
帽儿山林区紫椴群落物种多样性、种间关系及对环境因子的响应
2015-12-16 08:11张东来
森林工程 2015年6期
张东来,张 玲
(1.黑龙江省林业科学院,哈尔滨150081;2.黑龙江省林业科学研究所,哈尔滨150081)
紫椴(Tilia amurensis Rupr.)不仅是我国东北地区重要的珍贵阔叶树种,而且是重要的蜜源、城市绿化以及药用树种[1-2]。因为连年过量开采,致使天然紫椴种群无论从数量上还是从质量上都呈逐年下降与衰退的趋势,面积和蓄积量锐减,现在已被列为国家Ⅱ级保护植物。
紫椴天然更新良好,但研究发现紫椴人工培育困难,主要是受霜冻害的制约,到目前为止,紫椴人工培育过程中幼树易受霜冻危害的问题始终未得到有效解决,霜冻危害成为制约紫椴人工培育的瓶颈。加强紫椴人工林资源的培育研究,尤其是紫椴混交林的研究已迫在眉睫,这将对保护和扩大紫椴资源,使其永续利用具有深远历史意义和更重要的现实意义[3]。
本研究以帽儿山林区紫椴天然种群为研究对象,开展群落的物种多样性、种间关系及其与环境关系的研究,掌握紫椴群落的组成、结构和功能与环境关系,为今后紫椴人工林培育提供技术支撑。
1 研究区概况
研究地点位于黑龙江省尚志市东北林业大学帽儿山实验林场内,该林场位于北纬45°21'~45°25',东经 127°31'~127°34',林场平均海拔高度 300 m,最高海拔为805 m,属长白山系张广才岭西部小岭余脉,植被属长白植物区系,是东北东部山区较典型的天然次生林区,本区土壤多为典型暗棕色森林土,有白浆化暗棕壤、草甸暗棕壤两个亚类,土壤下层为岩石。地带性顶极植被为阔叶红松林,平均林龄50 a。主要阔叶树种有紫椴(Tilia amurensis Rupr.)、水曲柳(Fraxinus mandshurica Rupr.)、暴马丁香(Syringa amurensis Rupr.)春榆(Ulmus japonica(Rehd.)Sarg.)、忍冬(Lonicera naackii(Rupr.)Maxim)、色木(Acer mono Maxim)、卫矛(Euonymus alatus(Thunb.)Sieb.)等 13 种植物。本区属大陆性气候,年降雨量723.8 mm,无霜期120~140 d[4]。
2 研究方法
2.1 野外调查
采用典型样地调查方法,2014年6月末,在帽儿山林场紫椴天然次生林内,沿海拔高度,在研究区域内设10个调查样方,规格为20 m×20 m,并在每个乔木样方内随机设5个灌木样方,规格为5 m×5 m,每个灌木样方内随机设5个草本样方,规格为1 m×1 m。对样方内的植物进行常规调查和记录,并在每个样地的4个角和中间取等量表土(0~20 cm)混合均匀,去除杂物,带回、风干,待测[4-5]。
2.2 数据处理
多样性指数的计算依据为重要值。以各层多样性指数和为计算整个群落多样性指数的依据,每种群落的多样性指数以平均值为准[5]。
乔木重要值IV=(相对多度+相对频度+相对显著度)/3。
灌木和草本植物重要值IV=(相对高度+相对盖度)/2。
群落多样性的测度选用丰富度指数(S)、均匀度指数和物种多样性指数。其计算公式如下:
物种丰富度D=S。
Shannon-Wiener index H'= ∑PilnPi。
Pielou指数:Jsw=(∑PilnPi)/lnS。
式中:Pi为种i的相对重要值;S为中i所在样方的物种数,即丰富度指数。
2.3 成对物种间关联的测度
Ochiai指数 OI=a/ [(a+b)(a+c)]1/2。
Dice指数 DI=2a/(2a+b+c)。
Jaccard指数 JI=a/(a+b+c)。
式中:a为2个种都存在的取样数量;b为B种存在而A种不存在的取样数量;c为A种存在而B种不存在的取样数目;d为2个种都不存在取样多数。
以上3个无中心指数的值域为0到1。其值越趋近于1,则表明该种对的正联结越紧密[1,6]。
2.4 土壤样品分析
用风干法测量土壤含水量,pH值用S-3C型酸度计测定,全氮含量用半微量凯氏定氮法测定;有机质采用沙浴外加热法-重铬酸钾氧化容量法,全磷含量用硫酸高氯酸消煮-钼锑抗比色测定,全钾含量采用氢氟酸-高氯酸消煮-原子吸收测定。
3 研究结果
3.1 帽儿山林区紫椴群落多样性指数与均匀度
帽儿山林区紫椴群落草本层物种多样性丰富,达到19.01±1.29,其次是灌木层,见表1。帽儿山林区紫椴群落乔木层物种丰富度较低,乔木种类少,主要伴生种为山桃稠李(Podus maackiii)、水曲柳、五角槭、黄榆、蒙古栎,胸径平均值为22.2 cm,郁闭度为0.7,还有一些空间没被占据,分布格局相对不均匀。随着恢复时间逐渐增长,群落结构逐渐呈现出复杂化,均匀度指数也越来越大,反映出该群落越来越稳定,达到顶级群落。灌木层主要伴生种为暴马丁香、金银忍冬、五角槭、卫矛(Euonymus alatus)、毛榛子(Corylus heterophylla)、溲疏(Deutzia parviflora)。草本层以铃兰(Convallaria keiskei)、蚊子草(Filipendula palmate)、四花苔草(Carex quadriflora)、委陵菜(Potentilla chinensis)、玉竹(Polygonatum odoratum)、木赋(Equisetum hiemale)菊科、莎草科、毛茛科、蔷薇科为主。香浓指数则是乔木层和草本层均显著高于灌木层。
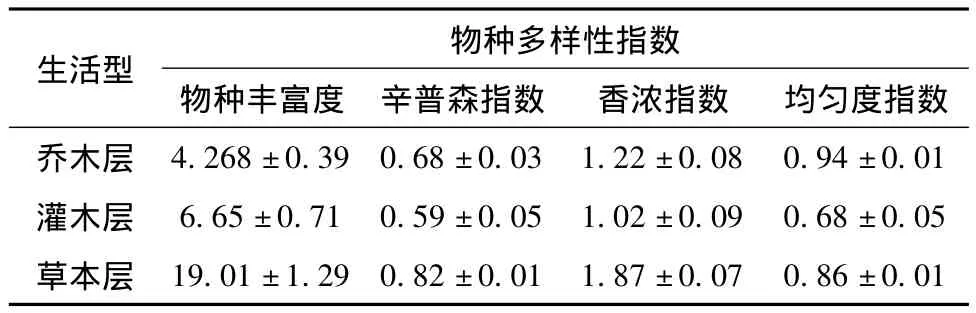
表1 帽儿山林区紫椴群落不同生长型物种多样性指数和均匀度指数Tab.1 Diversity and evenness indexes ofdifferent growth forms of Tilia amurensis community in Maoer mountain
3.2 种对间关联度分析
帽儿山林区紫椴群落有12个优势种对,其中有7个种对是正关联,占总种对数的58%,分别是紫椴-红松、紫椴-五角槭、紫椴-茶条槭、紫椴-卫矛、紫椴-裂叶榆、紫椴-暴马丁香、紫椴-糠椴(见表2)。紫椴种群是红松种群的伴生种群,种群格局随发育阶段呈由强至弱的聚集分布,主要是因为紫椴幼苗期易受霜冻危害,尽管冻后也能萌发,但会造成无明显主干,因此自然更新受到严重影响。

表2 帽儿山紫椴群落优势种间关联指数Tab.2 Interspecific relationship of dominant species in Tilia amurensis community
紫椴种对间极显著正关联有4对(0.01<P<0.05,ad>bc),紫椴-红松、紫椴-卫矛、紫椴-暴马丁香、紫椴-糠椴,显著正关联种对有1对(0.01<P<0.05,ad<bc)紫椴-裂叶榆,如图1所示。说明该树种对生境的要求和适应具有同一性或相似性,因此,优势种在相对稳定的群落中可以共同利用群落的非限制性资源,形成显著的正关联。
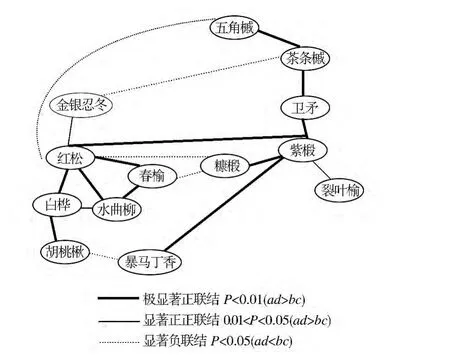
图1 紫椴群落种群间联结X2检验星座Fig.1 Constellation diagrams of X2-test of interspecific correlation in Tilia amurensis community
Ochiai指数指一个物种看作是另一个物种正常环境的一部分;Dice指数表示一个物种出现而另一物种也存在的概率。从表1可知,紫椴-五角槭、紫椴-卫矛、紫椴-糠椴,它们的OI值、DI值和JI值均在0.5,这是由于这些种群共同适应并占据着相近生境的缘故。OI值、DI值和JI值在0.3区间的种对有6对,分别是紫椴-白桦、紫椴-春榆、紫椴-水曲柳、紫椴-裂叶榆、紫椴-暴马丁香、紫椴-胡桃楸,说明这些种对的联结程度稍高,其它种对的联结程度则较低。
X2值不仅能检验联结的显著性,而且可用以种群间联结的测度,它在一定程度上衡量了种群间的相互关联和植物对环境综合生态因子反应的差异[7]。由图1可以看出,如本区紫椴种群出现时,可能有五槭、茶条槭、裂叶榆、暴马丁香、糖椴出现,这正是紫椴种群表现为顶级群落伴生种群的原因之一。
4 结果与讨论
帽儿山林区紫椴群落垂直结构物种多样性指数为:丰富度指数,草本层>灌木层>乔木层;辛普森指数,草本层>乔木层>灌木层。乔木层物种丰富度较低,草本层相对来说物种丰富并且分布相对较为均匀,灌木层物种尽管较为丰富。灌木层易受人为干扰(红松球果采摘、火灾)的影响,大部分物种个体分布不均匀,因此辛普森指数较低。该群落地带性植被为阔叶红松林,经过长期破坏和大肆开发利用(剩余的原始植被不足10%),导致原生植被已不复存在[8]。紫椴群落平均海拔325 m,郁闭度为0.7,坡向为南坡,土壤肥沃,更有利于草本生长发育。帽儿山林区是原始阔叶红松林分布区,原始阔叶红松林遭破坏后,种植人工林,由于种间竞争和更新能力不同形成不同的森林类型,立地质量的差异导致紫椴群落结构的差异[9]。
植物群落是土壤基本属性的综合指标,而特定的土壤、气候和地形条件孕育了不同的植物群落[10-12]。紫椴群落香浓指数表现为:草本层>乔木层>灌木层。由于人为经营活动和火烧干扰,灌木层物种个体均匀性降低,因此,灌木层香浓指数最小。
土壤是植物赖以生存和植被恢复的重要物质基础,因此土壤理化性质的变化直接影响植被的变化,相反,植被的演替进化可以影响着土壤的发育[13-14]。表3是多样性指数与环境因子关系,物种多样性指数中物种丰富度指数与土壤pH值明显相关,辛普森指数与海拔高度和土壤pH显著负相关,反映物种多度和物种数量的均匀程度的综合指数;香浓指数与海拔高度、有机质含量有明显相关,紫椴对土壤条件要求相对严格,多分布于山中、下腹的砂质壤或壤土上,特别适宜生长在土层深厚,排水良好的砂壤土上。紫椴群落均匀度指数与海拔、有机质含量、土壤总氮明显相关。
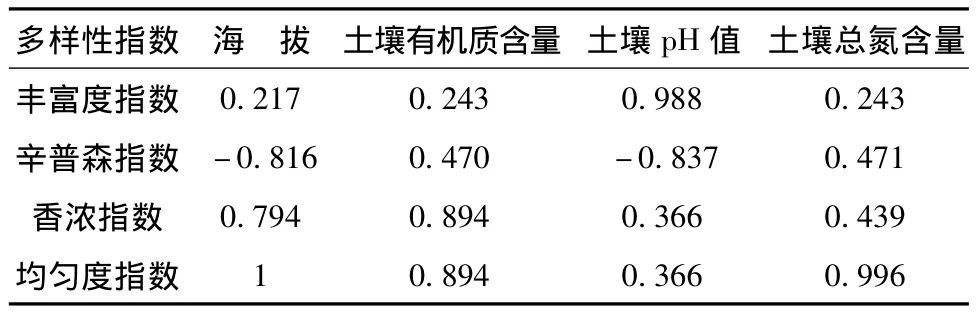
表3 物种多样性指数与环境因子的相关系数rTab.3 Correlation between species diversity and environmental factors
紫椴是红松的主要伴生树种,在不同生长阶段往往表现出不同的空间格局[15],Ochiai指数是一个物种看作是另一个物种正常环境的一部分。Dice表示一个物种出现而另一物种也存在的概率。相邻植物之间的竞争作用影响到植物本身生长发育、种群动态、群落演替和物种多样性,物种间相互作用与环境差异对植物分布的影响通过种间联结来反映[16]。帽儿山林区紫椴与五角槭、茶条槭、裂叶榆、暴马丁香、糠椴之间为正联结关系,这是它们与所分布地区的生态环境条件长期适应、形成的共憩促进关系。帽儿山紫椴群落Ochaia指数和Dice指数与土壤总氮含相关系数达0.938,0.940,说明紫椴种群分布与土壤含氮状况息息相关,见表4。
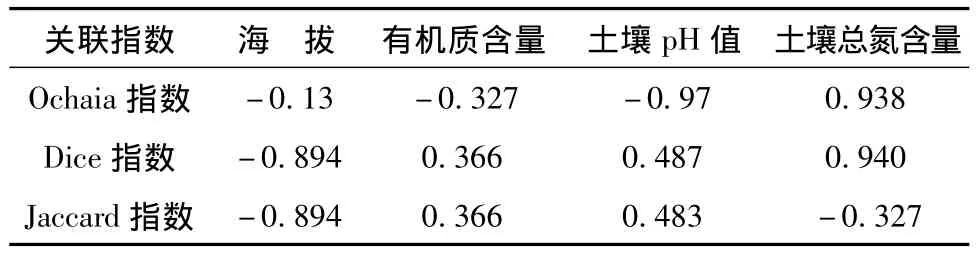
表4 种间相关指数与环境因子的相关系数Tab.4 Correlation between correlation index and environmental factors
群落中植物的生长、发育的各个阶段、都不同程度地受到环境和遗传基因的制约与支配,因而会产生不同程度的聚集和相关关联,所以测定不同个体在空间联结的程度,对于研究群落组成与动态及种群间的相互作用都具有重要意义[17-20]。帽儿山林区以人工林为主,人工林天然化发育的理想状态是目的树种生长正常,能够自我更新,物种多样性丰富,病虫危害少。处于不同发育阶段的紫椴种群与其它地带性植被存在相似性,紫椴群落物种多样性指数与种间关联指数相关性达到了较高水平,这是因为经过前期发育过程中的激烈竞争与遴选,一些物种被淘汰,余下的物种也分别找到了各自合适的位置,彼此间互惠共存,达到了相对稳定阶段。
[1]崔 磊,穆立蔷.黑龙江省紫椴典型分布区外生菌根形态与解剖特征[J].生态学杂志.2014,22(9):2490-2500.
[2]聂绍荃.紫椴种群生态学研究[M].哈尔滨:东北林业大学出版社,1992.
[3]郭忠玲,马元丹,郑金萍.长白山落叶阔叶混交林的物种多样性、种群空间分布格局及种间关联性研究[J].应用生态学报,2004,15(11):2013-2018.
[4]张 玲,袁晓颖,张东来.帽儿山落叶松群落主要树木种群间联结关系的研究[J].北京林业大学学报,2008,30(4):141-145.
[5]张 玲,袁晓颖,张东来.大、小兴安岭植物区及交错区物种多样性比较研究[J].植物研究,2007,27(3):236-360.
[6]王伯荪.植物群落学[M].北京:高等教育出版社,1987.
[7]郑超超,伊力塔,张 超,等.浙江江山公益林物种种间关系及CCA 排序[J].生态学报,2015,35(22):1-11.
[8]郑雪峰,侯照云.帽儿山林区次生林物种多样性分析[J].林业科技情报.2010,42(4):6-7.
[9]丁 壮.东北林业大学帽儿山实验林场原始红松林的破坏与恢复的雏仪[J].植物研究,2013,33(3):379-384.
[10]郭逍宇,张金屯,宫辉力,等.安太堡矿区复垦地植被种间关系及土壤因子分析[J].生物多样性,2007,15(1):46-52.
[11]宋创业,郭 柯.浑善达克沙地中部丘间低地植物群落分布与土壤环境关系[J].植物生态学报,2007,31(1):40-49.
[12]王顺忠,陈桂琛,柏玉平,等.青海湖鸟岛地区植物群落物种多样性与土壤环境因子的关系[J].应用生态学报,2005,16(1):186-188.
[13]赵景学,曲广鹏,多吉顿珠,等.藏北高寒植被群落物种多样性与土壤环境因子的关系[J].干旱区资源与环境,2011,25(6):105-108.
[14]张 玲,王承义.大小兴安岭过渡区阔叶红松林次生演替阶段群落多样性指数与环境因子关系[J].森林工程,2014,30(5):1-5.
[15]吴初平,张 骏,沈爱华,等.千岛湖次生林不同演替阶段林分空间结构研究[J].植物研究,2015,35(1):16-21.
[16]史作民,刘世荣,程瑞海,等.宝天曼落叶阔叶林种间联结性研究[J].林业科学,2001,37(2):29-35.
[17]贾呈鑫卓,李帅锋,苏建荣,等.择伐对思茅松天然林乔木种间与种内关系的影响[J].植物生态学报,2014,38(12):1296-1306.
[18]严令斌,余德会,李 鹤,等.赤水河清香木群落主要树种种间关系与生态种组[J].西部林业科学,2015,44(1):69-75.
[19]高金辉,张厚良,陈海波,等.小兴安岭天然阔叶混交林群落多样性特征[J].林业科技,2014,39(5):30-33.
[20]许林红,李思广,蒋云东,等.西南桦、山桂花、肉桂人工林林下物种多样性和生物量对比研究[J].四川林业科技,2014,35(6):51-55.
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
河南科学(2020年3期)2020-06-02
蔬菜(2018年5期)2018-05-17
乡村科技(2018年8期)2018-02-13
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
杂草学报(2015年2期)2016-01-04
草业科学(2015年9期)2015-12-11
