
蝗虫的聚集行为机制研究进展
2014-08-10 12:29封传红陈淑群张跃进刘万才罗林明
植物保护 2014年1期
封传红,陈淑群,郭 聪,张跃进,刘万才,罗林明
(1.四川省农业厅植物保护站,成都 610041;2.四川省仁寿县植物保护站,仁寿 620500;3.四川大学生命科学学院,成都 610064;4.全国农业技术推广服务中心,北京 100125)
蝗虫的聚集行为机制研究进展
封传红1,陈淑群2,郭 聪3,张跃进4,刘万才4,罗林明1
(1.四川省农业厅植物保护站,成都 610041;2.四川省仁寿县植物保护站,仁寿 620500;3.四川大学生命科学学院,成都 610064;4.全国农业技术推广服务中心,北京 100125)
蝗虫具有聚集为害的特征。群居型蝗虫形态、生理、生化、行为等与散居型明显不同。蝗虫的密度、资源的空间分布方式、化学信息素和特定行为均能够引起蝗虫聚集行为反应。蝗群聚集后,通过遵守简单的行为规则,形成了一个高度有序的复杂系统,在这个有序的复杂系统里群体蝗虫能够取得散居个体不能获取的生态效益。
蝗虫; 聚集; 序; 效益
蝗虫灾害与水、旱灾害常此起彼伏,交替发生,一直是严重威胁农业生产、影响人民生活和社会安定的三大自然灾害。世界上对农业生产危害最大的种类是沙漠蝗[Schistocercagregaria(Forska)]和飞蝗(LocustamigratoriaL.)。造成我国农牧业大面积、毁灭性灾害的主要是飞蝗,近年来亚洲小车蝗(OedaleusasiaticusB.-Bienko)、意大利蝗[Calliptamusitalicus(L.)]等在北方农牧交错区发生为害严重[1-2]。历史上由于蝗虫的为害,常造成“飞蝗蔽天,田禾尽损”、“饿殍枕道,人饥相食”现象[3]。蝗虫为害的严重性的重要原因之一是蝗虫具有聚集扩散和迁移习性,能将分散的为害集变为灾害。研究蝗虫聚集的生理生态基础、影响因子、聚集蝗虫序的形成和聚集的生态效益,尤其是蝗虫的聚集行为、聚集行为的生理生化背景和蝗虫群体行为规律、蝗虫从单一散乱的单个蝗虫聚变为行为一致、破坏力极强的蝗虫军团的灾变机制等,对于破解飞蝗聚集行为这一世界性难题具有重要意义。
1 蝗虫聚集的生理生态基础
1.1 群居型、散居型
许多昆虫均有不同的生态型,其中以飞蝗、沙漠蝗等型变现象比较典型。飞蝗具有群居型、散居型以及介于两者之间的中间型(过渡型)。群居型和散居型在形态特征、生理机能、行为及体色等方面存在明显差异[4]。有群居生活史的飞蝗为群居型;整个生活史都散居的飞蝗为散居型[5]。群居型和散居型具有不同的外部特征,群居型体色一般较深,飞蝗群居型为黑色,散居型有绿色、黄褐色、褐色、甚至品红色等颜色;群居型前胸背板略短,沟前区明显缩狭,沟后区较宽平,形成鞍状,侧面看,中隆线较平直或在中部微凹;散居型前胸背板稍长,沟前区不明显缩狭,沟后区略高,不呈鞍状,侧面看,中隆线呈弧状隆起,如屋脊形;此外在E/F(前翅长度/后足腿节长度)、F/C(后足腿节长度/头部最宽度)的比值上,不同龄期群居型、散居型也有较大的差异[6-7]。
1.2 发育及生理学
在蝗蝻发育历期上,总的趋势是散居型的发育时间比群居型的短。在耐寒性方面,群居型成虫的冰冻点及过冷却点都高于散居型,两者差异显著。但同一型雌、雄之间差异不显著。脂肪含量方面,东亚飞蝗群居型的脂肪含量比散居型的高,雄性又略高于雌性。含水量与脂肪含量相反,即不论雌蝗或雄蝗,群居型体内水分含量均低于散居型,雌、雄之间差异也不显著。生殖力方面,群居型卵巢管数量多于散居型,而产卵数量散居型高于群居型。另外,散居型飞蝗的锥形感受器和腔锥形感受器都比群居型飞蝗的多[7]。
1.3 生态型与聚集行为
聚集行为可能发生在群居型蝗虫,也可能发生在散居型蝗虫中。自然条件下,聚集行为是区分群居型蝗虫的必要条件,散居型可以终生没有聚集行为。群居型与散居型在行为上存在明显的差异。群居型总是寻求同类聚集而居,而散居型则躲避同类,行孤单生活;群居型经常在白天飞行进行迁飞扩散,而散居型则要到夜晚才偶尔飞行[8]。东亚飞蝗无论是雄性或雌性蝗虫,同一龄期不同生态型的转向次数和跳跃频率,均是群居型极显著地大于散居型,说明群居型处于一种更为活跃的状态[9]。群居型沙漠蝗会接受一种含天仙胺成分的食物,天仙胺是植物里含有的一种可以防止植物被天敌取食的生物碱,群居型沙漠蝗在取食含天仙胺类食物并不需要特别的代价(如自身中毒等),而散居型没有类似现象[10]。群居型这种行为的生态意义在于:通过取食含有有毒成分的植物从而获得了散居型不具备的对抗天敌的功能,因为天敌通过体色就可以分辨出群居型和散居型[11-12]。
2 影响蝗虫聚集的因子
2.1 蝗虫密度
虫口密度对飞蝗型变的影响是普遍认同的,实验验证了飞蝗的散居型、中间型和群居型3种形态之间可随其密度的增减而相互转变[13]。沙漠蝗散居型个体在相互接触几个小时后就可以表现出群居行为,它们不再回避同类个体[10]。初孵若虫的饲养密度可以影响高龄蝗蝻的体色,密度越高,蝗蝻的黑色越浓[14]。在笼养的条件下,密度为6头/m2是3龄飞蝗由群居型向散居型转变的临界密度,并且如果群居时密度越大,群居型蝗蝻越难转变为散居型。3龄期可能是东亚飞蝗由群居型向散居型转变的临界期。超过3龄,即使在低密度下也不易发生生态型转变[15]。
2.2 资源的空间分布
食物的空间分布对蝗虫的两型分化有重要影响,研究表明,寄主的聚集度越高,沙漠蝗转换成群居型的可能性就越大[16]。资源空间分布的分形维数与蝗虫型变具有正相关,食物分形维数与蝗虫聚集度关系式为:y=0.40+0.17x(R2=0.8,蝗虫数为10头。x为食物分形维数,y为聚集度,下同)或y=0.49+0.16x(R2=0.95,蝗虫数为20头),按分形原理,分形维数高表明食物聚集度高[17]。食物成团分布,蝗虫相互接触而行为节律同步,引起生理等系列变化,有利于群居型的产生[18];对卫星影像分析表明,大尺度的地貌分形维数可以影响到蝗群的总体密度和迁飞[19]。有研究认为,植物群落组成甚至能够作为预测沙漠蝗分布的依据,特别是在那些地处偏远的地方,可以利用沙漠蝗寄主的分布和生长情况来预测沙漠蝗的风险区域和监控管理[20]。
2.3 化学信息素
蝗虫粪便、虫体、卵囊及其泡沫等都是聚集信息素的来源。粪便挥发物对东亚飞蝗蝗蝻的聚集作用表明:蝗虫粪便挥发物对各龄蝗蝻都有一定的引诱作用和滞留作用。其中高龄蝗蝻粪便挥发物对同龄和低龄蝗蝻的聚集作用较好。东亚飞蝗蝗蝻的粪便挥发物与体表挥发物组分相同,主要有醇类、醛类、环烯类、酮类,还有少量的酚类[21]。沙漠蝗蝗蝻粪便挥发物中分离出愈创木酚(guaiacol)和苯酚(phenol),具有强烈的聚集作用[22]。引起沙漠蝗蝗蝻聚集的体表挥化物为己醛(hexanal)、辛醛(octanal)、壬醛(nonanal)、癸醛(decanal)、己酸(hexanoic acid)、辛酸(octanoic acid)、壬酸(nonanoic acid)及癸酸(decanoic acid),成虫为苯甲醚(anisole)、苯甲醛(benzaldehyde)、藜芦醚(veratrole)、愈创木酚(guaiacol)、苯乙腈(phenylacetonitrile)及苯酚(phenol)[23-24]。卵囊及其泡沫主要影响蝗虫对产卵环境的选择。由上可以看出,聚集素不是单一物质,而是一系列的物质谱,不同虫态信息素谱是不同的[25]。
2.4 行为调节
研究人员将2~3龄的沙漠蝗放入同种群体或者非洲飞蝗[L.migratoriamigratorioides(ReicheetFairmaire)]群体中,观察视觉、触觉和嗅觉对型变的影响。结果表明,型变的发生并没有亚种的特异性,同非洲飞蝗种群一起饲养的沙漠蝗完全表现出群居行为。触觉可以引起型变,视觉和嗅觉共同起作用引起型变,单独嗅觉刺激对型变是没有效果的[26]。散居型沙漠蝗的后腿股节前段(外端)1/4的部位在受到4 h重复的触觉刺激后,散居型会表现群居型行为,散居型的这一刺激部位存在大量的毛发状触觉感受器,其数量要比群居型多大约30%,但后腿的其他部位感受器则比群居型少[27]。
3 聚集蝗虫序的形成
3.1 蝗虫聚集行为形成
在行为、内分泌、聚集信息、食物等因素的作用下,蝗虫聚集成团。特别在资源变得缺乏时,它们就被迫相互影响,调整各自的行为,群体稳定时可以组成有秩序的蝗群,行动趋向一致,随着蝗群迁移,越来越多的蝗虫加入进来,包括群居型、散居型,甚至其他蝗虫,随着蝗虫的加入,聚集的强度越来越大,形成了一种正向反馈机制,滚雪球式的最终成为了巨大的蝗群[10]。一个有意思的现象是蝗群在由零星蝗虫汇聚成大型蝗群时,是经历了一个由紊乱无序状态到组织有序的过程。散居型蝗虫行动没有规律性可言,它们可以看作随机游走的粒子,随着密度的增加,各个蝗虫相互影响,当达到一定密度时,整个蝗群行动的方向、速度高度统一,即使遇到地形、人为干扰,也会很快重新达到有序状态。
3.2 蝗群运动复杂系统
群居不是蝗虫独特的现象,蚂蚁、蜜蜂、鱼群等都有群居习性。近年来,研究的一个热点是动物群居后整齐划一行为是如何产生的,系统科学的一些理论被用来解释群居现象。研究人员认为,群居动物是一个自组织系统,群居内单个动物通过一些简单的重复行为,就能使整个群体表现出复杂的结构和功能[28]。物理学家已经发现许多自组织现象,如贝纳德(Bé nard)流体的对流花纹,贝洛索夫-扎鲍廷斯基(Belousov-Zhabotinsky)化学振荡花纹与化学波,激光器中的自激振荡等。生物学可以借鉴其原理和方法来分析生物界自组织现象。对于鱼群,计算机模拟表明单个的鱼遵守以下3条规则:(1)分隔规则:尽量避免与临近伙伴过于拥挤;(2)对准规则:尽量与临近伙伴的平均方向一致;(3)内聚规则:尽量朝临近伙伴的中心移动,即可形成高度有序的鱼群,表现出转圈、平行前行等多种行为(见图1)[29-30]。对于蝗虫来说,可能通过遵守类似简单规则,如与临近蝗虫运动保持一致,实现蝗群从无序到高度有序的转变[31]。
3.3 序的维系机制
从昆虫行为学看,蝗群要想获得高度有序状态,可能有3种信息反馈机制:(1)视觉。动物脑90%以上的信息来自视觉,昆虫的视力一般来说不及高等动物,因为昆虫的复眼没有调焦能力,视距大多在1 m以内,但复眼对移动物体感受力极强,如捕食性昆虫尤其是蜻蜓,在捕捉猎物时,能够根据猎物移动的速度、距离,准确判断自己所需的加速度以及与猎物相遇的瞬间,完成捕捉动作。蝗虫的复眼存在一套调节机制,使蝗虫移动与背景移动相对速度达到最小值,通过这种机制,能够使蝗群避免在移动或飞行中相互碰撞[32]。(2)嗅觉。每只蝗虫都可以散发聚集信息素,高密度的蝗群信息源交叉重叠,作为行动的指向,难以使各个蝗虫达到步调一致的效果。(3)触觉。田间观察中,在蝗虫聚集初期蝗虫抱团打转时,单个蝗虫之间有相互接触现象,随着迁移的进行,个体接触不多。蝗虫密度过高时可以见到大团蝗虫,如果环境比较宽敞,蝗群会自动调整到一个适中密度。因此,蝗虫维持有序信息的为视觉和触觉调节。行为调节后如何反映到整个群体的协调运动,这是研究中较少涉及的领域。
3.4 有序蝗群的领导者
领导者对于群体的作用是非常明显的,领导者的先验作用对于群体可以节约许多资源。金体美洲鳊鱼是一种群集度高的鱼类,试验证实,具有取食经验的少数个体(如有经验与无经验的比例为1∶11)能够带领鱼群找到食物资源,当然,比例越高,成功率越高[33]。蝗群是否存在领导者则不得知,但在田间可以发现,后加入蝗群的,会跟随整个蝗群运动而行动。然而,对于蟑螂的研究表明,蟑螂群体在选择隐蔽场所时,是由相互平等的、没有先验、没有领导的个体,通过非线性的动态的抉择,在这种抉择机制下,群体中的大多个体会获得一个较为理想的利益[34]。模型研究以上两种情况都有其合理性[35-36]。同为昆虫的蝗虫,是否存在类似蟑螂的抉择机制,有待观察和研究。
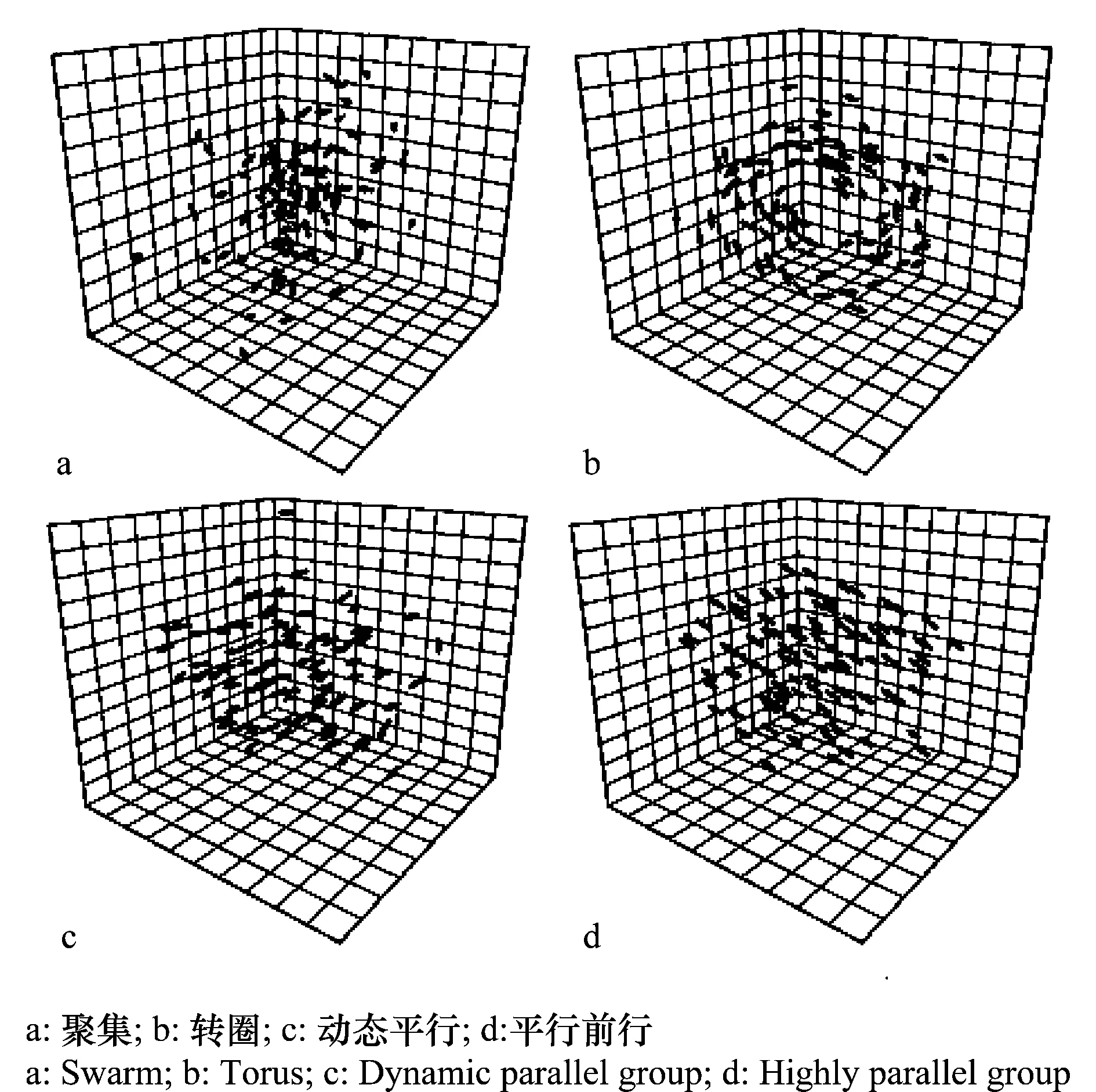
图1 模型鱼群行为(Iain D.Couzinn et al,2002)Fig.1 Collective behaviours of fishes exhibited by the model
4 聚集的生态效益
蝗虫通过聚集后,似乎获得散居型蝗虫不具备的行为。试验已经发现,群居型沙漠蝗在迁移途中遇到水渠,沙漠蝗会从水底爬过,水底穿行的过程可以维持9 min之久[37]。另一个现象是为了避免飞入海中,迁飞的蝗虫存在一种导向机制。2004年11月21—23日,观察到一个来自西奈半岛沙漠向东南飞行的蝗群,在迁飞至Aqaba海湾的北部海岸线时,为了避免飞入海里,蝗群折向北沿海岸飞行,直到飞到该海湾的最北端,蝗群又改变方向继续向东飞行。室内吊飞表明蝗虫不愿在光滑的反光的表面飞行,这减少了蝗群落入海水中的危险[38]。当然,也有蝗群越洋飞行的报告[39]。
目前涉及飞蝗聚集意义的研究相对较少。一般而言,聚集飞蝗通过群体信息传递机制,获得超出个体感官所能捕捉到的信息,如食物位置,增加了个体获得食物的机会。群居个体获得食物几率还与个体在群体中的位置和食物的丰盛度有关。模拟表明,当食物充足时,有利于处于群体边缘的个体取食;当食物稀缺时,处于群体中心的个体将处于优势地位[40]。另外,沙漠蝗转变成群居型后,会取食含有毒素的食物,再加上群居型深黑色的体色,从而获得对抗天敌的警示作用。最为直观的是群居飞蝗在交配时更为便利。复杂理论表明,大量个体可以自组织成有序体系,这个体系可以产生不同模式、储存信息、做出判断和决定,从而在动物漫长的自然选择压力中增加生存进化的几率[41]。可以设想蝗虫在进化中无需发展复杂的结构,而是通过这种简单有序叠加的行为获得散居蝗虫不能获得的生态效益,这对于种群进化具有重要意义。
[1] 朱恩林.中国东亚飞蝗发生与治理[M].北京:中国农业出版社, 1999.
[2] 黄辉,朱恩林.我国蝗虫发生防治动态[J].大自然, 2001(5):29.
[3] 施和金.论中国历史上的蝗灾及其社会影响[J].南京师大学报(社会科学版), 2002(2):148-154.
[4] 朱道弘, 阳柏苏.飞蝗变型及体色多型的内分泌控制机理[J].动物学研究, 2004, 25(5):460-464.
[5] Heifetz Y, Miloslavski I, Aizenshtat Z, et al.Cuticular surface hydrocarbons of desert locust nymphs,Schistocercagregaria, and their effect on phase behavior[J].Journal of Chemical Ecology, 1998,24(6): 1033-1047.
[6] 陈永林.飞蝗新亚种——西藏飞蝗Locustamigratoriatibetensissubsp.n.[J].昆虫学报, 1963,12(4):463-475.
[7] 赵春晓, 崔为正.飞蝗的型变及其调控机理[J].中国植保导刊, 2006, 26(9):11-13.
[8] Bruno H, Filip S, Tim V, et al.A miniaturized assay to quantify effects of chemicals or physical stimuli upon locust activity[J].Insect Science, 2006, 13(5):401-407.
[9] 郭志永, 石旺鹏, 张龙, 等.东亚飞蝗行为和形态型变的判定指标[J].应用生态学报, 2004, 15(5):859-862.
[10]Emma D.Locust plagues then and now[J].Bulletin of Entomological Society of Canada, 2005, 37(2):72-74.
[11]Emma D, Stephen J S.Food choices of solitarious and gregarious locusts reflect cryptic and aposematic antipredator strategies[J].Animal Behaviour, 2005, 69:471-479.
[12]Emma D, Stephen J S.Surviving the change to warning colouration: density-dependent polyphenism suggests a route for the evolution of aposematism[J].Chemoecology, 2005, 15:69-75.
[13]Faure J C.The phases of locusts in South Africa[J].Bulletin of Entomological Research, 1932,23:293-405.
[14]Koutaro Maeno, Seiji Tanaka.Effects of hatchling body colour and rearing density on body colouration in last-stadium nymphs of the desert locust,Schistocercagregaria[J].Physiological Entomology, 2007,32:87-94.
[15]张龙, 李洪海.虫口密度和龄期对东亚飞蝗群居型向散居型转变的影响[J].植保技术与推广, 2002,22(4):3-5.
[16]Mohamed Abdallahi Ould Babah, Gregory A S.Linking locust gregarization to local resource distribution patterns across a large spatial scale[J].Environmental Entomology, 2004,33(6):1577-1583.
[17]Matthew C, Emma D, Stephen J S, et al.Spatial scales of desert locust gregarization[J].Proceedings of the National Academy of Science of USA, 1998,95:13052-13055.
[18]Emma D, Simpson S J.Resource distribution mediates synchronization of physiological rhythms in locust groups[J].Proceedings of the Royal Society B, 2006,273:1517-1522.
[19]Emma D.Fractal index captures the role of vegetation clumping in locust swarming[J].Functional Ecology, 2003,17:315-322.
[20]Wopke V D W, Gebremedhin W, Arnold V H, et al.Plant communities can predict the distribution of solitarious desert locustSchistocercagregaria[J].Journal of Applied Ecology, 2005, 42:989-997.
[21]石旺鹏.化学信息素对东亚飞蝗聚集行为的调控作用[D].北京:中国农业大学, 2000.
[22]Obeng-Ofori D, Torto B, Njagi P G N, et al.Fecal volatiles as part of the aggregation pheromone complex of the desert locust,Schistocercagregaria(Forskal) (Orthoptera:Acrididae)[J].Journal of Chemical Ecology, 1994, 20:2077-2087.
[23]Torto B, Njagi P G N, Hassanali A, et al.Aggregation pheromone system of nymphal gregarious desert locust,Schistocercagregaria(Forskal)(Orthoptera:Acrididae)[J].Journal of Chemical Ecology, 1996, 22:2273-2281.
[24]石旺鹏.蝗虫化学信息物质研究进展[J].昆虫知识, 2005,42(3):244-249.
[25]Ahmed H, Peter G N N, Magzoub O B.Chemical ecology of locusts and related acridids[J].Annual Review of Entomology, 2004,50: 223-245.
[26]Leo R, Lester C G, Meir P P, et al.Stimuli inducing gregarious colouration and behaviour in nymphs ofSchistocercagregaria[J].Journal of Insect Physiology, 2005,51: 737-747.
[27]Stephen M R, Thomas M, Emma D, et al.Mechanosensory-induced behavioural gregarization in the desert locust[J].The Journal of Experimental Biology, 2003,206:3991-4002.
[28]Sumpter D J T.The principles of collective animal behaviour[J].Philosophical Transactions of the Royal Society of London Series B-biological Sciences, 2006, 361: 5-22.
[29]Iain D C, Jens K, Richard J, et al.Collective memory and spatial sorting in animal groups[J].Journal of Theoretical Biology, 2002, 218:1-11.
[30]Charlotte K H, Hanspeter K.Density distribution and size sorting in fish schools: an individual-based model[J].Behavioral Ecology, 2004,16:178-187.
[31]Buhl J, Sumpter D J T, Couzin I D, et al.From disorder to order in marching locusts[J].Science, 2006, 312:1402-1406.
[32]Spork P, Preiss R.Adjustment of flight speed of gregarious desert locust (Schistocercagregaria) flying side by side[J].Journal of Insect Behavior, 1994,7(2):217-227.
[33]Stephan G R.Can a minority of informed leaders determine the foraging movements of a fish shoal[J].Animal Behaviour, 2000,59: 403-409.
[34]Jean-Marc A, Jose H, Colette R, et al.Collegial decision making based on social amplification leads to optimal group formation[J].Proceedings of the National Academy of Science of USA, 2006, 103(15):5835-5840.
[35]Iain D C, Jens K, Nigel R F, et al.Effective leadership and decision-making in animal groups on the move[J].Nature, 2005,433:513-516.
[36]Conradt L, Roper T J.Group decision-making in animals[J].Nature, 2003, 421:155-158.
[37]Heitler W J, Mitchell J L, Dinwiddie L.Underwater locomotion in the desert locust: behavioural choice when confronted with an aquatic barrier[J].Journal of Insect Behavior, 2005, 18(5):669-683.
[38]Shashar N, Sabbah S, Aharoni N.Migrating locusts can detect polarized reflections to avoid flying over the sea[J].Biology Letters, 2005,1:472-475.
[39]Jane R, Peter J A B.Windborne displacements of desert locusts from Africa to the Caribbean and South America[J].Aerobiologia, 1999, 15(3): 167-175.
[40]Hirsch B T.Costs and benefits of within-group spatial position: a feeding competition model[J].The Quarterly Review of Biology, 2007,82:9-27.
[41]Julia K P, Leah E K.Complexity, pattern, and evolutionary trade-offs in animal aggregation[J].Science, 1999, 284:99-101.
Researchprogressonaggregationbehaviorsoflocusts
Feng Chuanhong1,Chen Shuqun2,Guo Cong3,Zhang Yuejin4,Liu Wancai4,Luo Linming1
(1.PlantProtectionStationofSichuan,Chengdu610041,China; 2.PlantProtectionStationofRenshou,Sichuan620500,China; 3.CollegeofLifeScience,SichuanUniversity,Chengdu610064,China; 4.NationalAgriculturalTechnologyExtensionandServiceCenter,Beijing100125,China)
Locusts show aggregation behaviors in the fields.Solitary and gregarious locusts differ in many traits, such as morphology,physiological and biochemical characteristics and behaviors.The aggregation behavior is closely correlated with the density of locusts, distribution patterns of resources, pheromone and special action of locusts.After locusts crowed, movement of locust swarm in migration follows a complex system based on simple laws.Individuals within the group highly align collective motion.The highly ordered mechanism was beneficial to extend useful information in locust group so that the whole population could approach the largest ecological benefits.
locusts; aggregation behavior; ordered; ecological benefits
2013-03-25
: 2013-06-07
公益性行业(农业)科研专项(200903004)
S 433.2
: ADOI: 10.3969/j.issn.0529-1542.2014.01.003
联系方式 E-mail:fengchuanhong8@163.com
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
学苑创造·A版(2022年3期)2022-03-29
民间故事选刊·上(2022年1期)2022-02-09
幼儿智力世界(2021年8期)2021-12-17
故事作文·高年级(2021年11期)2021-11-28
小小说月刊·下半月(2020年9期)2020-10-09
科学大众(2020年10期)2020-07-24
植物保护(2020年2期)2020-06-08
故事作文·低年级(2019年6期)2019-06-20
幼儿教育·父母孩子版(2017年10期)2017-12-13
