
寄主大小及寄生顺序对蝇蛹佣小蜂寄生策略的影响
2013-12-16 08:19詹月平陈中正段毕升胡好远
生态学报 2013年11期
詹月平,周 敏,贺 张,陈中正,段毕升,胡好远,*,肖 晖
(1.安徽师范大学生命科学学院,安徽省高校生物环境与生态安全重点实验室,芜湖 241000;2.中国科学院动物研究所,动物进化与系统学重点实验室,北京 100101)
寄生蜂存在着对寄生行为的主动调控能力。研究表明,一些寄生蜂在选择寄主时具有区分寄主是否被寄生的能力以及对寄主大小和年龄的判定能力等[1-3]。寄生蜂根据种群结构特征和环境资源等因素主动调节性比的现象是性别分配理论的重要证据。寄生蜂为单双倍型性别决定方式,一般来说,此类动物未受精的单倍型卵发育成雄体,而受精的二倍型卵发育成雌体;其母体可通过控制受精作用主动调节后代性比[2-6]。人们认为寄主大小等质量特征是影响其后代质量的主要环境资源因素,从大寄主上羽化成熟的寄生蜂后代个体较大;寄主大小模型认为寄生蜂后代性比与寄主大小相关,产卵者倾向于在大寄主上产出更多雌性后代[3-8]。
蝇蛹佣小蜂(Spalangia endius)属于膜翅目Hymenoptera、小蜂总科Chalcidoidea、金小蜂科Pteromalidae,为世界范围内的广布种[9]。蝇蛹佣小蜂在单个寄主内一般仅有单头后代能完成发育,属于单寄生性寄生蜂,并且寄生时,能将寄主麻痹,属于抑性寄生蜂。该蜂在选择寄主时,具有对寄主大小和年龄进行判定、调节后代性比的能力[3,10]。但是,该寄生蜂还具有过寄生习性,相关研究均未排除过寄生影响[3,10]。为了探讨蝇蛹佣小蜂的生物学特性和过寄生对寄生蜂性比策略的影响程度,本文研究了以家蝇蛹为寄主时,蝇蛹佣小蜂在产卵期中的雌雄后代数量和雄性百分比,探讨了单次寄生情况下,寄主大小及寄生顺序对寄生蜂后代性比等的影响。
1 材料与方法
1.1 寄生蜂来源和培养
供试家蝇(Musca domestica)采自安徽师范大学赭山校区食堂附近,并参照何凤琴[11]的方法在室内进行人工饲养,以获得家蝇蝇蛹。定期将1—2日龄蝇蛹放入黄色的广口容器中,覆以大孔纱网,置于校园内诱集寄生蜂。蝇蛹放置3—4d后取回,并每天定时观察是否有寄生蜂产出。同时,对从家蝇蛹中羽化出的寄生蜂进行形态鉴定,获得蝇蛹佣小蜂,建立繁殖系,并连续在实验室内饲养1a左右。饲养该蜂的培养器皿为直径5 cm、高9 cm的透明玻璃杯,蝇蛹和小蜂比例为10∶1,同时饲以10%的蜂蜜水。培养在人工气候箱内进行,温度为(25±1)℃,光照强度为60%(约2000lx)(L ∶D=14∶10),相对湿度为 RH=(50±5)%。
1.2 寄生蜂存活期内的产卵规律
使用吸虫管收集羽化24h内的蝇蛹佣小蜂,集中于上述玻璃杯中,喂食蜂蜜水,让寄生蜂自由交配24h,进行下一步实验。寄生蜂产卵规律实验共设30个重复组,每杯为1个重复。在上述的玻璃杯中引入1只交配后的雌蜂,接入10头1—2日龄家蝇蛹,同时以棉絮沾取10%蜂蜜水补充营养。每天定时取出杯中蝇蛹,再放入10头1—2日龄的蝇蛹并更换蜂蜜水棉球。实验持续至雌蜂死亡。每日将各杯中更换后的家蝇蛹分别装入直径1cm、高5cm的冻存管中,棉塞封口。定期记录各管内羽化的雄蜂和雌蜂数量,管内蝇蛹在体式显微镜下镜检,记录未能成功钻出寄主蛹的寄生蜂后代。统计获得寄生蜂的成蜂寿命、产卵期、每日雌蜂和雄蜂数量及性比、雌蜂和雄蜂的发育历期等。数据经过筛选,排除仅产雄性后代的重复组,有效实验重复组为28组。
1.3 家蝇蛹大小对蝇蛹佣小蜂寄生的影响
通过控制家蝇幼虫培养杯中幼虫密度,获得大小不一的家蝇蛹,使用游标卡尺估计蛹大小,大蛹长度在5.2 mm以上,小蛹在4.8 mm以内。蝇蛹佣小蜂单次寄生仅产下单枚卵[12]。根据寄生蜂寄生行为的发生,可以获得仅仅寄生单次的寄主,从而排除过寄生的影响。实验共设20个重复组,每个重复组内置1—2日龄内的大蛹和小蛹各5头,大蛹和小蛹相间排列成圆环状。取集中交配后的雌蜂1头,置于上述玻璃杯中,倒扣在蛹的上方,杯内置沾有蜂蜜水的棉球,每个杯为1重复。按寄生蜂对蛹寄生的先后确定寄生顺序,持续观察记录寄生蜂寄生行为,待寄生蜂完成1次寄生行为并离开寄主后,立即取出被寄生的蛹并按顺序记录,实验持续至连续4h寄生蜂均不再出现寄生行为为止。实验后的家蝇蛹分别装入直径1cm、高5cm的冻存管中,棉塞封口。待出蜂完毕,观察记录每管内羽化的雄蜂、雌蜂和家蝇数量,管内蝇蛹在体式显微镜下镜检。使用数码体式显微镜(MC-D310U,凤凰光学集团有限公司)进行蛹和寄生蜂图像拍摄,并测量寄生蜂后足胫节长度、寄主蛹的长度和宽度,蛹的体积计算参考Napoleon和King[10],按长椭球形计算体积:V=2/3×π×(蛹宽/2)2×蛹长。数据经过筛选,排除仅产雄性后代的重复组,有效实验重复组为18组。
1.4 数据分析
在R2.6.1软件中使用广义线性模型(Generalized Linear Model,GLM)分析了雌蜂产卵期对寄生蜂后代数量、后代性比的影响。数量数据使用泊松分布模型,比例数据使用二项分布模型。建立模型后,根据模型的HF(残差/df)值判定数据与泊松分布或二项分布的符合程度。较大的HF值(HF>1)意味着偏离泊松分布或二项分布,会导致显著性检验程度被高估;用近似泊松分布(quasipoisson)或近似二项分布(quasibinomial)进行模型拟合,对最后获得的模型用F检验进行分析[13]。其它分析使用SPSS11.5软件完成,在比较寄生蜂雌雄两性后代发育历期数据时,使用双样本异方差t检验,自由度经过了校正。
2 结果
2.1 蝇蛹佣小蜂的产卵期和后代数量变化
蝇蛹佣小蜂的产卵期为(8.93±3.34)d(N=28),最长产卵期为16 d,平均寿命为(10.86±3.27)d(N=28),最长寿命 19 d。单个雌蜂能生产雌性后代(34.11±16.34)头和雄性后代(11.04±8.87)头,并且后代中雄性百分比为0.24±0.11。
寄生蜂雌雄两性后代发育历期分别为(18.74±1.38)d和(18.09±1.92)d,两者间差异极显著(t415.70=5.49,P<0.01)。随寄生蜂成蜂日龄的增大,寄生蜂每天所产后代的数量趋于减少(F1,302=74.18,P<0.01),并且后代中雌蜂数量显著降低(F1,302=78.60,P<0.01),而雄性数量显著增加(F1,302=4.24,P=0.040)(图1),雄性百分比显著增大(F1,231=13.14,P<0.01)。
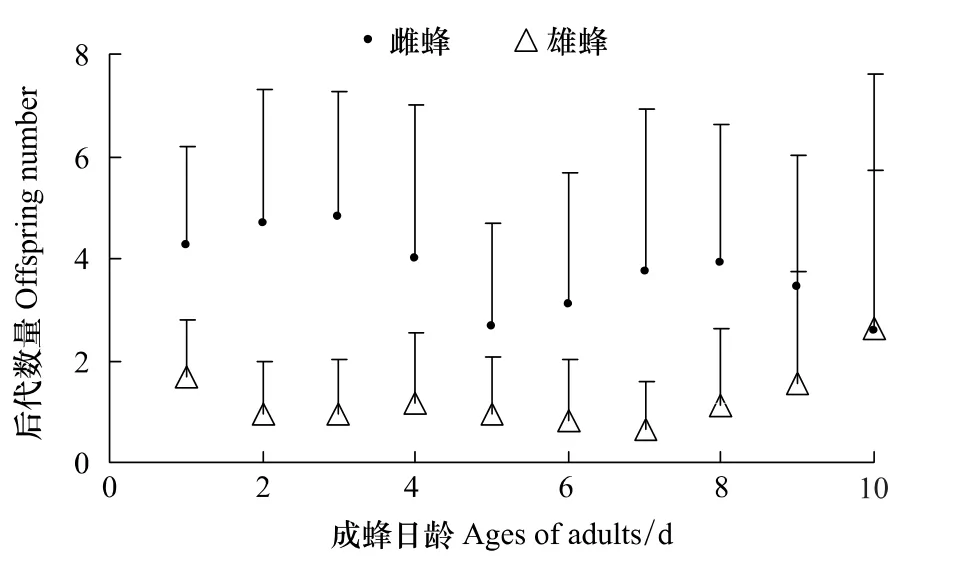
图1 不同日龄时蝇蛹佣小蜂的后代数量(平均数±标准差)Fig.1 Progeny number of Spalangia endius at different adult ages
2.2 蝇蛹佣小蜂对不同大小的寄主的选择顺序、后代性比和个体大小
蝇蛹佣小蜂寄生家蝇蛹时,存在明显地优先选择较大寄主蛹的习性,寄生顺序与蛹大小间存在显著的负相关(r=-0.22,P=0.01)(图2)。
在寄生行为发生后,寄主蝇蛹仅部分可羽化出寄生蜂,部分蝇蛹既无寄生蜂产出也无家蝇羽化,部分蝇蛹仍能羽化出家蝇(表1),该结果表明寄生行为发生后,寄生蜂并非能够成功完成寄生。在发生了寄生行为的家蝇蛹中,能羽化出寄生蜂的蛹的比例不随寄生顺序的变化而显著变化(F1,8=1.83,P=0.21),羽化家蝇的蛹比例也不随寄生顺序的变化而显著改变(F1,8=0.78,P=0.40)。
对所获得的寄生蜂后代胫节长度进行了测量,结果表明雌、雄蜂后代胫节长度为分别为(0.46±0.02)mm(N=34)和(0.43±0.03)mm(N=14),两者间存在极显著差异(t46=3.98,P<0.001),雄蜂显著小于雌蜂。
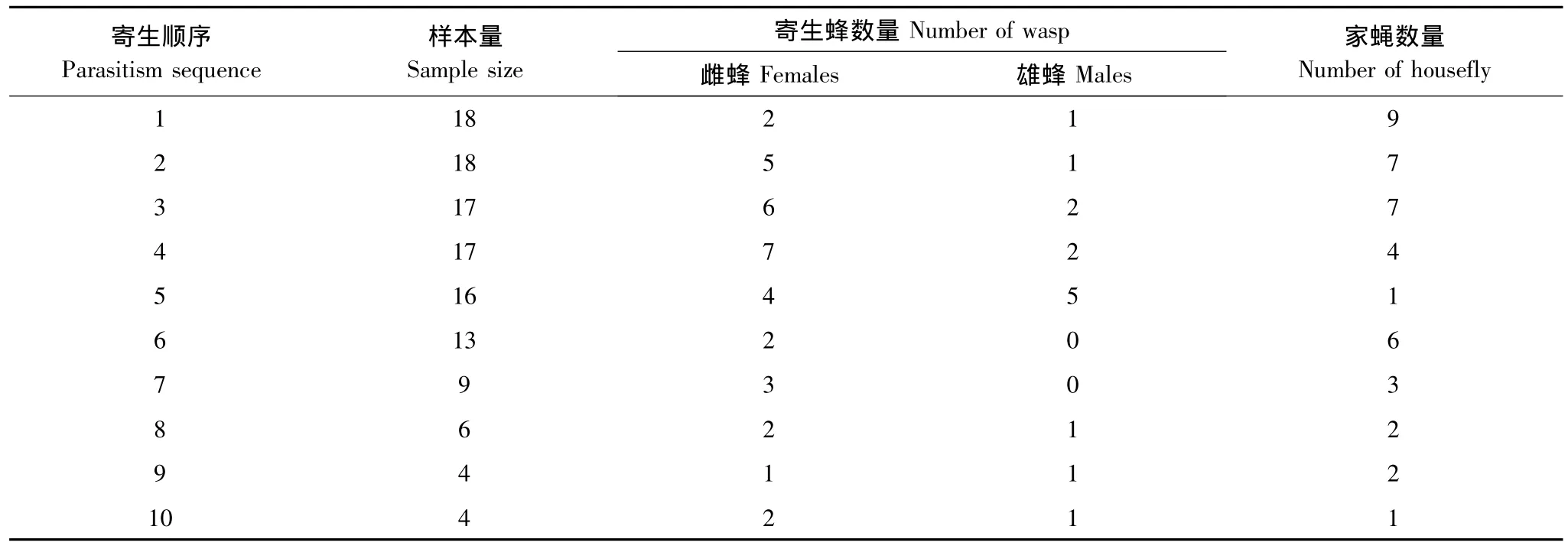
表1 不同寄生顺序时寄主内羽化的寄生蜂和家蝇数量Table 1 Number of parasitoid wasps and houseflies from hosts at different parasitism sequences
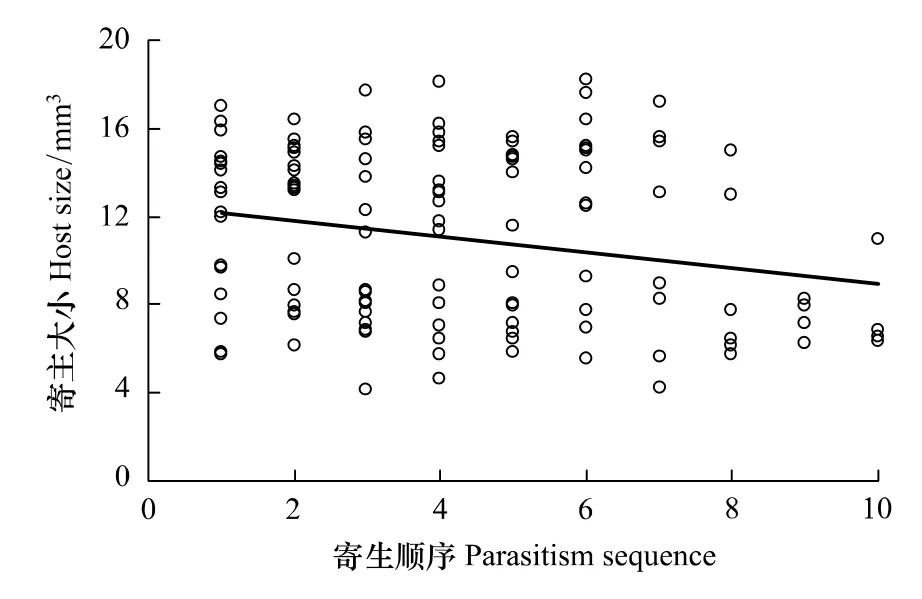
图2 蝇蛹佣小蜂产卵顺序与家蝇蛹大小之间关系Fig.2 Relationship between housefly pupae size and parasitism sequences of Spalangia endius
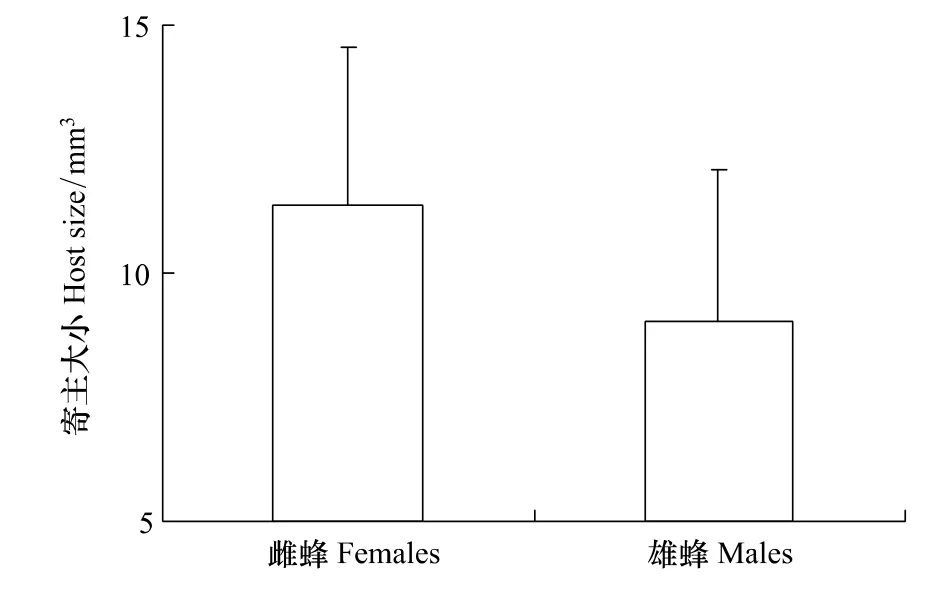
图3 蝇蛹佣小蜂雌雄后代的寄主家蝇蛹大小比较Fig.3 Size of housefly pupae emerged the female and male progeny of Spalangia endius
羽化雌雄蜂的家蝇蛹大小分别为(11.38±3.20)mm3和(9.02±3.06)mm3,两者间差异显著(t46=2.35,P=0.02)。由此说明,在较大的寄主蛹上,寄生蜂倾向于产更多的雌性后代(图3)。
3 讨论
寄生蜂在寄生时,存在对寄主的判定和选择[1-3]。寄生蜂可以根据寄主是否被寄生,来选择性地调整寄生策略[1,12,14]。对于寄生蜂而言,寄主大小和龄期等质量特征是影响其后代质量的主要环境资源因素;寄主大小模型认为寄生蜂后代性比与寄主大小相关,产卵者倾向于在大寄主上产出更多雌性后代,在小寄主上产出更多雄性后代[3-8]。寄主大小模型已在多种寄生蜂中得到证实,尤其是寄生时即杀死寄主的抑性(idiobiont)寄生蜂[6]。然而,很多寄生蜂都具有过寄生性特点,在对蝇蛹佣小蜂性比与寄主大小关系的研究方面,相关研究均未排除过寄生的影响。在寄主数量较少时,蝇蛹佣小蜂过寄生常常发生,在单头寄主上,产下多枚卵[12]。当单头雌蜂在一个寄主上多次产卵时,过寄生可能会导致寄生蜂后代性比的改变,如过寄生时死亡率增加且雌雄两性间死亡率存在差异,过寄生将引起性比改变,偏向存活率更高的性别;在控制实验中,由于常常给予较少的寄主和较长的寄生时间,过寄生更易发生[15]。单寄生性的前裂长管茧蜂(Diachasmimorpha longicaudata)在寄生墨西哥按实蝇(Anastrepha ludens)蛹时,过寄生显著影响到寄生蜂后代的性比,过寄生比例越高,后代雄性百分比越高[16]。对同属于佣小蜂属的寄生蜂S.cameroni的研究也表明,过寄生会导致寄生蜂后代性比偏雌[17]。单寄生性寄生蜂发生过寄生行为时,雌性后代比雄性后代具有更强的竞争力,从而导致次级性比与初级性比之间的偏离[16-18]。本文的结果排除了过寄生的影响,表明在单次寄生时,蝇蛹佣小蜂后代性比与寄主大小之间关系符合寄主大小模型,寄生蜂能够判定寄主大小并在较大的寄主上产下更多的雌性后代。
寄生蜂对寄主大小的选择行为的适应价值体现在不同大小雌雄后代的价值差异上;较大雌蜂比较大雄蜂更有价值,如较大雌蜂具有更长的产卵历期[3-6,10]、更大的后代产量[3-6,10]、更强的扩散能力[10]等,而大小不同的雄蜂在交配能力等方面差异不大[3,10,19]。相比较而言,在较大的寄主上,雌雄两性后代个体均呈增大趋势;但由于较大的雌蜂后代更有意义,对产卵者来说,在较大的寄主上产下更多的雌性后代的行为具有更大的适合度[4-5,20]。
在寄生顺序上,寄生蜂仅选择合适大小(龄期)的寄主进行寄生,如白蜡吉丁柄腹茧蜂(Spathius agrili)在寄生白蜡窄吉丁(Agrilus planipennis)幼虫时,仅选择3—4龄寄主产卵[21]。寄生蜂存在对大小寄主选择顺序差异,寄生松墨天牛(Monochamus alternatus)幼虫的聚寄生性寄生蜂哈氏肿腿蜂(Sclerodermus harmandi)优先寄生较大的寄主,在较大寄主上,产卵频率较高[22]。这种选择性反映了寄生蜂的主动调控能力,相比较而言,较大寄主能提供更丰富的营养[5,14,22],寄生蜂后代具有较低的死亡率[22-23]。研究结果表明,在蝇蛹佣小蜂寄生时,较大的寄主被优先选择,表明蝇蛹佣小蜂存在对寄主大小的判定能力并能根据寄主大小进行选择性产卵。
在蝇蛹佣小蜂的整个产卵期中,寄生蜂后代性比一般偏雌,但在产卵后期,蝇蛹佣小蜂的雄性百分比呈现增大的趋势。一般来说,此类动物未受精的单倍型卵发育成雄体,而受精的二倍型卵发育成雌体;交配后,精子储存在雌蜂的受精囊中;雄性比例的增加表明该产卵者可能在产卵后期出现精子枯竭。大多数寄生蜂在生活史中常常仅能够交配1次,为单配制[24],对蝇蛹佣小蜂的观察也证明了这一点。精子枯竭现象在寄生蜂中,可能较多地存在,对寄生蜂 Uscana semifumipennis、Anaphes nitens和蝇蛹金小蜂(Pachycrepoideus vindemmiae)的研究均发现寄生蜂在产卵后期的雄性后代比例增大和精子枯竭现象[25-28]。精子枯竭而导致的雄性比例增大可能是对寄生蜂种群偏雌性比的一种适应,这需要进一步的研究才能揭示。
蝇蛹佣小蜂为多种害虫蛹期的重要寄生蜂,多数寄主为世界范围内的重大害虫,如橘小实蝇(Bactrocera dorsalis)、瓜实蝇(B.cucurbitae)、地中海果实蝇(Ceratitis capitata)、按实蝇(Anastrepha spp.)和家蝇等[9,29-30]。在对害虫的控制上,雌蜂具有更大价值;根据掌握的性别分配行为规律,可以生产出更多、个体更大的雌蜂,从而提高生产效率,促进对害虫的控制效果。蝇蛹佣小蜂具有产卵期长,后代产量高的特点,具备很大的应用潜力。
[1] Goubault M,Krespi L,Boivin G,Poinsot D,Nenon J P,Cortesero A M.Intraspecific variations in host discrimination behavior in the pupal parasitoid Pachycrepoideus vindemmiae Rondani(Hymenoptera:Pteromalidae).Environmental Entomology,2004,33(2):362-369.
[2] West S A.Sex allocation.Princeton:Princeton University Press,2009.
[3] King B H,Napoleon M E.Using effects of parasitoid size on fitness to test a host quality model assumption with the parasitoid wasp Spalangia endius.Canadian Journal of Zoology,2006,84(11):1678-1682.
[4] Charnov E L,Los-den Hartogh R L,Jones W T,van den Assem J.Sex ratio evolution in a variable environment.Nature,1981,289(5793):27-33.
[5] Charnov E L.The theory of sex allocation.Princeton:Princeton University Press,1982.
[6] West S A.Sheldon B C.Constraints in the evolution of sex ratio adjustment.Science,2002,295(5560):1685-1688.
[7] Zhang X L,Meng L,Li B P.Effects of Pieris rapae pupa mass on offspring number,sex ratio,and body size of gregarious parasitoid Pteromalus puparum(Hymenoptera:Pteromalidae).Chinese Journal of Ecology,2009,28(4):677-680.
[8] Pekas A,Aguilar A,Tena A,Garcia-Marí F.Influence of host size on parasitism by Aphytis chrysomphali and A.melinus(Hymenoptera:Aphelinidae)in Mediterranean populations of California red scale Aonidiella aurantii(Hemiptera:Diaspididae).Biological Control,2010,55(2):132-140.
[9] Noyes J S.Interactive catalogue of world Chalcidoidea(CD-ROM).London:Taxapad and The Natural History Museum,2002.
[10] Napoleon M E,King B H.Offspring sex ratio response to host size in the parasitoid wasp Spalangia endius.Behavioral Ecology and Sociobiology,1999,46(5):325-332.
[11] He F Q.Massive rearing of the houseflies.Beijing:Jin Dun Publishing House,2006:39-67.
[12] Propp G D,Morgan P B.Superparasitism of house fly,Musca domestica L.,pupae by Spalangia endius Walker(Hymenoptera:pteromalidae).Environmental Entomology,1983,12(2):561-566.
[13] Crawley M J.The R book.Chichester:John Wiley and Sons,2007.
[14] Li G Q,Mu L L.Competition among parasitoids for host during foraging and oviposition.Acta Ecologica Sinica,2006,26(4):1261-1269.
[15] King B H.Offspring sex ratios in parasitoid wasps.Quarterly Review of Biology,1987,62(4):367-396.
[16] Montoya P,Cancino J,Pérez-Lachaud G,Liedo P.Host size,superparasitism and sex ratio in mass-reared Diachasmimorpha longicaudata,a fruit fly parasitoid.Biocontrol,2011,56(1):11-17.
[17] Tormos J,Asís J,Sabater-Muñoz B,Baños L,Gayubo S F,Beitia F.Superparasitism in Laboratory rearing of Spalangia cameroni(Hymenoptera:Pteromalidae),a parasitoid of medfly(Diptera:Tephritidae).Bulletin of Entomological Research,2012,102(1):51-61.
[18] Lebreton S,Chevrier C,Darrouzet E.Sex allocation strategies in response to conspecifics′offspring sex ratio in solitary parasitoids.Behavioral Ecology,2009,21(1):107-112.
[19] Lacoume S,Bressac C,Chevrier C.Effect of host size on male fitness in the parasitoid wasp Dinarmus basalis.Journal of Insect Physiology,2006,52(3):249-254.
[20] Martel V,Darrouzet É,Boivin G.Phenotypic plasticity in the reproductive traits of a parasitoid.Journal of Insect Physiology,2011,57(6):682-687.
[21] Wang X Y,Yang Z Q,Liu G J,Liu E S.Relationships between the emergence and oviposition of ectoparasitoid Spathius agrili Yang and its host emerald ash borer,Agrilus planipennis Fairmaire.Acta Ecologica Sinica,2006,26(4):1103-1109.
[22] Liu Z,Xu B,Li L,Sun J.Host-size mediated trade-off in a parasitoid Sclerodermus harmandi.PLoS One,2011,6(8):e23260.
[23] Wang X Y,Yang Z Q,Wu H,Gould J R.Effects of host size on the sex ratio,clutch size,and size of adult Spathius agrili,an ectoparasitoid of emerald ash borer.Biological Control,2008,44(1):7-12.
[24] King B H,Bressac C.No fitness consequence of experimentally induced polyandry in a monandrous wasp.Behaviour,2010,147(1):85-102.
[25] Henter H J.Constrained sex allocation in a parasitoid due to variation in male quality.Journal of Evolutionary Biology,2004,17(4):886-896.
[26] Santolamazza-Carbone S,Nieto M P,Rivera A C.Maternal size and age affect offspring sex ratio in the solitary egg parasitoid Anaphes nitens.Entomologia Experimentalis et Applicata,2007,125(1):23-32.
[27] Chen Z Z,Liu J B,He Z,Duan B S,Hu H Y.Strategies of Pachycrepoideus vindemmiae parasitizing pupae of houseflies.Chinese Journal of Applied Entomology,2011,48(6):1765-1769.
[28] Hu H Y,Chen Z Z,Duan B S,Zheng J T,Zhang T X.Effects of female diet and age on offspring sex ratio of the solitary parasitoid Pachycrepoideus vindemmiae(Rondani,1875)(Hymenoptera,Pteromalidae).Revista Brasileira de Entomologia,2012,56(2):259-262..
[29] Petersen J J,Meyer J A,Stage D A,Morgan P B.Evaluation of sequential releases of Spalangia endius(Hymenoptera:Pteromalidae)for control of house flies and stable flies(Diptera:Muscidae)associated with confined livestock in eastern Nebraska.Journal of Economic Entomology,1983,76(2):283-286.
[30] Zhang Y P,Li D S,Zhang B X,Chen M Y,Zhong J,Song Y.Functional response of Spalangia endius Walker to pupae of Bactrocera dorsalis(Hendel)and influence of temperature and relative humidity on longevity of adult S.endius.Chinese Journal of Biological Control,2010,26(4):385-390.
参考文献:
[7] 张晓岚,孟玲,李保平.菜粉蝶蛹体型大小对蝶蛹金小蜂后代数量、性比及体型大小的影响.生态学杂志,2009,28(4):677-680.
[11] 何凤琴.家蝇规模化养殖.北京:金盾出版社,2006.39-67.
[14] 李国清,慕莉莉.拟寄生蜂搜索产卵过程中对寄主的竞争.生态学报,2006,26(4):1261-1269.
[21] 王小艺,杨忠岐,刘桂军,刘恩山.白蜡吉丁柄腹茧蜂的羽化和产卵与寄主之间的关系.生态学报,2006,26(4):1103-1109.
[27] 陈中正,刘继兵,贺张,段毕升,胡好远.蝇蛹金小蜂对家蝇蛹的寄生策略.应用昆虫学报,2011,48(6):1765-1769.
[30] 章玉苹,李敦松,张宝鑫,陈明洋,钟娟,宋月.蝇蛹俑小蜂对桔小实蝇蛹的功能反应及温湿度对蜂成虫寿命的影响.中国生物防治,2010,26(4):385-390.
猜你喜欢
西北农业学报(2022年7期)2022-07-22
昆虫学报(2021年5期)2021-07-07
奇闻怪事(2020年7期)2020-09-10
山东第一医科大学(山东省医学科学院)学报(2019年11期)2019-11-07
浙江大学学报(农业与生命科学版)(2019年4期)2019-09-19
河南农业科学(2018年6期)2018-01-18
西藏科技(2015年3期)2015-09-26
科学大众·小诺贝尔(2015年5期)2015-05-04
环境昆虫学报(2014年2期)2014-11-25
湖北科技学院学报(医学版)(2014年3期)2014-02-28
