
11β-羟基类固醇脱氢酶1型抑制剂对高脂血症大鼠肝脏、脂肪和骨骼肌脂代谢基因的影响
2016-12-17 07:57吴国琼郭菲菲杨艳群候亚莉邓春地袁惠萍
中国老年学杂志 2016年22期
吴国琼 林 梅 郭菲菲 杨艳群 张 雨 候亚莉 邱 璇 邓春地 袁惠萍
(华中科技大学同济医学院附属武汉普爱医院内分泌科,湖北 武汉 430034)
11β-羟基类固醇脱氢酶1型抑制剂对高脂血症大鼠肝脏、脂肪和骨骼肌脂代谢基因的影响
吴国琼 林 梅 郭菲菲 杨艳群 张 雨 候亚莉 邱 璇 邓春地 袁惠萍
(华中科技大学同济医学院附属武汉普爱医院内分泌科,湖北 武汉 430034)
目的 探讨11β-羟基类固醇脱氢酶1型(11β-HSD1)抑制剂BVT 2733对高脂血症大鼠肝脏、脂肪和骨骼肌脂代谢基因表达的影响。方法 采用高脂饮食喂养建立高脂血症大鼠模型。将40只高脂血症大鼠随机分为模型组和BVT 2733(20 mg/kg)组,每组20只。另取20只普通饲料喂养的SD大鼠作对照组。分别观察BVT 2733对甘油三酯(TG)、总胆固醇(TC)、低密度脂蛋白(LDL-C)、高密度脂蛋白(HDL-C)水平的影响,并测定肝脏和肾周、大网膜及睾周脂肪组织脂肪含量,测定腓肠肌组织中Rev-erba、FAT/CD36、SCD1及CPT1 mRNA的表达水平。结果 BVT 2733治疗后,高脂血症大鼠的TG、TC和LDL-C明显低于模型组(P<0.05),并且肝脏和内脏脂肪的脂肪含量明显低于模型组(P<0.05),腓肠肌组织中Rev-erba mRNA水平明显低于模型组(P<0.05),FAT/CD36、SCD1及CPT1 mRNA水平明显高于模型组(P<0.05)。结论 BVT 2733可改善高脂血症大鼠的血脂及脂蛋白水平紊乱,降低肝脏脂肪含量并稳定骨骼肌脂代谢基因表达,提示抑制11β-HSD1活性对高脂血症相关的肝脏、脂肪组织和骨骼肌异常脂质代谢有一定改善作用。
11β-羟基类固醇脱氢酶1型抑制剂;高脂血症;肝脏脂质代谢;骨骼肌脂代谢基因
肥胖人群中高脂血症的发病率高达50%以上〔1〕。高脂血症可增加冠心病、高血压、动脉硬化性血管病、2型糖尿病的发生和病死率。肥胖症的脂代谢异常发生机制较多。国内外学者仅分别就11β-羟基类固醇脱氢酶1型(11β-HSD1)在肥胖症、2型糖尿病患者及动物模型中的作用及相关机制做过初步研究,证实在脂代谢异常时11β-HSD1参与高脂血症的形成〔2~4〕,但11β-HSD1在高脂血症模型肝脏及骨骼肌脂代谢中的作用尚不清楚。本研究拟探讨11β-HSD1抑制剂BVT 2733对高脂血症大鼠肝脏脂代谢及骨骼肌脂代谢基因表达的影响。
1 材料与方法
1.1 主要材料 11β-HSD1抑制剂BVT 2733购自美国MCE公司,油红 O、苏木素-伊红(HE)染色液购自美国Sigma公司,QuantiTect SYBR Green PCR、QuantiTect Probe PCR kits试剂盒购自德国Qiagen公司,β-actin(NM_013200)、Rev-erbα(NM _145775)、SCD1(NM _139192)、FAT/CD36(NM_031561)、CPT1(NM_013200)引物均购自上海生工公司。
1.2 动物分组及大鼠模型建立 选取200~220 g SD雄性大鼠40只,采用高脂饮食饲养6 w诱导高脂血症大鼠模型。将这40只模型大鼠随机分为模型组和BVT 2733组,每组20只,其中BVT 2733组每天接受20 mg/kg BVT 2733灌胃处理,连续4 w,而模型组仅接受等体积生理盐水灌胃处理。另取20只普通饲料喂养的SD大鼠作对照组。本研究所用大鼠购自上海斯莱克实验动物有限公司。
1.3 血脂及脂蛋白水平检测 治疗4 w后腹主动脉取血5 ml,加入不含抗凝剂的离心管中,静置10 min后分离血清,采用Olympus AU 400型全自动生化仪检测血清甘油三酯(TG)、总胆固醇(TC)、低密度脂蛋白(LDL-C)、高密度脂蛋白(HDL-C)、载脂蛋白(Apo)AI和 Apo B水平。
1.4 肝脏组织病理学观察 取新鲜肝脏组织3~4块,修剪成1.0 cm×1.0 cm×1.0 cm的组织块,采用OTC包埋剂包埋,常规油红O染色后,采用Image-Pro Plus 5.0图像处理和分析软件分析肝脏脂肪含量。
1.5 脂肪组织重量与HE 染色观察 采集肾周、大网膜和睾周脂肪组织,采用滤纸吸干表面附着组织液后称量各部分重量。将取得的脂肪组织,用 4%多聚甲醛缓冲液固定后,依次经脱水、透明、包埋及染色后,采用Image-Pro Plus 5.0软件计算脂肪细胞平均面积。
1.6 骨骼肌组织中脂代谢相关基因的表达情况 分离大鼠腓肠肌,将其置于EP管(1.5 ml)中,置于液氮中研磨,提取骨骼肌组织中总RNA,采用荧光实时定量PCR法检测Rev-erbα、SCD1、FAT/CD36及CPT1 mRNA的表达情况。按照QuantiTect SYBR Green PCR试剂盒说明书操作扩增。
1.7 统计学分析 采用SPSS20.0软件,两组间均数比较采用t检验,多组间均数比较采用单因素分析。
2 结 果
2.1 各组大鼠的血脂水平比较 与对照组比较,模型组的TG、TC及LDL-C水平均升高,HDL-C水平降低(P<0.05);BVT 2733处理后TG、TC及LDL-C水平均低于模型组,HDL-C水平高于模型组(P<0.05),且与对照组仍有统计学差异(P<0.05)。见表1。
2.2 各组大鼠的载脂蛋白水平比较 与对照组比较,模型组的Apo AI、Apo AI/Apo B水平降低,Apo B水平升高(P<0.05);BVT 2733处理后的Apo AI、Apo AI/Apo B高于模型组,Apo B水平低于模型组(P<0.05),但与对照组仍有统计学差异(P<0.05)。见表2。
2.3 各组大鼠的肝脏脂肪含量比较 对照组的肝脏脂肪含量比为(0.17±0.03)%,低于模型组的(32.59±4.23)%(P<0.05);BVT 2733处理后肝脏脂肪含量比为(19.82±2.47)%,低于模型组(P<0.05),但仍高于对照组(P<0.05)。
2.4 各组大鼠脂肪分布比较 与对照组比较,模型组的肾周、大网膜、睾周脂肪垫重量及脂肪总量均升高(P<0.05);BVT 2733处理后肾周、大网膜、睾周脂肪垫重量及脂肪总量均低于模型组(P<0.05),且与对照组仍有差异(P<0.05)。见表3。
2.5 各组大鼠脂肪细胞形态学观察 模型组的脂肪细胞面积为(15.72±0.87)×104μm2,高于对照组的(11.39±1.26)×104μm2(P<0.05);BVT 2733组的脂肪细胞面积为(12.93±1.53)×104μm2,低于模型组,但仍高于对照组(P<0.05)。
2.6 各组大鼠骨骼肌代谢基因表达情况比较 与对照组比较,模型组骨骼肌中Rev-erba mRNA水平降低,FAT/CD36、SCD1及CPT1 mRNA水平升高(P<0.05);BVT 2733处理后Rev-erba mRNA水平高于模型组,其余3个基因的mRNA水平低于对照组(P<0.05);除CPT1外,BVT 2733组骨骼肌的Rev-erba、FAT/CD36及SCD1 mRNA水平与对照组仍有统计学差异(P<0.05)。见表4。
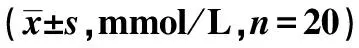
表1 各组大鼠的血脂水平比较
与对照组比较:1)P<0.05;与模型组比较:2)P<0.05;下表同
表2 各组大鼠Apo水平比较

表3 各组大鼠各部位脂肪垫重量及脂肪组织总量比较
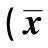
表4 各组大鼠骨骼肌代谢基因的表达情况±s,n=20)
3 讨 论
11β-HSD1是机体中重要的糖皮质激素代谢酶,可使无活性的可的松转变为有活性的氢化可的松,从而调节多种物质代谢〔5〕。我们前期的实验发现,高血糖可以直接促进胰岛β细胞上的11β-HSD1表达增加,与在肝细胞上作用类似,这种葡萄糖对11β-HSD1的直接作用提示葡萄糖可能是一个促进细胞内合成皮质酮的重要代谢信号,具有启动局部组织糖皮质激素代谢的作用〔6〕。研究表明11β-HSD1在调节脂肪细胞活动及脂肪代谢中发挥重要〔7~9〕,故针对11β-HSD1的治疗策略可能有一定效果。
本研究结果提示高脂血症模型制备成功。经BVT 2733处理后TG、TC、LDL-C水平降低,而HDL-C水平升高,提示抑制11β-HSD1活性具有良好的降血脂效果。脂蛋白在血脂调控中发挥重要作用,Apo B主要存在LDL-C表面,在介导细胞识别和摄取LDL-C中发挥重要作用〔10〕,而Apo AI为HDL-C的主要结构蛋白,Apo B及Apo AI水平可用于反映LDL-C和HDL-C水平〔11〕。本研究进一步发现抑制11β-HSD1活动对降低高脂血症大鼠Apo B水平及升高Apo AI水平有较好的效果,更加证实了11β-HSD1参与调控高脂血症大鼠脂质代谢的作用。
脂质代谢紊乱可引起肝脏细胞内脂肪过度蓄积,如不及时控制可引起肝脏脂肪化〔12〕。高脂血症大鼠接受BVT 2733处理后,肝脏脂肪比例降低,提示抑制11β-HSD1 可促进肝脏脂质代谢合成,改善脂质降解紊乱。此外,BVT 2733处理后高脂血症大鼠的各部位脂肪垫重量及脂肪组织总量较模型大鼠均降低,且脂肪细胞面积减少,为11β-HSD1调控高脂血症脂质代谢提供了直接证据。
骨骼肌是能量物质储存的主要外周组织,也是脂肪酸氧化代谢最活跃的部位之一。在骨骼肌脂质代谢过程,多个与脂代谢有关的基因参与其中。有研究〔13〕表明,骨骼肌中的11β-HSD1有代谢疾病缓解的作用,高血糖KK/Ta小鼠中,11β-HSD1抑制骨骼肌胰岛素受体底物mRNA表达增加,减少参与脂质代谢的基因。Rev-erba是脂代谢调节的核心作用因子,将其敲除后小鼠可出现脂代谢异常〔14〕。FAT/CD36是脂肪酸跨膜转运的关键转运体,可将循环脂肪酸转运至肌细胞,促进胞内脂质合成及脂肪的氧化代谢〔15〕。SCD1为多不饱和脂肪酸转化成单不饱和脂肪酸反应的关键限速酶,该反应可为肌细胞内脂质合成提供底物〔16〕。CPT1是脂肪酸β氧化的限速酶,在长链脂酰CoA的酯酰基转移至肉碱合成脂酰肉碱过程中发挥催化作用,最终转运至线粒体内膜参与β氧化〔17〕。本研究发现高脂血症大鼠肌细胞中脂质过载导致代谢异常,同时也增加了诱发胰岛素抵抗的风险;抑制11β-HSD1后以上指标获得改善,进一步提示11β-HSD1在维持肌细胞脂质动态平衡中起到重要作用,可能与其调节脂肪跨膜转运及氧化代谢等环节有关。
1 Zhang X,Yao S,Sun G,etal.Total and abdominal obesity among rural Chinese women and the association with hypertension〔J〕.Nutrition,2012;28(1):46-52.
2 计晨琳,孙 斌,韩素萍,等.不同年龄女性脂肪组织11β-HSD1 mRNA表达水平及临床意义〔J〕.中国老年学杂志,2012;32(18):3878-80.
3 邹显彤,纪立农.11β-羟基类固醇脱氢酶1型在代谢综合征中的作用〔J〕.中国糖尿病杂志,2015;23(4):378-80.
4 Walker GE,Marzullo P,Prodam F,etal.Obesity modifies expression profiles of metabolic markers in superficial and deep subcutaneous abdominal adipose tissue depots〔J〕.Endocrine,2014;46(1):99-106.
5 Gyllenhammer LE,Alderete TL,Mahurka S,etal.Adipose tissue 11βHSD1 gene expression,βcell function and ectopic fat in obese African Americans versus Hispanics〔J〕.Obesity (Silver Spring),2014;22(1):14-8.
6 林 梅,张木勋,张建华,等.葡萄糖上调胰岛β细胞系NIT-1 11β-羟类固醇脱氢酶1型的表达〔J〕.基础医学与临床,2009;23(3):296-300.
7 Koh EH,Kim AR,Kim H,etal.11β-HSD1 reduces metabolic efficacy and adiponectin synthesis in hypertrophic adipocytes〔J〕.J Endocrinol,2015;225(3):147-58.
8 Lai W,Tian X,Xiang Q,etal.11β-HSD1 modulates LPS-induced innate immune responses in adipocytes by altering expression of PTEN〔J〕.Mol Endocrinol,2015;29(4):558-70.
9 Pardina E,Baena-Fustegueras JA,Fort JM,etal.Hepatic and visceral adipose tissue 11βHSD1 expressions are markers of body weight loss after bariatric surgery〔J〕.Obesity (Silver Spring),2015;23(9):1856-63.
10 费 娜,刘 勇.短暂性脑缺血发作患者血清LP(a)、ApoB/ApoA1水平与颈动脉粥样硬化的关系〔J〕.中国老年学杂志,2015;35(3):792-4.
11 Sultana A,Cochran BJ,Tabet F,etal.Inhibition of inflammatory signaling pathways in 3T3-L1 adipocytes by apolipoprotein A-I〔J〕.FASEB J,2016;30(6):2324-35.
12 袁 丁,梁 华,刘宏霞,等.胰岛素及格列齐特治疗2型糖尿病大鼠肝脏脂质沉积的机制探讨〔J〕.中国病理生理杂志,2014;30(6):1070-5.
13 Vieira E,Marroquí L,Batista TM,etal.The clock gene Rev-erbα regulates pancreatic β-cell function:modulation by leptin and high-fat diet〔J〕.Endocrinology,2012;153(2):592-601.
14 Gathercole L,Gareth G,Stuart A.11β-Hydroxysteroid Dehydrogenase1:translational and Therapeutic Aspects〔J〕.Endocrine Rev,2013;34(4):525-55.
15 Berger E,Héraud S,Mojallal A,etal.Pathways commonly dysregulated in mouse and human obese adipose tissue:FAT/CD36 modulates differentiation and lipogenesis〔J〕.Adipocyte,2015;4(3):161-80.
16 Huang GM,Jiang QH,Cai C,etal.SCD1 negatively regulates autophagy-induced cell death in human hepatocellular carcinoma through inactivation of the AMPK signaling pathway〔J〕.Cancer Lett,2015;358(2):180-90.
17 Ratner C,Madsen AN,Kristensen LV,etal.Impaired oxidative capacity due to decreased CPT1b levels as a contributing factor to fat accumulation in obesity〔J〕.Am J Physiol Regul Integr Comp Physiol,2015;308(11):973-82.
〔2015-11-13修回〕
(编辑 徐 杰)
武汉市卫生局科技课题(WX13C16)
林 梅(1970-),女,博士,主要从事糖尿病、甲状腺疾病、骨质疏松及肥胖等研究。
吴国琼(1975-),女,在读硕士,主治医师,主要从事内分泌疾病研究。
R589.2
A
1005-9202(2016)22-5527-03;
10.3969/j.issn.1005-9202.2016.22.015
猜你喜欢
体育科技文献通报(2022年3期)2022-05-23
学习与科普(2019年6期)2019-09-10
中成药(2018年9期)2018-10-09
天然产物研究与开发(2018年2期)2018-04-04
中成药(2018年1期)2018-02-02
中成药(2017年4期)2017-05-17
中国运动医学杂志(2016年3期)2016-07-10
中国运动医学杂志(2016年3期)2016-07-10
中国卫生标准管理(2015年24期)2016-01-14
中国当代医药(2015年21期)2015-03-01
