
毛竹碳汇林营造初期林分非空间结构年际变化特征
2015-03-23 08:56施拥军周国模郑泽睿羽中
浙江农林大学学报 2015年2期
陈 婷,施拥军,2,周国模,2,郑泽睿,李 羽中
毛竹碳汇林营造初期林分非空间结构年际变化特征
陈 婷1,施拥军1,2,周国模1,2,郑泽睿1,李 羽中1
(1.浙江农林大学 浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安 311300;2.浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安311300)
在新造毛竹Phyllostachys edulis碳汇林内,设置80 m×100 m固定标准地,采用固定样地长期连续观测的调查方法,研究新造毛竹 碳汇林在成林前(第1~5年)林分非空间结构动态变化规律及影响因子。结果表明:①新造毛竹碳汇林成林初期,立竹度、林分平均胸径及林分平均竹高是林分结构变化的主导因子,且随着造林后年限的增加,立竹度、林分平均胸径及林分平均竹高均在不断增加。②新造毛竹碳汇林在成林过程中,立竹度(N)随造林后年限(y)的增加而增加,并满足异速生长方程:N=17.214y2+309.21y-217.4,R2=0.981 1。③随着造林后年限的增加,新出毛竹的平均胸径逐年增大,新竹平均胸径(DBH)与造林后年限(y)的异速生长方程为:DBH=0.015 7y2+0.727 9y+1.598 0,R2=0.756 5。④每年新出毛竹的平均竹高(H)随着造林后年限(y)的增加而增高,其拟合方程为:H=0.108 5y2+0.429 9y+2.155 3,R2=0.723 2。图4表8参20
森林测计学;新造毛竹林;碳汇林;立竹度;胸径;竹高;异速生长方程
在全球共同应对气候变化,大力发展低碳经济的背景下,森林生态系统具有强大的固碳减排功能,是陆地上最大的碳储存库,在低碳经济中有着不可替代的作用,森林碳汇成为相当重要的研究领域。碳汇造林也成为增加碳储量的一种重要的方式,被中国乃是世界各国广泛采用。竹林是重要的森林植被类型,也是特殊的生态系统单元。竹林生态系统的碳储量及固碳能力不可忽视,成为除乔木林以外最大的碳汇,且竹林拥有稳定的碳汇功能[1-3]。许多学者研究表明:毛竹Phyllostachys edulis具有很强的固碳能力,是速生阶段杉木的1.46倍,是热带山地雨林的1.33倍,是苏南27年生杉木林的2.16倍[4]。同时毛竹又具有良好的经济利用价值,深受山区农民喜爱,因此,毛竹造林成为中国碳汇造林的一种重要方式。碳汇造林是指在确定了基线的土地上,以增加碳汇为主要目的,对造林及林木(分)生长过程实施碳汇计量和监测而开展的有特殊要求的造林活动。与普通的造林相比,碳汇造林突出森林的碳汇功能,强调森林的多重效益,需要对碳储量的变化过程进行科学的计量与监测工作,以获得可测量、可报告、可核查的碳汇量,以满足后续碳汇交易的要求。新造毛竹林生长特殊,分为成林前和成林后2个阶段,成林之前的竹林林分生长较为复杂,随着造林后年限的增长,毛竹林林分的立竹度、平均胸径、平均竹高也会不断发生变化,这与成林后较为稳定的林分结构(立竹度、胸径、竹高)有着本质的区别。虽然,对毛竹林成林后的林分结构已经有了较多的研究[5-18],但是对于毛竹林成林前的研究几乎为空白。竹林成林前的林分结构和碳储量变化是当前竹林碳汇计量中面临的最大困难。为此,本研究拟通过碳汇造林试验和长期的监测工作,以探究毛竹成林前的林分结构动态变化特征,建立新造毛竹林成林前的异速生长模型,为毛竹林的碳汇计量监测提供支持。
1 研究区域概况
浙江农林大学2008年在浙江省临安市藻溪镇严家山,通过母竹移栽方式新造了1块毛竹碳汇林,面积为47.72 hm2。研究试验区选在新造的毛竹碳汇林内,中心地理坐标为30°10′20″N,119°29′40″E,平均海拔为550 m。该区属中亚热带季风气候,年平均气温为15.8℃,年降水量为1 500 mm左右,无霜期为236 d,地形地貌为低山丘陵,土类为红壤,亚类为黄红壤,土属为黄泥土,土种为黄泥砂土。成土母质主要为砂页岩类残坡积物和砂砾类残坡积物,土壤呈酸性。土壤pH 4.4,土壤有机质质量分数为36 g·kg-1,碱解氮254.3 mg·kg-1,速效磷126.3 mg·kg-1,有效磷为17.6 mg·kg-1。
毛竹碳汇林初植密度为450株·hm-2,在毛竹林发育成林阶段,竹林抚育措施为春季和初夏各中耕除草1次·a-1,造林后3 a内采用穴施方式施用尿素300~450 kg·hm-2·a-1,除挖取退笋、败笋外,全部留笋长竹,不进行伐竹作业。
2 野外调查方法
2.1 样地设置
2008年,在浙江省临安市藻溪镇严家山新造的毛竹碳汇林地内,选择典型的毛竹林设置80 m× 100 m固定标准地,固定标准地海拔为450~500 m。用高精度全站仪(NTS-352,测角精度2″,测距精度3 mm+2 mm/km)把固定标准地划分为完全等面积的20个20 m×20 m的样地(调查单元),使样地构成相邻格网。
2.2 坐标测定
采用全站仪(NTS-352)精确测定每个角桩的三维坐标(x,y,z)及每株毛竹基部三维坐标,其中x和y是平面直角坐标,z是海拔高度。
2.3 毛竹林林分因子调查
在每个调查单元内,对胸径DBH≥1 cm的毛竹进行每木调查。从2008年开始,每隔2 a测量新竹基部三维坐标,并进行每木检尺,测量并记录竹编号、竹龄、竹高、胸径等因子。对样地内新出毛竹进行连续不间断监测。胸径的测量采用围尺,以距地面1.3 m处胸高位置为测定标准,胸高处出现节疤,凹凸或其他不正常情况时,可在胸高上下距离相等而形状较正常处,测2个直径取平均数作为胸径;竹高的测量采用全站仪悬高测量法(测定空中某点距地面的垂直高度)。到2013年(为期5 a)共测定毛竹1 809株及母竹330株。由于母竹在造林后几年内被清除,所以本研究所述的立竹度为不包括母竹的新竹立竹度。
3 结果与分析
3.1 新造毛竹碳汇林林分结构年际变化特征
竹龄、立竹度、胸径和竹高等是林分结构变化的重要指标。新造毛竹林主要生长因子动态变化见表1。从表1可以看出:在研究样地内,新造毛竹林造林后第1年成竹数为48株,到造林后第5年立竹度达到1 809株,呈急速上升趋势;新造毛竹林内毛竹林林分平均胸径从第1年的2.6 cm增加到第5年的4.4 cm;新造毛竹林林分平均竹高也发生着明显的变化,造林后第1年林分平均竹高3.1 m,到造林后第5年林分平均竹高达到5.3 m。
综上可知:新造毛竹林在整个成林过程中,随着造林后年限的增加,立竹度在不断增加,林分平均胸径也在增大,林分平均竹高在不断增高。可以看出:新造毛竹林是一种随着造林年限不断动态变化着的,立竹度、林分平均胸径以及林分平均竹高与造林后年限之间存在异速生长模型。

表1 新造毛竹碳汇林林分特征Table 1 The characteristics of moso bamboo samples
3.2 新造毛竹林各年龄非空间结构指标的方差分析
为探究新造毛竹林非空间结构的各指标年际变化的影响及其显著性水平,采用SPSS 19.0软件,对造林后不同年限的毛竹的立竹度、新竹胸径、新竹竹高等做单因素方差分析,结果见表2。可以看出:新造毛竹林中不同造林后年限(竹龄)新竹的立竹度、胸径、竹高等因子均呈极显著差异(P<0.01),说明造林后年限的变化是新造毛竹林成林前的林分结构变化的重要因子。
为探究大样地内部是否存在差异性,对20个子样地做方差分析,结果见表3。表3表明:样地之间立竹度具有显著差异(P=0.000),但不管是对胸径还是对竹高而言,样地之间的P都远大于0.05,说明样地之间胸径和竹高没有显著差异;

表2 新造毛竹碳汇林不同年龄非空间结构各指标变化的显著性检验Table 2 Each index’s significant test of non spatial structurewith different ages

表3 新造毛竹碳汇林各子样地间非空间结构各指标变化的显著性检验Table3 Each index’s significant test of non spatial structurewith different sub-plots
3.3 新造毛竹碳汇林不同造林后年限毛竹立竹度变化
立竹度是竹林出笋前单位面积内正常生长的竹子株数,是新造毛竹林林分结构变化的重要指标,在毛竹林成林前期,会随着造林年限呈急速增加趋势。表4为新造毛竹林新竹株数的年际变化。可见,在研究样地内,新造毛竹林造林后第1年新竹的成竹数48株,造林后第2年新竹的成竹数为555株,为出笋成竹盛期,第3年和第4年新竹分别成竹229株和374株,第5年新竹出笋成竹数为603株。第1年新竹出笋成竹数较少,第2年和第5年新竹出笋成竹数较高,到达出笋成竹盛期。但林分内,随着造林年限的增加,立竹度随造林后年限的增加呈线性上升趋势(表1)。
采用Pearson相关性分析方法对新造毛竹林立竹度与造林后年限之间做相关性分析,呈显著相关性(R=0.991)。对不同造林后年限和立竹度进行方差分析(表2),F分布的显著性概率为0.000,达到了极显著水平,有统计学意义。这说明因变量和自变量的关系极显著。在成林过程中,立竹度(N)随造林后年限(y)变化的异速生长方程为:N=17.214y2+309.210y-217.400(R2=0.981 1)。据此模型,可以通过了解新造毛竹林的造林后年限计算出新造毛竹林各林分的立竹度。

表4 新造毛竹碳汇林新竹株数年际变化Table 4 Changes of bamboo stand culms with year
3.4 新造毛竹碳汇林各年龄毛竹胸径变化
新造毛竹林在成林过程中,随着造林后年限的增加,除了立竹度增加以外,新竹平均胸径也会逐步增大,与成林后的稳定竹林表现出完全不一致的特征[19]。本研究表明(表5):新造毛竹林在造林后第1年新竹平均胸径为2.6 cm,到成林后的第5年新竹平均胸径增加到5.8 cm,说明随着年龄的增大新造毛竹林新竹平均胸径在快速的增加,各年份最小胸径差异性不大,为1.0~1.4 cm;胸径的极大值变化较大,变化幅度为5.4~10.2 cm。随着造林年限的增加,毛竹林个体之间的差异也在不断变大,标准差从第1年的0.88 cm增加到第5年的1.56 cm。
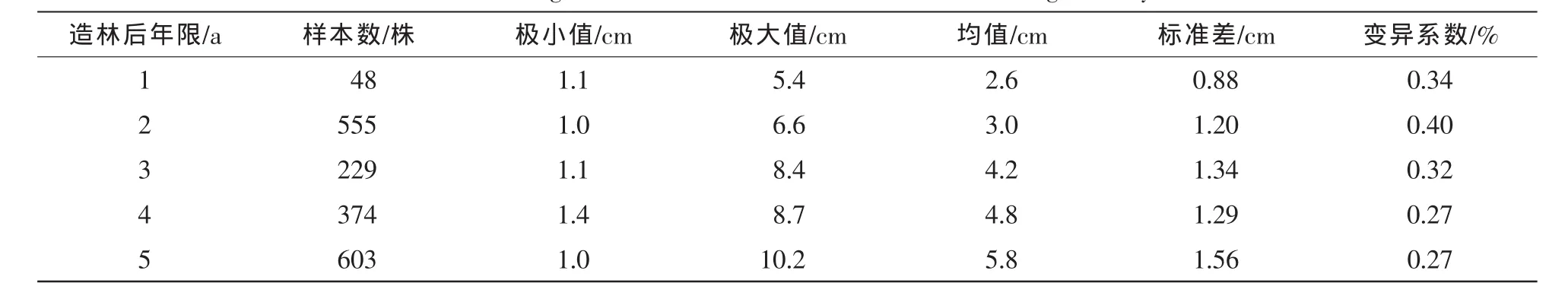
表5 新造毛竹碳汇林新竹胸径年际变化Table 5 Change of new bamboo stand diameter in breast height with year
采用Pearson相关性分析方法对新造毛竹林新竹胸径与造林后年限之间做相关性分析,呈显著相关性(R=0.992)。对新造毛竹林各子样地内不同年龄新竹胸径求平均,得到散点图(图1),并进行曲线拟合,得到不同新造毛竹林新竹胸径年际变化异速生长模型(表6),结果表明:新造毛竹林在成林过程中,新竹的平均胸径逐年增大。通过模型比较,多项式模型DBH=0.015 7y2+0.727 9y+1.598 0,R2=0.756 5,能够最好地反映新造毛竹林新竹平均胸径随年龄的递增状况。其次是线性模型D=0.824 8y+1.477 9,R2= 0.756 1。
从图2可以看出:新造毛竹林立竹度按径阶的分布符合威布尔分布,株数最多的集中在1.5~6.5 cm,随着造林后年限的增加,新竹的径阶分布将向大径阶移动,未来将逐渐趋向于正态分布。
同时,对研究样地内对不同造林年限的毛竹林立竹度和林分平均胸径进行相关性分析,发现新造毛竹林林分平均胸径和立竹度(林分密度)之间呈正相关(R2=0.974 0,P=0.005),对新造毛竹林内各年份林分平均胸径和立竹度进行曲线拟合,得到线性方程: DBH=0.000 9N+2.366 9,R2=0.950 0,函数曲线见图3。此模型说明:新造毛竹林在成林初期,随着造林后年限的增加,新竹平均胸径在年际增大的同时,立竹度也在不断增加。竹林立竹度的大小,与竹林新竹产量有密切关系。由此看来,应该是立竹度影响翌年的新竹大小。相对于毛竹胸径的测量,立竹度数据更便于获得。
3.5 新造毛竹碳汇林各年龄毛竹竹高变化
新造毛竹林在成林过程中,随着造林后年限的增加,除了立竹度和新竹平均胸径在不断增加,新竹平均竹高也会逐渐变高,与成林后的稳定竹林表现出完全不一致的特征[19]。本研究表明(表7),新造毛竹林在造林后第1年平均新竹竹高为3.1 m,到造林后的第5年新竹平均竹高迅速增加到7.3 m,说明随着造林后年限的增加,新造毛竹林新竹平均竹高也在快速的高,各年份新竹竹高最小值的差异性不大,为1.0~1.9 m;新竹竹高的变化主要体现在极大值上,变化幅度为6.3~18.0 m。随着造林年限的增加,毛竹林个体之间的差异也在不断变大,标准差从第1年的1.01 m增加到第5年的2.06 m。

表6 新造毛竹碳汇林新竹胸径年际变化异速生长模型比较Table 6 Different types of allometric models for predicting new moso bamboo by DBHand age

图1 新造毛竹碳汇林新竹平均胸径年际变化Figure 1 Change of bamboo stand diameterin bright with year

图2 新造毛竹碳汇林径阶分布Figure 2 Diameter distribution of mosobamboo

图3 不同年限立竹度和林分平均胸径之间的关系Figure 3 Relationships between mean DBHand culms of new moso bamboo

表7 新造毛竹碳汇林新竹竹高年际变化Table 7 Changes of the height of new moso bamboo stand with year
采用Pearson相关性分析方法对新造毛竹林新竹竹高和造林后年限之间做相关性分析,结果表明:新造毛竹林新竹竹高与造林后年限呈显著相关(R2=0.884 0)。对20个子样地中的不同造林后年限的新竹竹高分别取平均,生成散点图(图4),并进行曲线拟合,得到不同新造毛竹林新竹平均竹高年际变化异速生长模型(表8)。结果表明:新造毛竹林在成林过程中,新竹平均竹高和造林后年限呈正相关,随着新造毛竹林造林后年限的增加,新竹的平均竹高也逐年增大。通过模型比较发现,多项式模型:H= 0.108 5y2+0.429 9y+2.155 3,R2=0.723 2能够最好地反映新造毛竹林新竹平均竹高随不同造林后年限的递增状况;其次是线性模型H=1.099 8y+1.324 4,R2=0.713 5。
4 结论与讨论

图4 新造毛竹碳汇林子样地新竹平均竹高年际变化Figure 4 Changes of bamboo stand culm height with year in sub-plots
本研究在新造毛竹碳汇林地内设置典型固定大样地,并连续监测,共获得5 a新造毛竹碳汇林林分数据,对毛竹林非空间结构特征及动态变化进行研究,并对数据建模。得到如下结论。①新造毛竹碳汇林立竹度、造林后年限和林分平均胸径是新造毛竹碳汇林林分非空间结构变化的主导因子,林分平均竹高是新造毛竹碳汇林林分非空间结构变化的从属因子;②新造毛竹碳汇林在成林过程中,立竹度(N)随造林后年限(y)增加呈线性上升,并满足异速生长方程:N=17.214 0y2+309.210 0y-217.400 0,R2=0.981 1。③新造毛竹碳汇林在成林过程中,随着造林年限的增加,新竹的平均胸径逐年增大,新竹平均胸径(DBH)与造林后年限(y)满足异速生长方程为:DBH=0.015 7y2+0.727 9y+1.598 0,R2=0.756 5。④新造毛竹碳汇林在成林过程中,新竹平均竹高(H)随着造林后年限(y)增加逐渐增高,其拟合方程为:H=0.108 5y2+0.429 9y +2.155 3,R2=0.723 2。
本研究发现新造毛竹碳汇林成林初期林分平均胸径和立竹度呈正比,其线性关系为: DBH=0.000 9N+2.366 9,R2=0.950 0,而毛竹碳汇林成林之后立竹度与平均胸径呈反比[20],说明毛竹碳汇林成林前的林分结构变动特征与成林后的林分结构特征不一致,其可能的原因是:毛竹林成林前竞争较弱,新竹平均胸径每年不断增加,并且由于竹鞭的蔓延,大量发笋,导致立竹度也不断增加;成林后由于空间和土壤养分的限制,立竹度的大量增加会导致林分内的竞争渐趋激烈,从而影响发笋的数量和质量,因此随着立竹度的增加平均胸径会减小。
陈双林等[19]年对新造毛竹林林分结构演替规律的研究中提出过新造毛竹林的胸径异速生长方程,但采用的调查方法是在同一时间内对不同竹龄的进行调查研究,可能存在以时间换空间的概念,对竹子的年龄的判断具有不确定性,而且样地的初植密度不一,在建模时很容易产生误差。而本研究是基于相同初植密度的20块标准子样地每年连续调查的基础上进行建模,数据更加可靠。
本研究是基于新造毛竹碳汇林成林初期(第1~5年)所进行的研究,毛竹种植后至成林前,采用一般经营模式,通过不断发笋、留笋和养竹第7年后可以成林,一般毛竹到达成林稳定阶段往往需要6~7 a[19]或更长时间。本研究今后还将持续监测成林扩展特征,补充完善相关的模型。

表8 新造毛竹碳汇林新竹竹高年际变化异速生长模型比较Table 8 Different types of allometric models for predicting moso bamboo by culm height and age
[1] 郭起荣,杨光耀,杜天真,等.中国竹林的碳素特征[J].世界竹藤通讯,2005,3(3):25-28. GUO Qirong,YANG Gangyao,DU Tianzhen,et al.Carbon character of Chinese bamboo forest[J].World Bamboo Rattan,2005,3(3):25-28.
[2] 肖复明,范少辉,汪思龙,等.毛竹Phyllostachy pubescens杉木Cunninghamia lanceolata人工林生态系统碳储量及其分配特征[J].生态学报,2007,27(7):2794-2801. XIAO Fuming,FAN Shaohui,WANG Silong,et al.Carbon storage and spatial distribution in Phyllostachy pubescens and Cunninghamia lanceolata plantation ecosystem[J].Acta Ecol Sin,2007,27(7):2794-2801.
[3] 陈先刚,张一平,张小全,等.过去50年中国竹林碳储量变化[J].生态学报,2008,28(11):5218-5226.CHEN Xiangang,ZHANG Yiping,ZHANG Xiaoquan,et al.Carbon stock changes in bamboo stands in China over the last 50 years[J].Acta Ecol Sin,2008,28(11):5218-5226.
[4] 周国模,姜培坤.毛竹林的碳密度和碳贮量及空间分布[J].林业科学,2004,40(6):20-24. ZHOU Guomo,JIANG Peikun.Density,storage and spatial distribution of carbon in Phyllostachy pubescens forest [J].Sci Silv Sin,2004,40(6):20-24.
[5] 聂道平.江西省大岗山毛竹林的结构特征[J].林业科学研究,1992,5(6):693-699. NIE Daoping.The structure characteristics of baoboo groves in Dagangshan Region,Jiangxi Province[J].For Res, 1992,5(6):693-699.
[6] 聂道平.毛竹林结构的动态特性[J].林业科学,1994,30(3):201-208.NIE Daoping.Structural dynamics of bamboo forest stands[J].Sci Silv Sin,1994,30(3):201-207.
[7] 何东进,洪伟,吴承祯.毛竹林林分平均胸径模拟预测模型的研究[J].林业科学,2000,36(1):148-153. HE Dongjin,HONG Wei,WU Chengzhen.A study on simulating predictive model of mean DBH for bamboo stands[J].Sci Silv Sin,2000,36(1):148-153.
[8] 朱锦懋,黄茂提,陈由强,等.笋材两用毛竹林林分结构数量关系研究[J].植物生态学报,2000,24(4):483 -488. ZHU Jinmao,HUANG Maoti,CHEN Youqiang,at al.The structure of a culm and shoot producing stand of Phyllostachys pubescens[J].Acta Phytoecol Sin,2000,24(4):483-488.
[9] 汤孟平,徐文兵,陈永刚,等.地形条件对毛竹林分结构和植被碳储量的影响[J].林业科学,2011,47(8):1 -6. TANG Mengping,XU Wenbing,CHEN Yonggang,et al.Relationship between spatial structure and biomass of a close-to nature Phyllostachys edulis stand in Tianmu Mountain[J].Sci Silv Sin,2011,47(8):1-6.
[10] 邓英英,汤孟平,徐文兵,等.天目山近自然毛竹纯林的竹秆空间结构特征[J].浙江农林大学学报,2011,28(2):173-179. DENG Yingying,TANG Mengping,XU Wenbing,et al.Spatial structure of bamboo culm of an almost natural,pure Phyllostachys pubescens forest in Mount Tianmu[J].J Zhejiang A&F Univ,2011,28(2):173-179.
[11] 白尚斌,周国模,王懿祥,等.天目山国家级自然保护区毛竹扩散过程的林分结构变化研究[J].西部林业科学,2012,41(1):77-82. BAI Shangbin,ZHOU Guomo,WANG Yixiang,et al.Stand structure change of Phyllostachys pubescens forest expansion in Tianmushan National Nature Reserve[J].J West China For Sci,2012,41(1):77-82.
[12] 刘恩斌,施拥军,李永夫,等.浙江毛竹林分非空间结构特征及其动态变化[J].林业科学,2013,49(9):1-7. LIU Enbin,SHI Yongjun,LI Yongfu,et al.Non spatial structural characteristic of moso bamboo forest and its dynamics in Zhejiang Province[J].Sci Silv Sin,2013,49(9):1-7.
[13] 徐振国,郭起荣,冯云,等.淡竹林分结构与林分因子相关研究[J].中南林业科技大学学报,2014,34(2):72-76. XU Zhenguo,GUO Qirong,FENG Yun,et al.Related research on structure characteristics and stand factors of Phyllostachys glauca[J].J Cent South Univ For&Technol,2014,34(2):72-76.
[14] 范叶青,周国模,施拥军,等.地形条件对毛竹林分结构和植被碳储量的影响[J].林业科学,2013,49(11):178 -182. FAN Yeqing,ZHOU Guomo,SHI Yongjun,et al.Effects of terrain on stand structure and vegetation carbon storage of Phyllostachys edulis forest[J].Sci Silv Sin,2013,49(11):178-182.
[15] ARUN J N,GITASREE D,ASHESH K D.Above ground standing biomass and carbon storage in village bamboos in North East India[J].Biom Bioenergy,2009,33(9):1188-1196.
[16] DU Huaqiang,CUI Ruirui,ZHOU Guomo,et al.The responses of moso bamboo (Phyllostachys heterocycla var. pubescens)forest aboveground biomass to Landsat TM spectral reflectance and NDVI[J].Acta Ecol Sin,2010,30 (5):257-263.
[17] YEN T M,JI Y J,LEE J S.Estimating biomass production and carbon storage for a fast-growing makino bamboo (Phyllostachys makinoi)plant based on the diameter distribution model[J].For Ecol Manage,2010,260(3):339 -344.
[18] FU Weijun,JIANG Peikun,ZHAO Keli,et al.The carbon storage in moso bamboo plantation and its spatial variation in Anji County of southeastern China[J].J Soils Sediment,2014,14(2):320-329.
[19] 陈双林,吴柏林,吴明,等.新造毛竹林林分结构年际演替规律及影响因子[J].浙江林学院学报,2004,21(4): 393-397. CHEN Shuanglin,WU Bolin,WU Ming,et al.A study of the interannual succession rule and influential factors of young stands structures of Phyllostachys pubescens[J].J Zhejiang For Coll,2004,21(4):393-397.
[20] YEN T M,LEE J S.Comparing aboveground carbon sequestration between moso bamboo(Phyllostachys heterocycla)and China fir(Cunninghamia lanceolata)forests based on the allometric model[J].For Ecol Manage,2011,261(6): 995-1002.
Interannual variation characteristics for stand structures in the early stages of new moso bamboo carbon sink stands
CHEN Ting1,SHI Yongjun1,2,ZHOU Guomo1,2,ZHENG Zerui1,LI Chong1
(1.Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration,Zhejiang A &F University,Lin’an 311300,Zhejiang,China;2.The Nurturing Station for the State Key Laboratory of Subtropical Silviculture,Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
To determine a rule for interannual variation characteristics in non-spatial,structural stands of new moso bamboo (Phyllostachys edulis)based on carbon sink (1-5 a).An 80 m×100 m fixed plot was established at a long-term,afforestation,fixed observation research site for carbon sequestration.Linear regression and correlation were used for the analysis.Results showed that 1)stand density,DBH,and culm height were the dominating factors for interannual component changes,and these increased with bamboo age.2)In the young stand,stand density(N)increased with stand year(y)with a fitted regression equation of:N=17.214y2+309.21y-217.4,R2=0.981 1.3)The allometric equation for mean DBH and stand year(y)fitted with linear regression was DBH=0.015 7y2+0.727 9y+1.598 0,R2=0.756 5.4)The linear regression for the relationship between culm height(H)and stand year(y)was projected to be H=0.108 5y2+0.429 9y+2.155 3, R2=0.723 2.Therefore,in the young moso bamboo stand,stand density,DBH,and culm height all had a positive relationship with age.[Ch,4 fig.8 tab.20 ref.]
forest mensuration;new moso bamboo forest;carbon sequestration forest;stand density;DBH;culmheight;allometric model
S758
A
2095-0756(2015)02-0181-07
浙 江 农 林 大 学 学 报,2015,32(2):181-187
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.02.003
2014-07-26;
2014-08-28
“十二五”国家科技支撑计划项目(2012BAD22B0503);国家林业局引进国际先进农业科学技术计划(“948”计划)项目(2013-4-71);国家自然科学基金资助项目(31370637);浙江省重点科技创新团队项目(2010R50030);浙江省自然科学基金资助项目(Y5110145)
陈婷,从事森林碳汇计量与监测研究。E-mail:ctzun_1990@126.com。通信作者:施拥军,副教授,从事森林碳汇计量与监测研究。E-mail:syjwwh@163.com
猜你喜欢
科教新报(2022年19期)2022-06-13
科教新报(2022年17期)2022-05-24
保健与生活(2022年10期)2022-05-06
天津农林科技(2022年2期)2022-04-19
浙江农林大学学报(2020年5期)2020-11-05
保健与生活(2020年10期)2020-05-28
湖州职业技术学院学报(2020年4期)2020-03-30
阅读(高年级)(2018年6期)2018-05-14
阅读(高年级)(2018年1期)2018-05-14
小天使·一年级语数英综合(2015年8期)2015-07-06
