
采伐对长白山阔叶红松林生态系统碳密度的影响
2013-12-16 08:18于大炮周旺明赵福安王长海代力民
生态学报 2013年10期
齐 麟,于大炮,周旺明,周 莉,赵福安,王长海,代力民,*
(1.森林与土壤生态国家重点实验室,中国科学院沈阳应用生态研究所,沈阳 110016;2.中国科学院研究生院,北京 100049;3.吉林森工集团露水河林业局,白山 134506)
森林作为陆地生态系统主要的碳库,占据着全球非冰陆地表面的40%,与其他生态系统相比,其分布面积最大,生产力和生物量积累最高,在全球的碳循环中发挥着重要的作用[1]。人类干扰对森林生态系统碳循环有重要影响[2]。研究表明,若植被完全被采伐并进行土地耕种,除地上碳全部损失以外,土壤1m深度内土壤有机碳将损失25%—30%,皆伐后进行耕作15a后土壤有机碳将损失70%[3-5]。若植被没有被完全采伐,则森林碳密度随采伐强度的变化而有所不同,高强度采伐不仅将导致植被组成和生物量的损失在短期内难以恢复,更会导致采伐迹地裸露面积增大,雨水冲刷严重,使土温升高,从而加速土壤中C/N元素的释放和流失[6];经适当强度采伐的林分,若其结构及功能的变化在稳定、平衡的生态系统阈值内,使既能保持系统的平衡,又能调节林内环境因子,有利于系统内的元素转化利用,促进林木的快速生长和生态系统碳汇功能的增强[7]。
长白山林区位于我国最大的东北林区。该区森林经营活动频繁,经过多年采伐利用,该区现存的森林主要是少部分的原始阔叶红松林和大部分破坏后恢复的天然次生林。森林经营活动直接使阔叶红松林植被的物种组成和数量产生变化,而对其植被和凋落物碳密度产生巨大影响[8-10],而植被的改变又会导致微气候的改变,进而影响到土壤碳密度[11-12]。为了明确森林经营活动对阔叶红松林生态系统各子碳库碳密度的影响,本研究选择长白山地区具有典型代表性的露水河林业局,基于该林业局经营管理资料和野外调查数据,以原始阔叶红松林和经采伐干扰后形成的次生林为研究对象,研究采伐对阔叶红松林植被碳库、枯死物碳库和土壤碳库的影响,并通过研究次生林植被碳密度采伐强度,恢复时间的相关关系,确定生态系统恢复、木材生产与固碳效益综合效益最大化的采伐强度与轮伐期,为制定合理的林业经营措施提供参考。
1 研究地区和研究方法
1.1 研究区域概况
本研究样地均位于露水河林业局(127°29'—128°02'E,42°20'—42°40'N)。该局位于长白山西北麓的抚松县境内,年平均气温4.5—7.8℃,年降水量800—1040 mm,海拔700—1000m。,土壤为山地暗棕色森林土,土壤剖面发育明显,平均深度50cm。该区地带性植被为阔叶红松林,1970年开始森林采伐,经营历史较短,经营措施记录清晰。迄今为止,既有未受干扰保护完好的原始阔叶红松林,又有受不同强度采伐,经不同恢复时间而形成的各种次生阔叶红松林。
1.2 数据采集
1.2.1 生态系统碳密度计算
本研究中森林生态系统碳密度指单位面积森林生态系统有机碳储量,它包括植物、凋落物和土壤碳库(不包括土壤动物碳储量)。
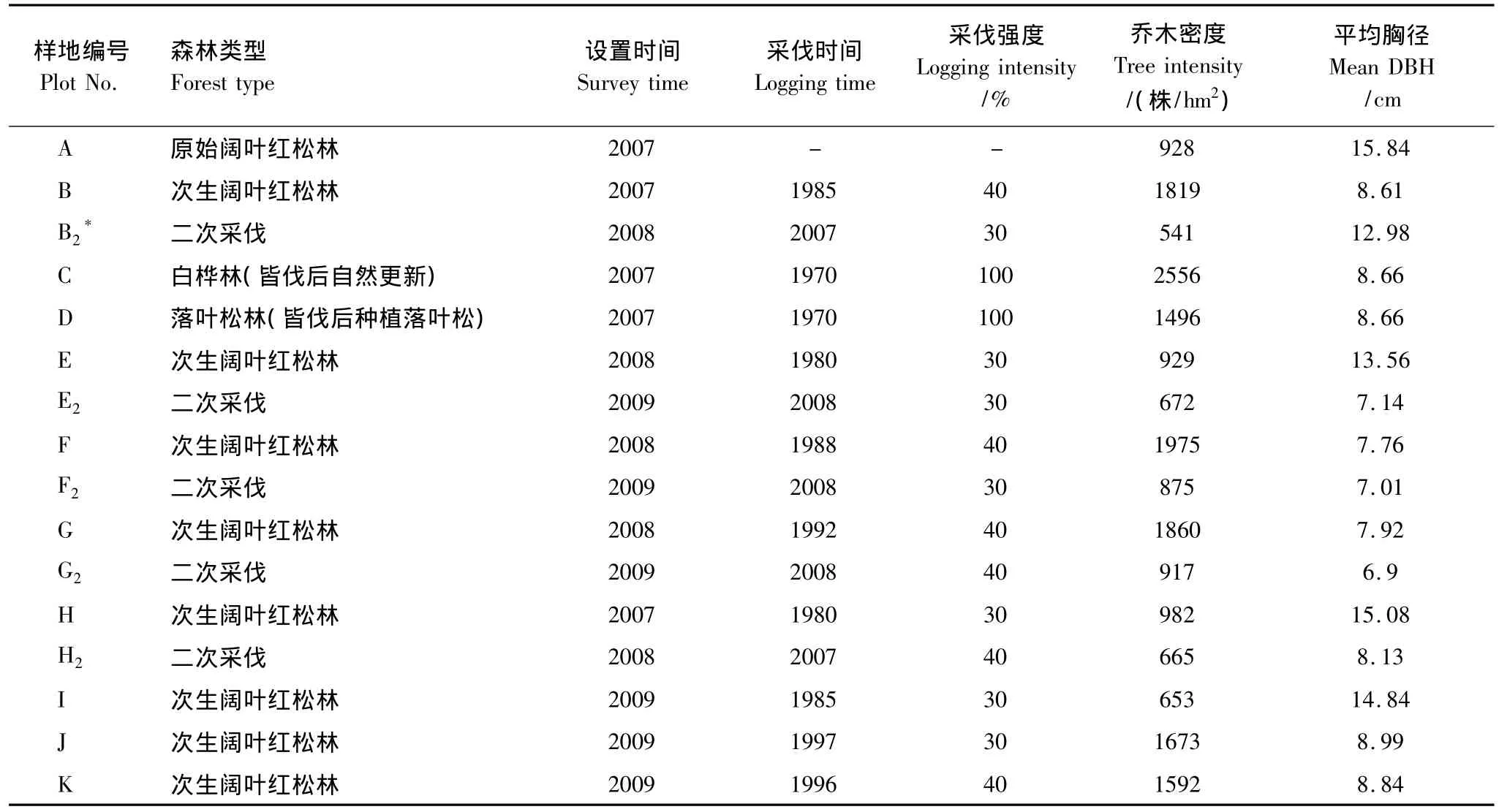
表1 研究样地情况介绍Table 1 General information of study sites
1.2.2 植被调查与碳密度计算
2007至2009年,在露水河林业局东升林场,设置11块100 m×100 m面积为1hm2调查样地,森林类型包括原始阔叶红松林、经过不同干扰强度、恢复时间以及不同恢复措施形成的次生林(表1)。将1 hm2样地分为25块20 m×20 m的小样方,详细调查、记录和标记DBH≥2cm的木本植物的种类、DBH和树高,将每个小样方植被碳密度作为一个重复,计算1hm2样地植被碳密度。
将2cm≤DBH<8cm作为下木层、8cm≤DBH<20cm作为次林层、DBH≥20cm作为主林层归类,并根据吉林省一元材积表将乔木分3类:针叶树、一类阔叶树和二类阔叶树。根据东北主要树种生物量异速生长方程提供的木本植物胸径与各器官生物量的关系与树种在表2中所属的种类计算样地中每株木本植物的生物量[13]。根据生物量与碳储量的转换系数[14-16]计算每株树木的碳储量(碳储量=生物量×0.5),再根据植被调查结果推算样地尺度植被碳密度。
1.2.3 凋落物碳密度计算
凋落物碳库包括所有地表枯枝落叶和粗木质残体两部分,在上述11块样地中沿样地对角线(东北-西南方向)的5个小样方中进行凋落物调查。凋落物碳密度计算:将凋落物分为基部直径大于10cm的粗木质残体和小于10cm的枯枝落叶。调查粗木质残体两头直径,长度和腐烂级,根据圆台体积公式和腐烂级与碳储量的转换系数[17]计算每个粗木质残体的碳储量。在每个小样方中,随机选择3块面积为1m×1m的样方,将所有样方内所有枯枝落叶装袋回实验室分析,根据烘干重和测定碳含量,计算凋落物碳密度。
1.2.4 土壤碳密度计算
在上述11块样地中每块样地对角线(东北—西南方向)的5个小样地中挖50cm深的土壤剖面,分为10 cm深的土壤共5层,用100 cm3的环刀在每层土壤中取土样1份,装袋回实验室分析。根据土壤容重和碳含量计算土壤碳密度。
在上述取样与调查中碳含量分析均采用常规测定方法[18]。
1.3 数据分析
采用EXCEL 2010,Origin 8.5和SPSS16.0软件进行数据统计分析和绘图,采用回归方法进行采伐强度、恢复时间与碳密度关系方程拟合,并以均方根误差(RMSE)和相对均方根误差(RMSEr)为指标进行拟合效果验证。采用单因素方差分析(One way ANOVA)和Duncan法进行差异性比较各样地碳密度。
2 结果与分析
2.1 样地植被、凋落物、土壤和生态系统碳密度
在11块样地中,原始阔叶红松林(A)的植被层碳密度最大,为152.87 MgC/hm2,显著高于其他各样地(P<0.01);采伐时间距调查时间较短的样地碳密度最小,值为64.52 MgC/hm2(B2),77.4 MgC/hm2(E2),62.21 MgC/hm2(F2),45.14 MgC/hm2(G2)和 60.18 MgC/hm2(H2),显著低于其他各样地(P <0.01)(表2)。

表2 各样地生态系统碳密度Table 2 Carbon density in 11 sites

图1 不同类型森林各径级乔木碳密度Fig.1 Carbon stocks of forest each diameter class in three types of forest
原始阔叶红松林植被碳库主要集中于主林层(DBH≥20cm)乔木层中,其中下木层(DBH<8cm)、次林层(8cm≤DBH<20cm)、主林层乔木碳密度分别占植被碳密度的1.73%、6.87%和91.4%。白桦林与落叶松林中次林层与较主林层乔木碳密度相差不大(白桦林:林下层占 2.90%,次林层占 41.52%,主林层占55.57%;落叶松林:林下层占 2.46%,次林层占31.26%,主林层占66.27%)(图1)。皆伐后次生阔叶红松林中林下层乔木碳密度所占植被碳密度的比例要高于原始阔叶红松林,而主林层乔木碳密度所占植被碳密度的比例远低于原始阔叶红松林(表2)。
在11块调查样地中,凋落物碳密度值显著分为3个水平,D > A、B、E、F、G、H、I、J、K > C,即落叶松林 >阔叶红松林>白桦林(P<0.01)(表2)。各样地凋落物碳密度值与采伐强度和恢复时间无显著相关关系(P>0.05)。
各样地中的土壤碳密度均值在100—160 MgC/hm2之间,且无显著差异(P>0.05)。原始林样地中的土壤碳密度与植被碳密度相当。而样地B、C、D、F、G、H、I、J和K的土壤层碳密度均大于样地植被层碳密度,即经采伐干扰对样地植被碳密度的影响要高于对照样地中土壤层碳密度的影响(表2)。各样地的植被、凋落物和土壤的碳密度汇总得到生态系统碳密度。原始林样地的生态系统碳密度最大,其余各经采伐干扰的样地的生态系统碳密度均小于原始林样地(表2)。
2.2 采伐强度与恢复时间对植被碳密度的影响
采伐干扰对样地植被碳密度的影响规律性较强,即采伐强度越小,植被恢复时间越长(采伐时间越早),调查时样地的植被层碳密度越大(图2)。其中采伐强度与植被碳密度呈显著的线性负相关关系(R2=0.726,P<0.05)。植被层碳密度则随着时间呈显著线性增长(图2)。
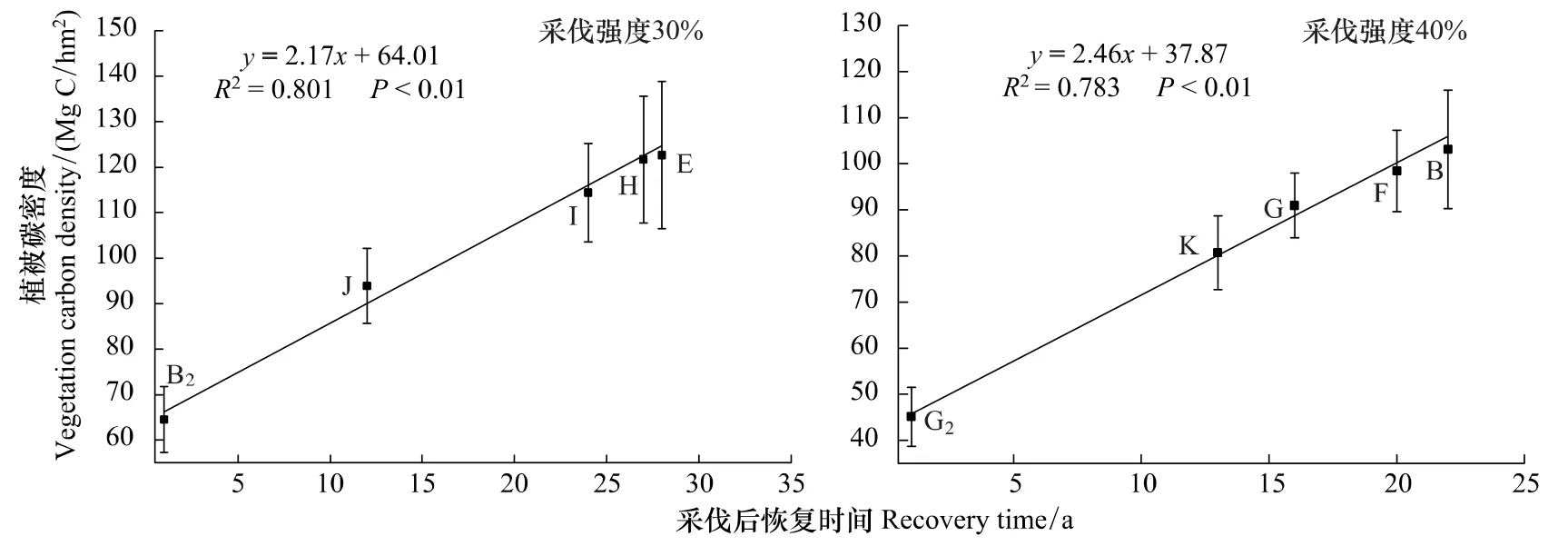
图2 恢复时间与植被碳密度相关关系Fig.2 Relationship between vegetation carbon density and recovery time
经采伐形成的次生阔叶红松林中不同径级乔木碳密度随恢复的时间而发生改变。林下层乔木碳密度随着恢复时间的增加而降低,主林层乔木碳密度随着恢复时间的增加而增加,次林层乔木碳密度随时间的增加变化不大(图4)。
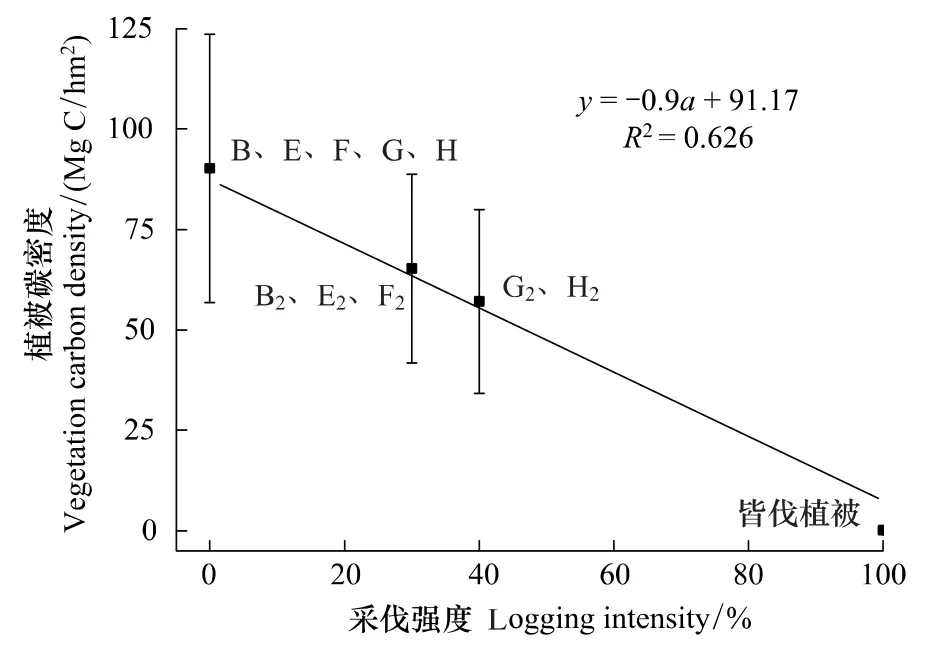
图3 碳密度与采伐强度相关关系Fig.3 Relationship between carbon density and logging intensity
2.3 采伐对土壤碳密度的影响
各样地的土壤碳密度均随深度的增加而减少,不同类型样地在土壤表层(0—10cm和10—20cm)的碳密度有较大差异:在0—10cm层,原始阔叶红松林土壤碳密度最高,为65.18 MgC/hm2,落叶松林土壤碳密度最低,为51.48 MgC/hm2;在10—20cm层,原始阔叶红松林土壤碳密度最高,为49,01 MgC/hm2择伐后的从次生林最低,为36.22 MgC/hm2。20—50cm 各层中不同类型样地土壤碳密度差异很小(图5)。
经过采伐干扰的次生林土壤碳密度均低于原始阔叶红松林土壤碳密度。其中恢复时间较长的白桦林与落叶松林土壤碳密度与原始阔叶红松林土壤碳密度较为接近(表1)。各择伐后次生林样地土壤碳密度与采伐强度无显著相关关系(R2= -0.073,P>0.05),但与恢复年限有显著相关性(R2=0.847,P<0.01)(图3)。

图4 不同径级植被碳密度变化与的关系Fig.4 Vegetation carbon stocks changes of different diameter classes along recovery time
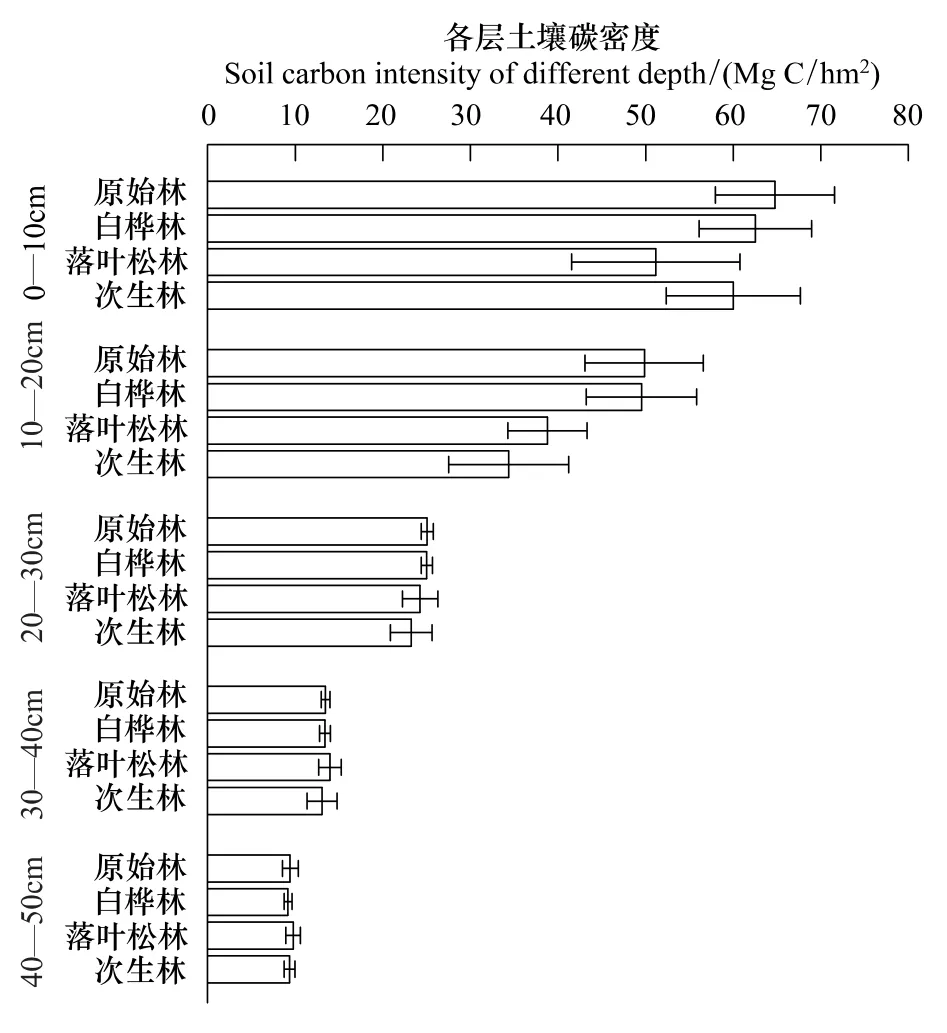
图5 各类型样地土壤碳密度垂直分布Fig.5 Vertical distribution of soil organic carbon in each type of site
3 讨论
3.1 阔叶红松林生态系统碳密度特征
作为东北地区典型森林生态系统之一,阔叶红松林生态系统有着较高的碳密度。在本研究中,11块样地的平均地上碳密度为98.02 MgC/hm2,高于全国植被平均碳密度 57.07 MgC/hm2[10]41.32 MgC/hm2[19]、41.00 MgC/hm2[20]的估计值,低于李文华等人对阔叶红松林地上碳密度134.50 MgC/hm2的估算[21]。唐凤德等人通过(Sim-cycle)模型,估算长白山原始阔叶红松林植被碳密度182.5 MgC/hm2,与本研究中原始阔叶红松林植被碳密度相当[22],而凋落物碳密度为8.7 MgC/hm2,仅为本研究中凋落物碳密度21.1 MgC/hm2的一半。土壤碳密度为358.3 MgC/hm2,远高于本研究原始林土壤碳密度157.1 MgC/hm2。闫平等人对阔叶红松林土壤碳密度的研究结果为220.30 MgC/hm2,与本研究相近[23]。造成这种差异的原因可能是:(1)研究方法,本研究与闫平等人的研究均为在样地调查的基础上得出,而唐等人的结果为模型推算,且与本研究的研究尺度不同。(2)生态系统起源不同,原始林在地上部分达到碳平衡后,土壤碳储量依旧上升,因此,不同林龄的同一类型的原始林在土壤碳储量上也可能存在较大差异。(3)样地本底异质性,土壤碳储量不仅仅受到地上植被的影响,土壤发育,地形的因素也会对土壤碳密度造成加大影响,因此,由于研究区域的不同也会造成研究结果的较大差异。
在原始的阔叶红松林中,植被碳密度与土壤碳密度相当,两者占生态系统碳密度的90%以上。其余各次生林的植被碳库与土壤碳库均小于原始林,说明采伐造成了植被碳库与土壤碳库的流失,而土壤碳库的变化范围低于植被碳库,说明采伐对植被碳库的影响要高于对土壤碳库的影响(表3)。
3.2 经营方式对阔叶红松林植被、凋落物、土壤碳密度的影响
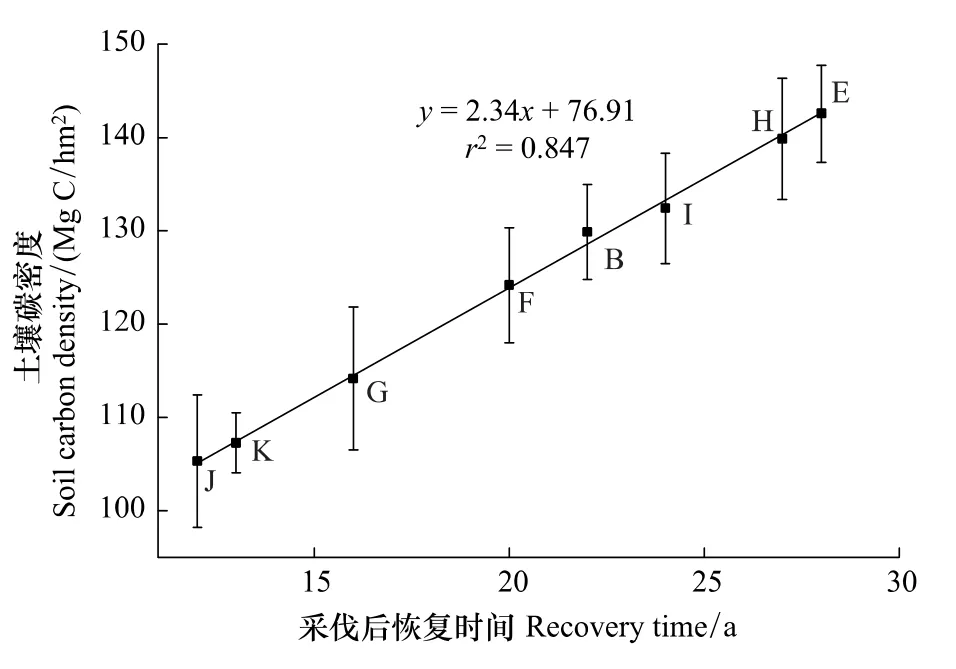
图6 土壤碳密度与采伐后恢复时间的相关关系Fig.6 Relationship between soil carbon density and recovery time
采伐直接导致植被碳库碳密度减少,且采伐方式与强度直接影响植被碳库减少的程度(图2,图3),由于采伐所选择的乔木皆为主林层中的乔木,所以采伐后主林层乔木碳密度占植被总植被碳密度的比例也随之降低。由于主林层乔木的去除,森林中乔木竞争压力的减少,林下层乔木生长旺盛,故其碳密度所占比例要高于原始阔叶红松林。随着恢复时间的增加,林下层乔木生长至次林层,而次林层乔木生长至主林层,主林层乔木碳密度所占的比例增加,而由于竞争压力的增加,小径级乔木碳密度所占的比例下降(图4)。另一方面,采伐后,恢复的方式对森林群落更新和演替有重要的影响,恢复的乔木种类直接影响到植被碳密度恢复的速度。在本研究中,C、D两块样地尽管是皆伐,但因其恢复树种是白桦和落叶松这样的速生先锋物种,其植被碳密度在30a的增量远远高于择伐后自然恢复的次生阔叶红松林30a的植被碳密度增量(表2)。总体来说,采伐导致了森林生物量的下降,而在较长时间尺度上,植被生物量应该呈逻辑斯蒂增长,植被碳密度得变化也应该与之相符,而本研究中各样地植被碳密度随时间呈线性增长,是本研究的时间跨度为40a左右,植被碳密度尚未达到顶级群落的稳定状态所致。
森林经营对生态系统凋落物碳密度的影响体现在采伐方式和更新方式的不同上。不同的采伐方式和更新方式,导致了凋落物的组成发生了变化,其中择伐没有使凋落物的种类和组成发生改变,皆伐后天然更新使阔叶凋落物(主要是白桦落叶)占据了凋落物的主要储量,而皆伐后人工更新的落叶松林其凋落物的主要组成为针叶。凋落物的种类与组成不同,其C/N也不同,周转速率也不同。阔叶林中凋落物的C/N比较低,因此周转较快,针叶凋落物C/N较高,周转较慢。在本研究中,落叶松林凋落物碳库碳密度远高于择伐后次生林中凋落物碳密度,而采伐强度对次生阔叶红松林生态系统凋落物碳密度的影响并不明显。
森林经营对阔叶红松林生态系统土壤碳库的影响较为复杂,一方面,由于采伐改变了林内植被的组成和微气候,进而改变了土壤微生物的组成与数量,改变了土壤碳循环,使土壤碳库碳密度发生变化。落叶松林中因凋落物难以分解,因此,有大量的有机碳储存在凋落物层中而向表层土壤输入的有机碳较少,导致了土壤表层有机碳低于原始阔叶红松林。另一方面,采伐作业本身也对土壤表层理化及生物结构造成了巨大的破坏,导致了在次生阔叶红松林中表层土壤碳密度远低于其他样地。在本研究中,不同经营方式均导致了土壤碳的流失,且影响的垂直深度在20cm左右,即森林经营对土壤表层碳密度影响较大。土壤碳密度与恢复时间相关性分析的起点是12a,而相关分析的结果是线性正相关说明了样地土壤有机碳流失过程的时间少于12a,且流失的速度远大于积累的速度。
3.3 采伐强度、轮伐期与植被碳密度
由植被碳密度与采伐后恢复时间的相关关系(图1,图2)可以计算出,当采伐强度为40%时,植被碳密度恢复到原始阔叶红松林碳密度150MgC/hm2的水平大约需要55a,而采伐强度为30%时,则只需要45a。由此可以确定,当采伐强度为40%时,合理的轮伐期应为55a,而当采伐强度为30%时,合理的轮伐期应为45a。而从有利于森林固碳的采伐经营来看,30%的采伐强度可以减小采伐时带来的植被碳密度损失。另外,从各径级乔木碳密度随恢复时间变化的规律来看,30%采伐强度后,大径级乔木碳密度的恢复速度要远高于40%采伐(图4)。
对于同一个阔叶红松林生态系统,如采用40%的采伐强度,55a的轮伐期,170a左右生态系统经3次采伐后植被碳密度均重新恢复到原始阔叶红松林水平,生产了原始阔叶红松林3×40%蓄积量的木材(图3)。而如果采用30%的采伐强度,仅在180a中则就会经历4次采伐并恢复到原始状态,并会生产原始阔叶红松林蓄积4×30%木材。与同一时间中采伐强度为40%的采伐方式生产的木材数量相当。
4 结论
综上所述,择伐、皆伐以及皆伐后不同的更新方式都对阔叶红松林生态系统中植被、凋落物以及土壤碳密度产生了不同的、深远的影响。无论从生产力还是从固碳与生态恢复角度,较低的择伐强度,较长的伐期都是有利于阔叶红松林可持续经营的采伐方式。
[1] Zhao S D,Wang Y X,Forest and carbon cycle.Impact of Science on Society,2001,03:38-41.
[2] Dixon R K,Solomon A M,Brown S,Houghton R A,Trexier M C,Wisniewski J.Carbon pools and flux of global forest ecosystems.Science,1994,263(5144):185.
[3] Johnson D W.Effects of forest management on soil carbon storage.Water,Air,& Soil Pollution,1992,64(1):83-120.
[4] Lee J,Morrison I K,Leblanc J D,Dumas M T,Cameron D A.Carbon sequestration in trees and regrowth vegetation as affected by clearcut and partial cut harvesting in a second-growth boreal mixedwood.Forest Ecology and Management,2002,169(1/2):83-101.
[5] Saari P,Saarnio S,Kukkonen J V K,Akkanen J,Heinonen J,Saari V,Alm J.DOC and N2O dynamics in upland and peatland forest soils after clearcutting and soil preparation.Biogeochemistry,2009,94(3):217-231.
[6] Johnson D W,Curtis P S.Effects of forest management on soil C and N storage:meta-analysis.Forest Ecology and Management,2001,140(2/3):227-238.
[7] Mazzei L,Sist P,Ruschel A,Putz F E,Marco P,Pena W Ferreira J E.Above-ground biomass dynamics after reduced-impact logging in the Eastern Amazon.Forest Ecology and Management,2010,259(3):367-373.
[8] Fang J Y,Chen P A.Dynamic Forest Biomass Carbon Pools in China and Their Significance.Acta Botanica Sinica,2001,09:967-973.
[9] Yang L Y,Luo T,Wu S T.Fine root biomass and its depth distribution across the primitive Korean pine and broad-leaved forest and its secondary forests in Changbai Mountain,northeast China.Acta Ecologica Sinica,2007,27(9):3609-3618.
[10] Zhou Y R,Yu Z L,Zhao S D.Carbon storage and budget of major chinese forest types.Acta Phytoecologica Sinica,2000,05:518-522.
[11] Yang L Y,Dai L M.Storage and decomposition of fallen wood in dark coniferous forest on the North Slope of Changbai Mountain.Chinese Journal of Applied Ecology,2002,13(9):1069-1071.
[12] Zhou L,Dai L M,Gu H Y,Yu D P.Dynamics of soil nutrient contents in cutting forestlands of broad-leaved Korean pine forest on Changbai Mountains.Chinese Journal of Applied Ecology,2004,15(10):1771-1775.
[13] Jilin Forestry Department,Standing timber volume table of Jilin.Changchun:Jilin Forestry Department.2003:185-192.
[14] Guan N,Chen C.wood density and cutting resistance of Korean pine and Fraxinus mandshurica.Scientia Silvae Sinicae,1990,26(2):149-155.
[15] Guan N.11 Coniferous wood density and cutting resistance.Scientia Silvae Sinicae,1991,27(006):630-638.
[16] Guan N.15 Stella material in the cutting thickness,rake angle and moisture content of wood cutting resistance.Scientia Silvae Sinicae,1994,30(2):134-139.
[17] Gu H Y,Dai L M.Effect of Human Disturbance on Coarse Woody Debris in Korean Pine and Broad-Leaved Mixed Forest on Changbai Mountain.,2006,42(10):1-5.
[18] Liu G S.Soil physical and chemical analysis and profile description.1996.Beijin:Standards Press of China.
[19] Zhao M,Zhou G S.Carbon Storage of Forest Vegetation and Its Relationship with Climatic Factors.Scientia Geographica Sinica,2004,24(1):50-54.
[20] Fang J Y,Guo Z D,Piao S L.1981— 2000 Chinese terrestrial vegetation carbon sinks estimating.Science in China(Series D:Earth Sciences),2007,37(6):804-812.
[21] Li W H,Deng K M,Li F.Changbai Mountain ecosystem biomass production.Forest Ecosystem Research,1981,2(34):22-24.
[22] Tang F D,Han S J,Zhang J H.Carbon dynamics of broad-leaved Korean pine forest ecosystem in Changbai Mountains and its responses to climate change.Chinese Journal of Applied Ecology,2009,20(6):1285-1292
[23] Yan P,Feng X C.Spatial Distribution and Carbon Storage in Primitive Broadleaved Korean Pine Forests.Journal of Northeast Forestry University,2006,34(005):23-25.
参考文献:
[1] 赵士洞,汪业勖,森林与碳循环.科学对社会的影响,2001,03:38-41.
[8] 方精云,陈安平,中国森林植被碳库的动态变化及其意义.植物学报,2001,09:967-973.
[9] 杨丽韫,罗天祥,罗松涛.长白山原始阔叶红松(Pinus koraiensis)林及其次生林细根生物量与垂直分布特征.生态学报,2007,27(9):3609-3618.
[10] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡.植物生态学报,2000,05:518-522.
[11] 杨丽韫,代力民.长白山北坡暗针叶林倒木贮量和分解的研究.应用生态学报,2002,13(9):1069-1071.
[12] 周莉,代力民,谷会岩,于大炮.长白山阔叶红松林采伐迹地土壤养分含量动态研究.应用生态学报,2004,15(10):1771-1775.
[13] 吉林省林业厅.吉林省一元立木材积表.长春:吉林省林业厅,2003:185-192.
[14] 管宁,陈森.红松水曲柳木材密度与切削阻力关系的研究.林业科学,1990,26(2):149-155.
[15] 管宁.11种针叶树木材密度与切削阻力关系的研究.林业科学,1991,27(006):630-638.
[16] 管宁.15种叶树材中切削厚度,刀具前角和木材含水率对切削阻力的影响.林业科学,1994.30(2):134-139.
[17] 谷会岩,代力民.人为干扰对长白山红松针阔叶混交林粗木质残体的影响.林业科学,2006,42(10):1-5.
[18] 刘光崧.土壤理化分析与剖面描述.1996:中国标准出版社.
[19] 赵敏,周广胜.中国森林生态系统的植物碳贮量及其影响因子分析.地理科学,2004,24(1):50-54.
[20] 方精云,郭兆迪,朴世龙,陈平安.1981—2000年中国陆地植被碳汇的估算.中国科学:D辑,2007,37(6):804-812.
[21] 李文华,邓坤枚,李飞.长白山主要生态系统生物量生产量的研究.森林生态系统研究,1981,2(34):22-24.
[22] 唐凤德,韩士杰,张军辉.长白山阔叶红松林生态系统碳动态及其对气候变化的响应.应用生态学报,2009,20(6):1285-1292.
[23] 闫平,冯晓川.原始阔叶红松林碳素储量及空间分布.东北林业大学学报,2006,34(005):23-25.
猜你喜欢
北京林业大学学报(社会科学版)(2022年4期)2023-01-04
建筑与预算(2022年6期)2022-07-08
小哥白尼(野生动物)(2021年9期)2022-01-17
绿色科技(2020年20期)2020-11-20
中国林副特产(2020年5期)2020-10-09
防护林科技(2017年7期)2017-09-01
江苏农业科学(2017年6期)2017-05-11
林业勘查设计(2017年1期)2017-03-11
前卫文学(2016年3期)2016-07-01
国际木业(2016年6期)2016-02-28
