
牛LF基因结构和功能的生物信息学分析
2016-06-21 03:53宁素恒李强子
浙江农业科学 2016年6期
关键词:生物信息学
宁素恒,徐 军,李强子,张 丽
(1.宁夏农垦贺兰山奶业有限公司,宁夏银川 750024;2.西北民族大学生命科学与工程学院,甘肃兰州 730030)
牛LF基因结构和功能的生物信息学分析
宁素恒1,徐 军1,李强子2,张 丽2
(1.宁夏农垦贺兰山奶业有限公司,宁夏银川 750024;2.西北民族大学生命科学与工程学院,甘肃兰州 730030)
摘 要:本试验采用生物信息学方法预测和分析牛乳铁蛋白基因(LF)编码产物的理化特性、结构功能、信号肽、N-糖基化位点及其同源进化关系,以期为牛的抗病分子育种和经济性状相关基因筛选提供理论依据。研究结果显示牛LF基因编码708个氨基酸,产物为一种不稳定的水溶性蛋白,二级结构以a-螺旋为主,还有6个糖基化位点。蛋白功能以信号转导、胁迫应答和免疫应答的概率相对较高,这表明LF基因可能在牛免疫应答和生长调节过程中发挥重要作用,能对牛的抗病力和生产性能起作用。同源进化关系来看,牛LF基因与马的亲缘关系最近。因此LF基因可作为荷斯坦牛抗病育种的候选基因进行进一步研究。
关键词:牛;LF基因;抗病育种;生物信息学
文献著录格式:宁素恒,徐军,李强子,等.牛LF基因结构和功能的生物信息学分析[J].浙江农业科学,2016,57(6): 904-907.
徐 军(1978—),男,宁夏青铜峡人,畜牧师,本科,从事奶牛育种与疾病防治研究工作,E-mail: 409967130@qq.com。
乳铁蛋白(lactoferrin,LF)是一种天然活性铁结合糖蛋白,广泛存在于动物的乳汁、血液、唾液、小肠液等外分泌液中,具有抑菌、抗病毒感染、免疫调控、激活免疫应答及溶菌酶再生等生理功能[1-2]。将乳铁蛋白运用于动物生产中可以显著降低动物缺铁性贫血发病率、提高动物免疫力和改善肠道微生态环境,促进动物健康生长。牛LF基因被定位于牛的第22号染色体上[3]。研究发现牛的乳铁蛋白浓度可遗传(h2=0.4)[4]。Molenaar等[5]研究表明患有乳腺炎的牛乳腺组织中LF基因的表达量比未患有乳腺炎的牛群体高,说明LF基因表达与炎症反应相关联。此外,Dinesh等[6-10]通过研究均表明牛LF基因多态性与牛的泌乳性状和乳腺炎抗性密切相关。基于LF基因的可遗传性以及与乳腺炎的相关性,可以对LF基因的结构和功能进行进一步的深入研究。
为深入探讨牛LF基因的理化性质和生物学功能,避免该基因演化造成的差异,本研究利用生物信息学方法对牛LF基因序列结构及相关生物信息学特征进行预测和分析,同时构建系统发育进化树,为揭示该基因理化性质、功能信息、遗传特性及相关生理机制提供理论基础。
1 材料与方法
1.1 牛LF基因序列
从GeneBank中查找得到牛的LF基因的编码区序列(Codingsequences,CDS)(Genebank no.NC-007320)。
1.2 方法
对牛LF基因CDS编码产物的氨基酸序列、其蛋白质特性及功能结构域进行分析与预测。所用工具软件与网址为开放阅读框分析(http://www.ncbi.nlm.nih.gov/gorf/gorf.html),编码蛋白质理化质分析(http://web.expasy.org/protparam/),编码蛋白质疏水性/亲水性分析(http://web.expasy.org/protscale/),编码蛋白质的N-糖基化位点分析(http://www.cbs.dtu.dk/services/NetNGlyc/),编码蛋白质二级结构预测(http://npsa-pbil.ibcp.fr/cgi-bin/npsa-automat.pl?page=npsa-sopma.html),编码蛋白质三级结构(http://swissmodel.expasy.org/和PyMOL软件),编码蛋白质的信号肽跨膜区预测(http://www.cbs.dtu.dk/services/SignalP/),编码蛋白质的功能预测(http://www.cbs.dtu.dk/services/ProtFun/),同源系统发育进化树分析(DNAStar软件中的MegAlign程序)。
2 结果与分析
2.1 牛LF基因序列的开放阅读框(ORF)
开放阅读框为基因从起始密码子到终止密码子之间的一段核苷酸序列。牛LF基因ORF分析结果如图1,全长为2 127 bp。5′端起始密码子是ATG,3′端终止密码子是TAA,编码蛋白质由708个氨基酸组成。

图1 牛LF基因CDS区的ORF分析
2.2 牛LF基因编码氨基酸理化性质的预测
牛LF基因编码708个氨基酸,20种氨基酸所占比例亮氨酸(Leu)数目最多(表1),占整个氨基酸组成的10.3%,甲硫氨酸(Met)数目最少,占0.7%,负电荷残基总数(Asp + Glu)为76,正电荷残基总数(Arg+Lys)为92,其分子式为C3449H5437N965O1022S40,分子质量为10.913 ku,半衰期为30 h,基因编码产物不稳定指数为40.99,大于40,表明该基因编码产物不稳定[11]。
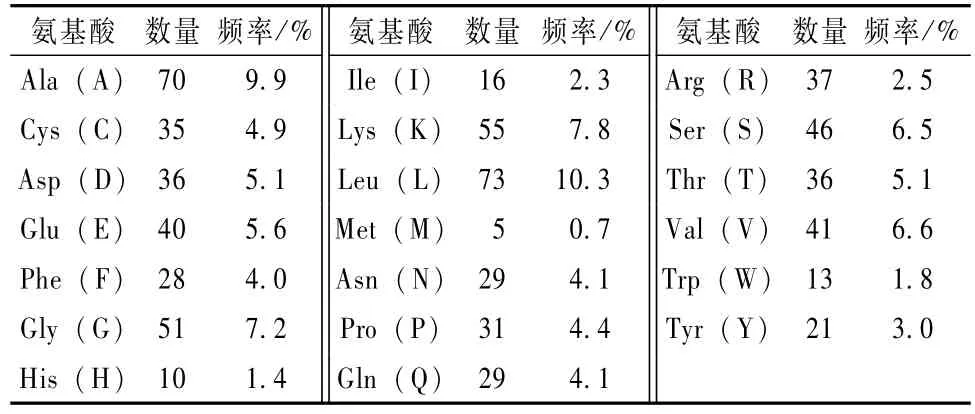
表1 牛LF编码蛋白的氨基酸组成
2.3 牛LF基因编码蛋白质的理化性质
牛LF基因编码氨基酸序列亲水性/疏水性预测结果如图2所示,其中第7位丙氨酸(Ala)疏水性最强(+2.400);第243位的天冬氨酸(Asp)亲水性最强(-2.882)。整条氨基酸肽链中,亲水氨基酸占62.15%,疏水氨基酸占37.85%,总平均疏水性为-0.328,表现为亲水性,因此,可推断牛LF基因编码的蛋白质是一种可溶性蛋白。
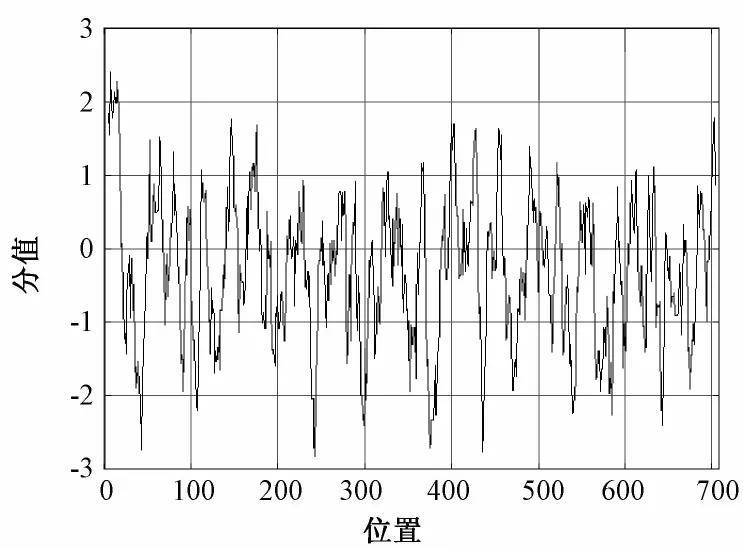
图2 牛LF基因编码蛋白的疏水性预测结果
2.4 牛LF基因编码蛋白质N-糖基化位点的预测
蛋白质糖基化是真核生物蛋白质翻译后加工的重要修饰之一,蛋白质糖基化修饰对蛋白质折叠、分选及其定位有重要影响,使用在线软件http://www.cbs.dtu.dk/services/NetNGlyc/对LF基因N-糖基化位点进行预测,结果如图3所示。牛LF基因编码蛋白氨基酸序列中存在6个潜在的N-糖基化位点。
2.5 牛LF基因编码蛋白质的二级结构和三级结构预测
使用在线软件http://npsa-pbil.ibcp.fr/cgibin/npsa-automat.pl?page=npsa-sopma.html对牛乳铁蛋白二级结构组分进行分析,其中a-螺旋(Hh)占34.89%,β-折叠(Ee)占19.49%,β-转角(Tt)占11.44%,无规则卷曲(Cc)占34.18%。乳铁蛋白二级结构中,Hh占主导地位。三级结构见图4,从预测结果来看,三级结构主要成分为Hh,与二级结构预测结果一致。
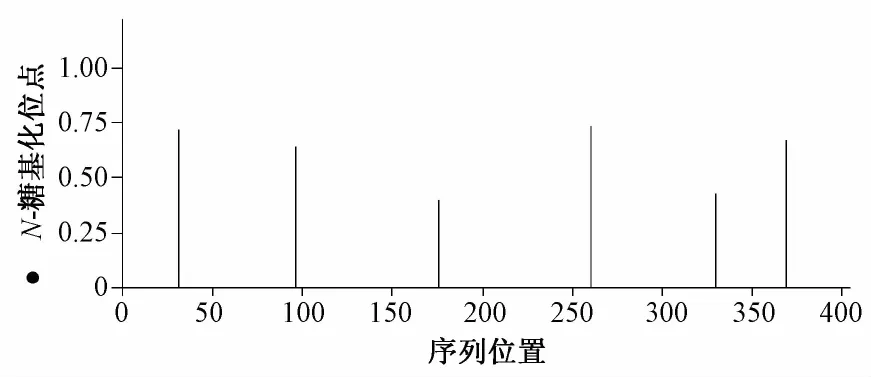
图3 荷斯坦牛LF基因编码蛋白N-糖基化位点
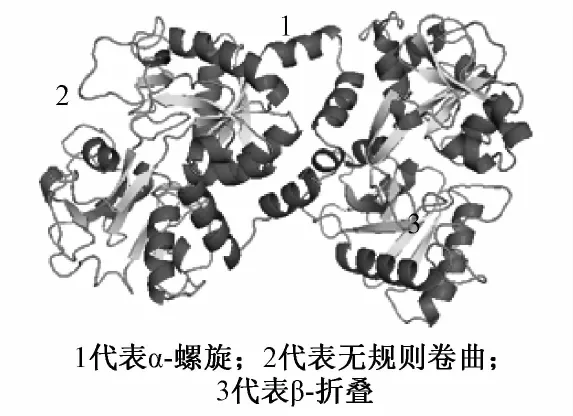
图4 荷斯坦牛LF基因编码蛋白的三级结构模型预测
2.6 牛LF基因编码蛋白质的功能预测
牛LF基因编码蛋白质的功能预测见表2,该蛋白信号转导、胁迫应答和免疫应答的概率相对较高,分别为0.985,0.830和0.357,这表明LF基因可能在牛免疫应答和生长调节过程中发挥重要作用,可能对牛的抗病力和生产性能起作用。
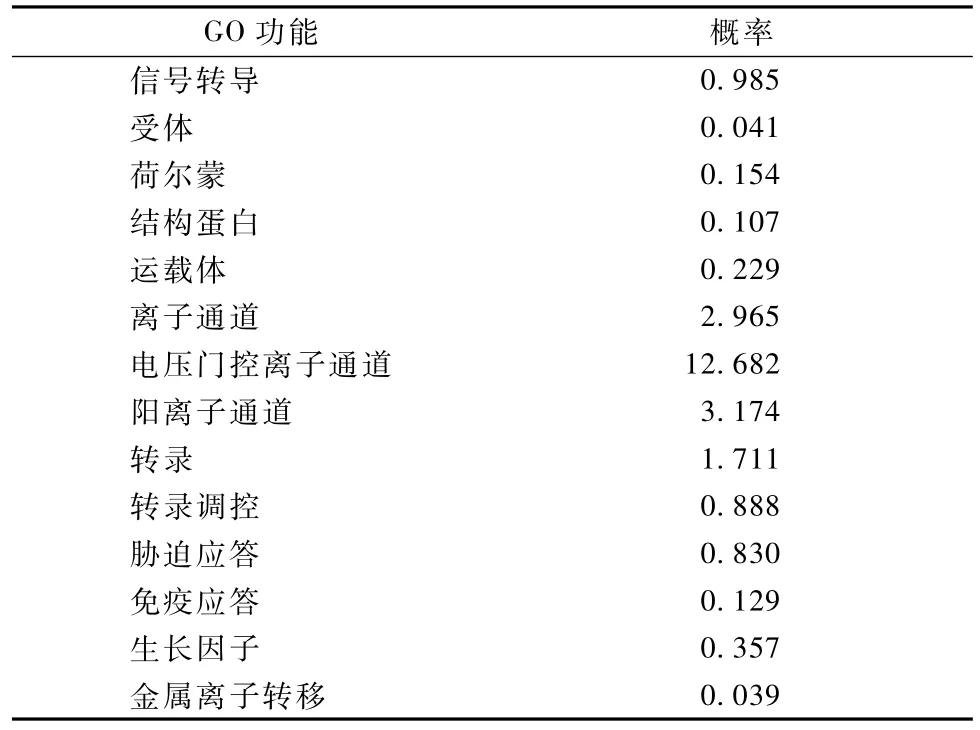
表2 牛LF基因编码蛋白质的功能预测
2.7 牛LF基因编码蛋白质信号肽跨膜区的预测
信号肽是决定新生肽链在细胞中的定位或决定某些氨基酸残基修饰的一些肽段。在线软件http://www.cbs.dtu.dk/services/SignalP/分析结果显示(图5),牛LF基因编码产物C值为0.728,Y值为0.819,S值为0.961,其切割点位于19~20位的氨基酸之间,基本可以判定LF基因编码产物存在信号肽。
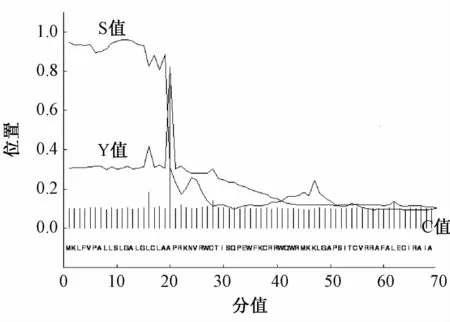
图5 荷斯坦牛LF基因编码蛋白质信号肽跨膜区
2.8 牛LF基因编码蛋白的同源序列分析
使用DNAStar软件对6个物种的LF基因编码产物序列进行同源性分析。由图6可知,LF基因几乎在所有的哺乳动物中都有表达。从进化树分析结果来看,进化树分为两支,其中灵长目、偶蹄目和奇蹄目聚为一类,啮齿目聚为一类。其中牛LF基因亲缘关系与马最近,这一结果符合动物分类学分类标准。
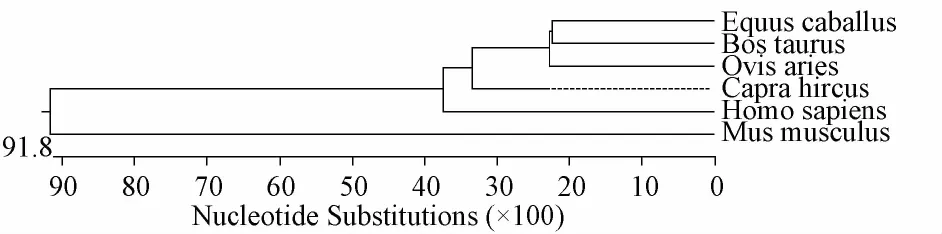
图6 荷斯坦牛LF基因编码蛋白质的氨基酸系统进化树
3 讨论
在哺乳动物中,LF基因是基因组中多态性较丰富的区域,对LF基因遗传变异进行生物分析可以提供物种的遗传多样性信息,对动物种质资源的保护具有重要的生物学意义。同时LF基因与牛的繁殖性状存在关联性[12]。因此对LF基因的生物信息学分析也对实际生产有一定的帮助。
蛋白质结构决定其功能特点,进而影响其生命活动的代谢。牛LF基因CDS区序列全长2 127 bp,编码708个氨基酸,编码产物为不稳定蛋白。研究表明,蛋白质半衰期越长则蛋白质稳定性越高[13],而本研究发现LF编码产物氨基酸具有较长的半衰期(30 h)却是不稳定蛋白,出现这一特性可能与其不同生理功能的发挥存在某种联系。糖基化是真核生物蛋白质翻译后重要的修饰之一,它可以影响蛋白质的抗原决定簇、电荷性质、热稳定性、免疫分子的结构和功能,从而影响机体对抗原的应答反应[14],它还可以调控蛋白质在组织和细胞中的定位、功能和活性[15],糖基化位点发生变化可能与疾病的发生有关[16],因此荷斯坦牛LF基因蛋白上潜在的糖基化位点可能会影响基因与病原相关分子的连接绑定,从而影响机体对乳腺炎的抵抗力。研究表明,无规则卷曲易受侧链相互影响而改变空间构象,a螺旋和无规则卷曲是蛋白质肽链中构成配体受体结合的活性部位[17],LF基因编码蛋白二级结构中大量的无规则卷曲可能影响蛋白质肽链的结合活性,从而影响蛋白质的功能。荷斯坦牛LF基因编码蛋白的信号转导、胁迫应答、免疫应答和生长因子的概率相对较高,其中免疫应答和生长因子的概率最高,说明该基因可能对荷斯坦牛的免疫力和生产力具有较高的调节能力;LF基因可能在荷斯坦牛免疫应答过程中发挥重要作用,可能对荷斯坦牛的抗病力起作用[18]。
为了保证研究结果的准确性,本研究对各类型的预测均选用了多种不同软件,不同算法,并进行多次预测。预测所得结果一致性较高,具有较高的可信度。对牛LF基因的研究将有利于对该基因及其编码蛋白功能的进一步分析,了解牛LF基因与相关疾病的遗传效应可以为利用LF基因改良牛经济性状提供参考。
参考文献:
[1] ORSIN.Theantimicrobialactivityoflactoferrin: currentstatus and perspectives[J].BiometalsAn InternationalJournalon theRoleofMetalIonsin BiologyBiochemistry& Medicine,2004,17(3): 189-196.
[2] BELJAARS L,WIEGMANS FC,HARMSEN M C,etal.Inhibition ofcytomegalovirusinfection bylactoferrin in vitroand in vivo[J].AntiviralResearch,2004,63(3): 197-208.
[3] SEYFERTH M,TUCKORICZ A,INTERTHAL H,etal.Structure ofthe bovine lactoferrin-encoding gene and its promoter[J].Gene,1994,143(2): 265-269.
[4] KAMINSKIS,OLENSKIK,BRYMP,etal.Singlenucleotide polymorphismin thepromoterregion ofthelactoferrin geneand itsassociationswith milk performancetraitsin polish Holsteinfriesiancows[J].Genetika,2006,42(8): 1117-1120.
[5] MOLENAARAJ,KUYS YM,DAVIS S R,etal.Elevation of lactoferrin geneexpression in developing,ductal,resting,and regressingparenchymalepithelium oftheruminantmammary gland[J].JournalofDairy Science,1996,79(7): 1198-1208.
[6] DINESHK,VERMAA,GUPTAID,etal.Identification of polymorphism in exons7 and 12 oflactoferrin geneand its association with incidenceofclinicalmastitisin Murrah buffalo [J].TropicalAnimalHealth & Production,2015,47(4): 643-647.
[7] CARVAJALAM,HUIRCANP,LEPORIA.Singlenucleotide polymorphismsin immunity-related genesand theirassociation with mastitis in Chilean dairy cattle[J].Genetics and MolecularResearch,2013,12(3): 2702-2711.
[8] SINGH A P,RAMESHA K P,ISLOOR S,etal.Single NucleotidePolymorphismsin Lactoferrin GeneareAssociated with Lactoferrin Contentin Milk and SomaticCellCountin Deoni(Bosindicus)Cows[J].Pakistan VeterinaryJournal,2015,35(3): 303-308.
[9] 陈仁金,王珍珍,毛永江,等.中国荷斯坦牛乳铁蛋白基因外显子1多态性与泌乳性状及体细胞评分的关联分析[J].畜牧与兽医,2013,5(9): 53-56.
[10] O' HALLORAN F,BERRY D P,BAHAR B,etal.Polymorphismsin thebovinelactoferrin promoterareassociated with reproductiveperformanceand somaticcellcount[J].JournalofDairyScience,2010,93(3): 1253-1259.
[11] GURUPRASADK,REDDYBV,PANDITM W.Correlation between stabilityofaprotein and itsdipeptidecomposition: a novelapproach forpredictingin vivostabilityofaprotein from itsprimarysequence[J].Protein Engineering,1990,4(2): 155-161.
[12] HAJIBEMANIA,SHARIFIYAZDIH,MIRZAEIA,etal.Characterization ofsinglenucleotidepolymorphism in the5′-untranslated region(5′-UTR)ofLactoferrin gene and its association with reproductiveparametersand uterineinfection in dairycattle[J].VeterinaryResearch Forum,2012,3(1): 37-43.
[13] 贾浩,张小白,宋晓峰.人类胞内蛋白半衰期与其亚细胞定位的相关性研究[J].计算机与应用化学,2011,28 (4): 411-414.
[14] CALARESED A,SCANLAN C N,ZWICK M B,etal.Antibodydomain exchangeisan immunologicalsolution to carbohydrateclusterrecognition[J].Science,2003,300 (5628): 2065-2071.
[15] ARNOLDJN,WORMALDM R,SIM RB,etal.Theimpact ofglycosylation on thebiologicalfunction and structureofhuman immunoglobulins[J].AnnualReviewofImmunology,2007,25: 21-50.
[16] LITYNSKA A,PRZYBYUULO M,POCHEC E,et al.Comparison ofthelectin-bindingpattern in differenthuman melanomacelllines[J].MelanomaResearch,2001,11(3): 205-212.
[17] 郑海军,朱荣,葛春蕾,等.人白细胞介素-29的生物信息学分析[J].中国生物制品学杂志,2013,26(2): 209-212,217.
[18] 徐飞,成述儒,罗玉柱.绵羊DRB1基因生物信息学分析[J].生物技术通报,2011(1): 113-118.
(责任编辑:卢福庄)
中图分类号:S823
文献标志码:A
文章编号:0528-9017(2016)06-0904-04
DOI:10.16178/j.issn.0528-9017.20160635
收稿日期:2016-02-15
基金项目:西北民族大学引进人才科研项目(xbmuyjrc201316);西北民族大学国家级大学生创新创业训练计划项目(201510742084)
作者简介:宁素恒(1984—),男,山西榆社人,助理畜牧师,本科,从事奶牛育种与繁殖工作,E-mail: 107154324@qq.com。
通信作者:张 丽,E-mail: shiningstar2013@sina.cn。
猜你喜欢
中国中药杂志(2016年22期)2017-02-13
中国校外教育(下旬)(2016年11期)2016-12-27
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
电脑知识与技术(2016年27期)2016-12-15
科技视界(2016年23期)2016-11-04
考试周刊(2016年78期)2016-10-12
课程教育研究·学法教法研究(2016年10期)2016-06-16
安徽农学通报(2015年20期)2015-11-07
现代电子技术(2015年11期)2015-07-28
