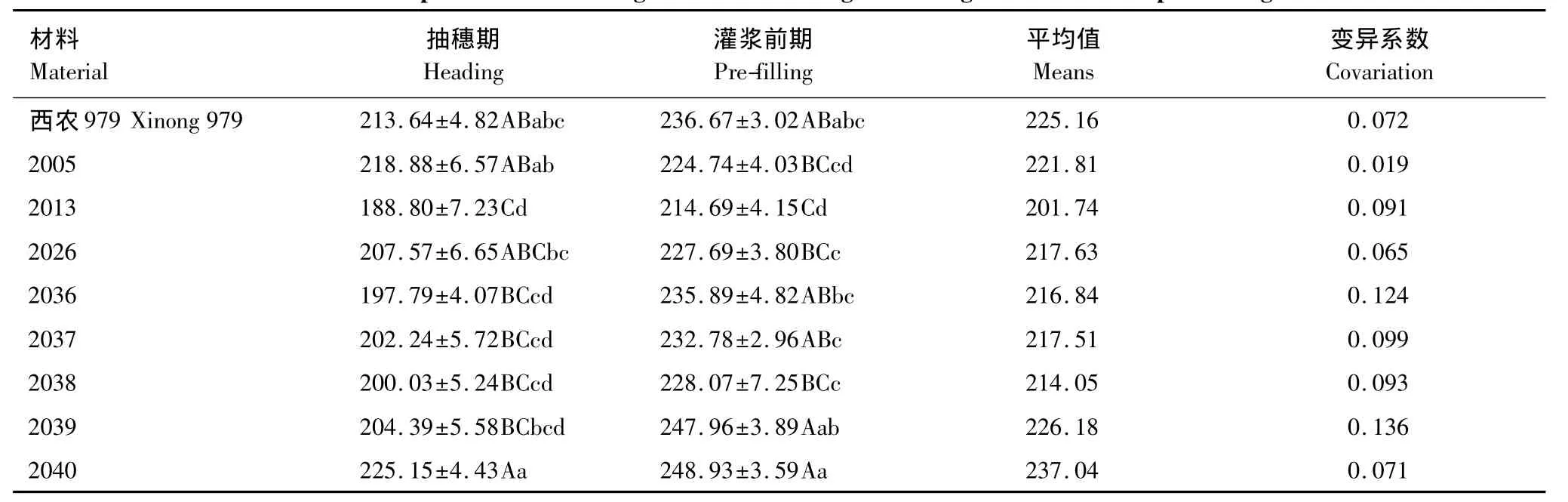
表3 小麦不同生育期旗叶LDMC比较/(mg/g)Table3 Comparison of wheat flag leaf LDMC during different growth and development stages
2.3 小麦不同生育期叶片营养元素平均含量的变化
供试8个小麦新品系和对照品种叶片营养元素含量在拔节期表现差异显著性不同,2013、2037、2040品系的叶片N含量与对照相比差异显著,2005、2013和2036品系P含量高于对照,新品系K含量都高于对照,2037品系与对照N/K差异达极显著水平(表4)。大穗型小麦叶片养分含量表现为N>4.05%、P>0.14%、K>2.25、N/P>17.61、N/K>1.03,拔节期2013品系的 N、P含量分别高于对照13.7%、30%,2037的 K含量高于对照达95%,2040 N/P高于对照26.9%。叶片养分含量大小比较为N>K>P,在小麦拔节期各个新品系表现出了异于对照品种的养分含量。
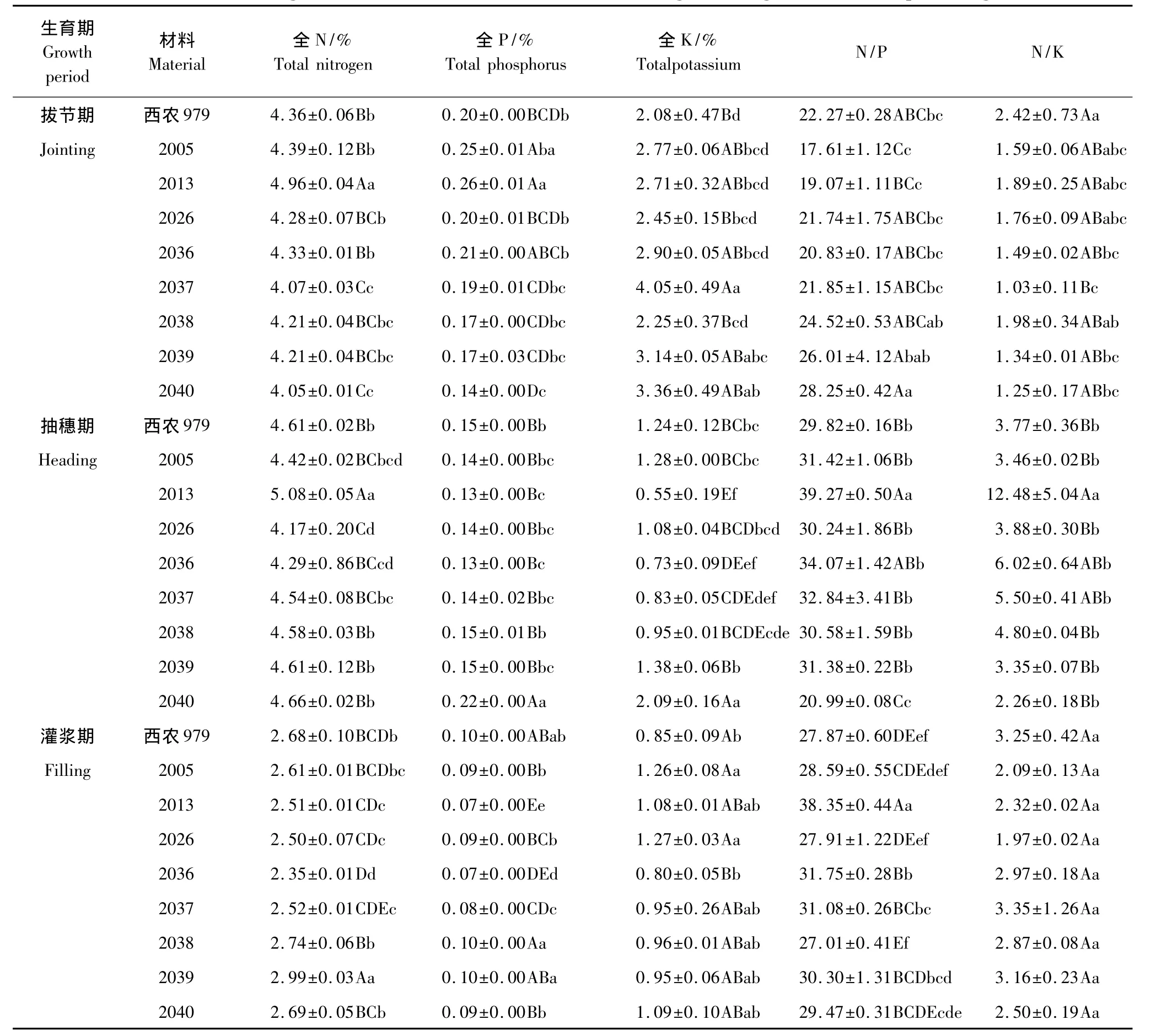
表4 小麦不同生育期叶片营养元素含量变化Table4 The changes of wheat leaf nutrition element contents during different growth and development stages
在小麦抽穗期叶片N含量高于拔节期,除2040品系外其他品系(种)P、K含量下降(表4)。2013品系的N含量、N/P、N/K显著高于对照品种,2040品系的P、K含量显著高于对照品种,其P含量由拔节期所有参试材料中的较小值变为较大值,变化明显。
小麦灌浆期叶片营养元素N、P含量下降,K含量变化范围较小(表4),2039品系的N含量高于对照品种11.57%,2038品系的P含量高于对照6.35%,2026品系的K含量高于对照49.5%,大穗型小麦新品系和对照处理N/K差异均不显著,灌浆期新品系的平均K、N/P都高于对照品种。总之,大穗型小麦不同生育期叶片营养元素含量均变化明显,不同小麦品系的变化程度也不尽相同。
2.4 小麦抽穗期叶片性状与营养元素的相关性
在小麦抽穗期SLA和LDMC呈极显著负相关关系,SLA和N含量呈不明显的正相关,和P、K呈极显著的负相关关系,N与P表现出一定的不明显正相关(表5),小麦旗叶叶性状与养分含量之间表现出的相关性各不相同。
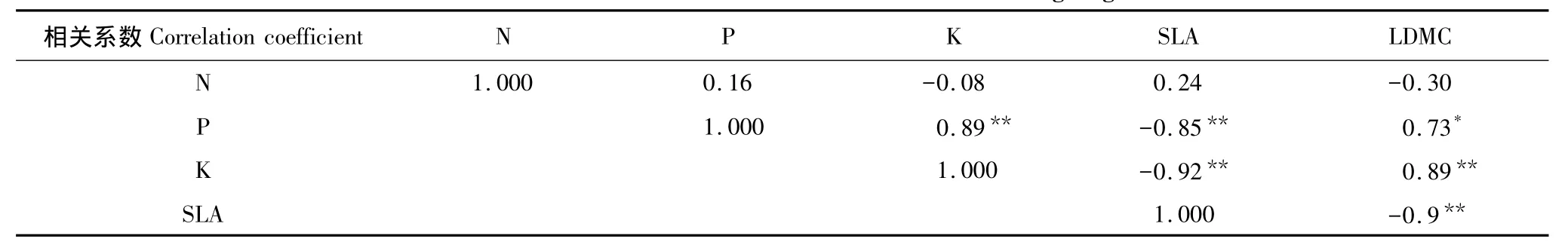
表5 小麦抽穗期叶片性状与营养元素之间的相关性分析Table5 Correlation of leaf traits and nutrient in heading stage
2.5 小麦成熟期氮素在小麦不同器官的积累与分配
小麦成熟期不同品系(种)氮素含量和积累量存在差异,各器官氮素积累量及分配比例不同(表6),成熟期各器官氮素积累能力和分配比例大小顺序为籽粒>茎+鞘>穗草>旗叶>倒二叶>余叶,说明籽粒是活性最大的库,成熟期营养器官中的氮素转运至籽粒,籽粒中氮素积累量和分配比例较大。小麦新品系各器官氮素积累能力都高于对照品种,氮素积累量和分配比例均以籽粒最高,余叶最低。大穗型小麦新品系籽粒氮素的积累量可达300kg/hm2以上,2037和2038品系旗叶与倒二叶氮素积累和分配比例较高,余叶器官以2040品系的积累和分配比例为高,说明生育后期2040品系余叶含有较高的氮含量,2013品系的茎+鞘氮素积累和分配比例较高,穗草的氮素积累量和分配比例以2026品系为高,大穗型新品系器官氮素积累量和分配比例大多高于对照品种,不同大穗型品系之间积累量和分配比例亦不同,说明器官氮含量的高低取决于品系(种)本身的遗传特性,大穗小麦的氮素积累能力可能强于西农979。
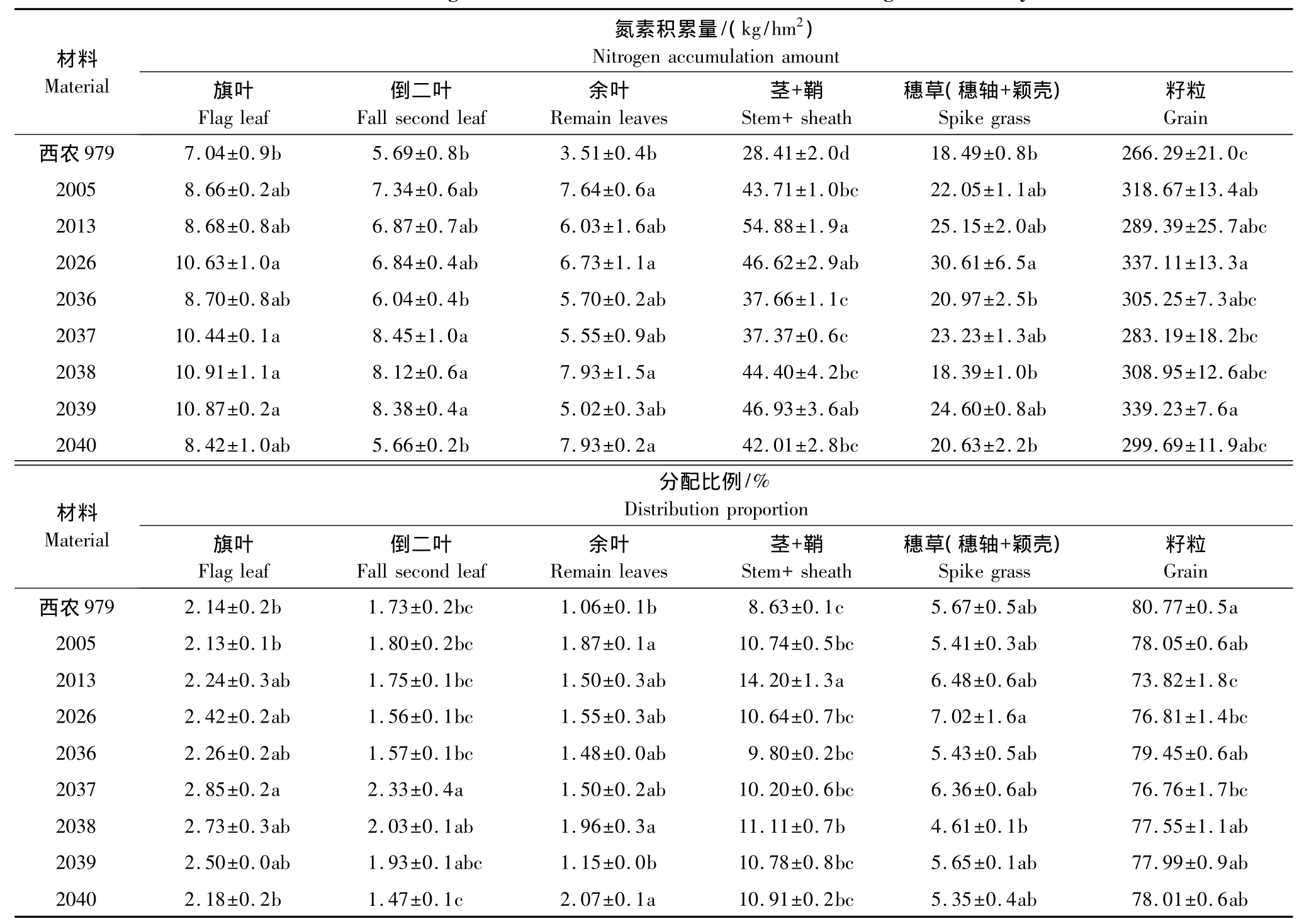
表6 小麦成熟期氮素在不同器官中的积累与分配Table6 The nitrogen accumulation and distribution in different organs at maturity
供试的8个大穗型小麦品系开花期平均营养器官氮素积累量、成熟期平均籽粒积累量分别高于对照品种10.31%、16.48%(表7),2037品系的贡献率高于对照品种的11.5%,转移率除2013、2039品系外新品系和对照品种差异不显著,开花后大穗品系向籽粒的平均转移量、转移率和贡献率低于西农979,表明大穗型小麦新品系开花前的高营养器官氮素积累量向籽粒供应氮素的能力有待提高。
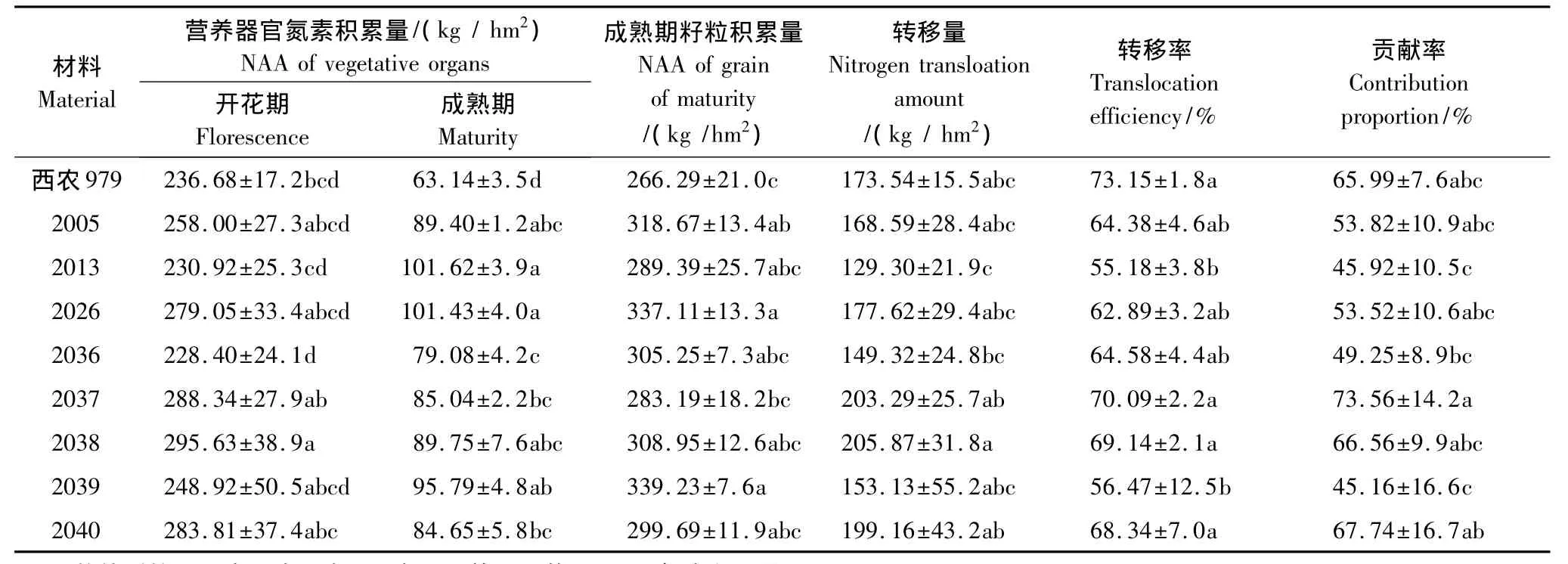
表7 小麦花后营养器官氮素向籽粒的转移情况Table7 Nitrogen translocation from vegetative organs to grain after anthesis
3 讨论
植物生长特性和形态特征的改变是适应各种环境的重要策略[16],尤其是植物叶片性状更能反映植物对环境的适应程度及环境变化对于植物的影响[17]。本试验结果表明不同小麦品系SLA和LDMC在不同生育期具有不同的响应,而且不同的小麦品系和品种在同一生育期的变化也不尽相同,大穗型小麦品系平均SLA和LDMC均小于西农979,这可能与其自身N可利用性、水分利用状况和一些遗传特征有关。植物LDMC增加,相应SLA减小,能降低植物体内的水分散失[18],并增强其抗非生物侵害的能力[19]。本试验中所有参试小麦材料,以2040品系的平均SLA较小,LDMC含量较高,使叶片内部水分向叶片表面扩散的距离或阻力扩大,叶片相对更坚韧,因此,2040品系叶片较高的LDMC较能抵抗非生物胁迫且有较好的资源保有能力和水分利用效率。
本研究中大穗型小麦品系养分含量和叶片性状在不同的生育时期变化较为敏感,各小麦品系(种)由于自身遗传特性的不同而表现出与环境协同变化的显著特征,郑淑霞和上官周平[20]对黄土高原126种植物叶片进行研究发现叶片N、P、K的变化范围分别为0.82%—4.58%、0.06%—0.35%和0.24%—4.21%,叶片N/P的变化范围为7—29。本研究中小麦拔节期2013品系、抽穗期CK、2013、2039和2040品系的N含量略有超出,而抽穗期和灌浆期N/P大部分超出此范围,这表明不同研究区域、不同研究材料其变化范围有所不同。养分N和P是陆地植物主要限制元素,N/P的比例是描述植结构、功能和养分限制的重要指标[21],Koerselman等[22]认为当N/P<14时,植物生长主要受N限制;N/P>16时,植物生长主要受P限制。在本试验中小麦叶片N/P的比例均大于16,并且生育期中有超过或接近30的比值,说明该地区参试小麦品系或品种的生长主要受P限制,这可能与该区域土壤P含量或小麦本身的生长发育特性有关。小麦叶片性状与养分含量的变化与环境条件关系密切,本研究分析了其抽穗期叶片性状之间的相关性,SLA和LDMC呈负相关关系,这在有些试验中已得到证实[2,23],郑淑霞和上官周平[20]指出黄土高原126个植物样叶片N、P之间呈极显著的正相关关系,本研究与邓蕾等[10]研究表明的水蚀风蚀交错区锦鸡儿叶片SLA与N、P、K之间表现一定的不显著正相关研究结果不太一致,这说明影响植物SLA的因素较为复杂,除了本身的水分和养分外,可能还存在一些植物群体之间的竞争关系等,小麦不同发育阶段对营养元素的需求也不同。
小麦产量和品质与氮素的吸收、积累和转运密切相关,遗传特性和栽培措施等对其有重要影响。小麦开花后营养器官氮素的转运对籽粒氮素累积有较大贡献,籽粒中的氮素有相当一部分来自于营养器官氮素的花后再分配[24]。本项试验表明,8个大穗型小麦品系成熟期各器官氮素积累能力,除2038品系的穗草和2040品系的倒二叶氮积累量低于西农979外,都显著高于其对照品种;成熟期各器官氮素积累量及分配比例顺序为籽粒>茎+鞘>穗草(穗轴+颖壳)>旗叶>倒二叶>余叶。邵云等以西农979品种为试材,认为成熟期不同器官氮积累量从大到小依次为籽粒>叶鞘>叶>茎>颖壳>穗轴,这主要因为小麦开花期和成熟期氮素转运受环境条件、施肥和品种的制约[25-26],所研究的器官和对象的不同,小麦不同器官氮素积累量与运转情况亦不同。Xu等[27]研究表明,灌水量过高或过低时小麦氮素的转移量和转移率均降低,适当水分亏缺有利于提高营养器官贮存的氮素对籽粒氮的贡献率[28],土壤干旱促进了氮素从叶片向籽粒的转移,提高了籽粒氮素水平[29]。另有研究认为保持开花后适宜的土壤相对含水量可提高氮素转移量[30],段文学等的测墒补灌法表明,补灌时期不同对品种开花后营养器官氮素转运的调节效应不同[31],在本试验条件下大穗型小麦品系的转移率均低于西农979,2026、2037、2038和2040品系在单位面积内的氮素转移量高于对照,2037、2038和2040品系营养器官对籽粒的氮素贡献率较西农979为高,所以在实际生产中,充分利用残留在茎秆和其他营养器官中的氮素尤为重要,合理的水肥调控措施对提高氮素利用效率具有重要作用。
总之,提高小麦的氮素利用效率,一方面应重视小麦品种的营养高效性,从遗传改良来提高小麦氮素利用效率,另一方面可通过合理的水肥调控措施,以实现以肥调水、以水促肥的效果。另外,小麦叶片性状因品种和生境条件而异,由本试验结果可知相同的生长环境和农艺管理条件下,大穗型小麦新品系和对照品种对环境条件的响应策略有所不同。在植物叶片结构性状中,SLA还受叶厚的影响较大,两者呈负相关关系[32-33],小麦SLA和LDMC与叶片厚度的关系还鲜有报道,由于产量形成是由很多性状决定的,为优化调控小麦叶片结构性状和养分利用,小麦叶片主要性状因子之间的综合作用及其管理措施的优化仍需做进一步研究。
[1] Zhao CJ,Guo X W,Zhang Q D,Kang S J.Fluorescence induction transient and emission spectrum characteristics of winter wheat leaves of different spike type.Scientia Agricultura Sinica,1999,32(2):43-46.
[2] Vile D,GarnierÉ,Shipley B,Laurent G,Navas M L,Roumet C,Lavorel S,Díaz S,Hodgson J G,Lloret F,Midgley G F,Poorter H,Rutherford M C,Wilson P J,Wright I J.Specific leaf area and dry matter content estimate thickness in laminar leaves.Annals of Botany,2005,96(6):1129-1136.
[3] Pontes L D S,Soussana J F,Louault F,Andueza D,Carrère P.Leaf traits affect the above-ground productivity and quality of pasture grasses.Functional Ecology,2007,21(5):844-853.
[4] Li Y L,Cui JH,Su Y Z.Specific leaf area and leaf dry matter content of some plants in different dune habitats.Acta Ecologica Sinica,2005,25(2):304-311.
[5] Watson D J.The physiological basis of variation in yield.Advances in Agronomy,1952,4:101-145.
[6] Fu Z L,Ma B Z,Wang G J,Zhao Y H,Li G K.Relationship between the flag leaf and the grain weight per spike in wheat.Journal of Triticeae Crops,2001,21(1):92-94.
[7] Guo W S,Feng C N,Yan L L,Peng Y X,Zhu X K,Zong A G.Analysis on source-sink relationship after anthesis in wheat.Acta Agronomica Sinica,1995,2l(3):334-340.
[8] Smith CJ,Whitfield D M.Nitrogen accumulation and redistribution of late applications of15N-labelled fertilizer by wheat.Field Crop Research,1990,24(3/4):211-226.
[9] Wright I J,Reich P B,Westoby M,Ackerly D D,Baruch Z,Bongers F,Cavender-Bares J,Chapin T,Cornelissen JH C,Diemer M,Flexas J,Garnier E,Groom P K,Gulias J,Hikosaka K,Lamont B B,Lee T,Lee W,Lusk C,Midgley JJ,Navas M L,NiinemetsÜ,Oleksyn J,Osada N,Poorter H,Poot P,Prior L,Pyankov V I,Roumet C,Thomas SC,Tjoelker M G,Veneklaas E J,Villar R.The worldwide leaf economics spectrum.Nature,2004,428(6985):821-827.
[10] Deng L,Wang H Z,Shangguan Z P,Liu G C.Variations of specific leaf area and nutrients of Chinese caragana in the Loess Plateau region suffering both wind and water erosions.Acta Ecologica Sinica,2010,30(18):4889-4897.
[11] Zhang Q J,Zhang L Y,Bi H W.The absorption,accumulation and translocation of nitrogen and their relationships to grain protein content in spring wheat variety.Acta Agronomica Sinica,1997,23(6):712-718.
[12] Zhu X K,Guo W S,Feng CN,Peng Y X,Ling Q H.Nitrogen absorption and utilization differences among wheat varieties for different end uses.Plant Nutrition and Fertilizer Science,2005,11(2):148-154.
[13] Garnier E,Shipley B,Roumet C,Laurent G.A standardized protocol for the determination of specific leaf area and leaf dry matter content.Functional Ecology,2001,15(5):688-695.
[14] Page A L,Miller R H,Keeney D R.Methods of Soil Analysis.Part 2.Chemical and Microbiological Properties.2nd ed.Madison:American Society of Agronomy Press,1982.
[15] Zhao M X,Zhou JB,Yang R,Zheng X F,Zhai B N,Li SX.Characteristics of nitrogen accumulation,distribution and translocation in winter wheat on dryland.Plant Nutrition and Fertilizer Science,2006,12(2):143-149.
[16] Shi SB,Li H M,Wang X Y,Yue X G,Xu W H,Chen G C.Comparative studies of photosynthetic characteristics in typical alpine plants of the Qinghai Tibet Plateau.Acta Phytoecologica Sinica,2006,30(1):40-46.
[17] Meziane D,Shipley B.Interacting determinants of specific leaf area in 22 herbaceous species:effects of irradiance and nutrient availability.Plant,Cell and Environment,1999,22(5):447-459.
[18] Cornelissen J H C,Lavorel S,Garnier E,Díaz S,Buchmann N,Gurvich D E,Reich P B,ter Steege H,Morgan H D,van der Heijden M G A,Pausas JG,Poorter H.A handbook of protocols for standardized and easy measurement of plant functional traits worldwide.Australian Journal of Botany,2003,51(4):335-380.
[19] Lü J,Li JQ.Research on leaf traits and their interrelationships of plants in Beijing Baihua Mountain.Science Technology and Engineering,2008,8(12):3287-3289.
[20] Zheng SX,Shangguan Z P.The spatial patterns of the leaf nutrient composition in the Loess Plateau.Progress in Natural Science,2006,16(8):965-973.
[21] Reich P B,Oleksyn J.Global patterns of plant leaf N and P in relation to temperature and latitude.Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[22] Koerselman W,Meuleman A F M.The vegetation N:P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33(6):1441-1450.
[23] Shipley B,Vu T T.Dry matter content as a measure of dry matter concentration in plants and their parts.New Phytologist,2002,153(2):359-364.
[24] Guitman M R,Arnozis P A,Barneix A J.Effect of source-sink relations and nitrogen nutrition on senescence and N remobilization in the flag leaf of wheat.Physiologia Plantarum,1991,82(2):278-284.
[25] Barbottin A,Lecomte C,Bouchard C,Jeuffroy M H.Nitrogen remobilization during grain filling in wheat:genotypic and environmental effects.Crop Science,2005,45(3):1141-1150.
[26] Shao Y,Li W C,Feng R C,Jiang L N,Yao L J,Li CX.Nitrogen translocation and conversion rate to grain of Xinong 979 in wheat-corn rotation area.Journal of Triticeae Crops,2011,31(2):265-269.
[27] Xu Z Z,Yu Z W,Wang D.Nitrogen translocation in wheat plants under soil water deficit.Plant and Soil,2006,280(1/2):291-303.
[28] Xu Z Z,Yu Z W,Wang D,Zhang Y L.Nitrogen accumulation and translocation for winter wheat under different irrigation regimes.Journal of Agronomy and Crop Science,2005,191(6):439-449.
[29] Sinclair T R,Pinter P JJr,Kimball B A,Adamsen F J,LaMorte R L,Wall GW,Hunsaker DJ,Adam N,Brooks T J,Garcia R L,Thompson T,Leavitt S,Matthias A.Leaf nitrogen concentration of wheat subjected to elevated[CO2]and either water or N deficits.Agriculture Ecosystems and Environment,2000,79(1):53-60.
[30] Ma D H,Wang Y F,Zhao C X,Lin Q.Effects of nitrogen fertilizer rate and post-anthesis soil water content on characteristics of nitrogen metabolism and grain protein content in wheat.Plant Nutrition and Fertilizer Science,2008,14(6):1035-1041.
[31] Duan W X,Zhang Y L,Shi Y,Yu Z W.Effects of soil-moisture monitoring supplemental irrigation on water consumption characteristics and nitrogen accumulation and translocation in different wheat cultivars.Plant Nutrition and Fertilizer Science,2011,17(6):1309-1317.
[32] Westoby M.A leaf-height-seed(LHS)plant ecology strategy scheme.Plant and Soil,1998,199(2):213-227.
[33] Wilson P J,Thompson K,Hodgson JG.Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.New Phytologist,1999,143(1):155-162.
参考文献:
[1] 赵春江,郭晓维,张其德,康书江.不同穗型冬小麦叶片荧光诱变及光谱特性.中国农业科学,1999,32(2):43-46.
[4] 李玉霖,崔建垣,苏永中.不同沙丘生境主要植物比叶面积和叶干物质含量的比较.生态学报,2005,25(2):304-311.
[6] 傅兆麟,马宝珍,王光杰,赵玉华,李广科.小麦旗叶与穗粒重关系的研究.麦类作物学报,2001,21(1):92-94.
[7] 郭文善,封超年,严六零,彭永欣,朱新开,宗爱国.小麦开花后源库关系分析.作物学报,1995,2l(3):334-340.
[10] 邓蕾,王鸿喆,上官周平,刘广全.水蚀风蚀交错区柠条锦鸡儿叶片比叶面积和营养元素变化动态.生态学报,2010,30(18):4889-4897.
[11] 张庆江,张立言,毕恒武.春小麦品种氮的吸收积累和转运特征及与籽粒蛋白质的关系.作物学报,1997,23(6):712-718.
[12] 朱新开,郭文善,封超年,彭永欣,凌启鸿.不同类型专用小麦氮素吸收积累差异研究.植物营养与肥料学报,2005,11(2):148-154.
[15] 赵满兴,周建斌,杨绒,郑险峰,翟丙年,李生秀.不同施氮量对旱地不同品种冬小麦氮素累积、运输和分配的影响.植物营养与肥料学报,2006,12(2):143-149.
[19] 吕佳,李俊清.北京百花山植物叶片性状相互关系研究.科学技术与工程,2008,8(12):3287-3289.
[20] 郑淑霞,上官周平.黄土高原地区植物叶片养分组成的空间分布格局.自然科学进展,2006,16(8):965-973.
[26] 邵云,李万昌,冯荣成,姜丽娜,姚利娇,李春喜.小麦-玉米轮作区西农979不同器官氮素转运及对籽粒贡献率的影响.麦类作物学报,2011,31(2):265-269.
[30] 马东辉,王月福,赵长星,林琪.施氮量和花后土壤含水量对小麦氮代谢特性和子粒蛋白质含量的影响.植物营养与肥料学报,2008,14(6):1035-1041.
[31] 段文学,张永丽,石玉,于振文.测墒补灌对不同小麦品种耗水特性和氮素分配与转运的影响.植物营养与肥料学报,2011,17(6):1309-1317.
猜你喜欢
农技服务(2023年2期)2023-03-15
园林科技(2020年2期)2020-01-18
山东林业科技(2019年2期)2019-06-03
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
江苏农业科学(2017年1期)2017-02-27
湖北农业科学(2014年16期)2014-10-20
江苏农业科学(2014年7期)2014-09-02
作物研究(2014年6期)2014-03-01
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
