
毕赤酵母表达罗氏耶尔森氏菌(Yersinia rohdei)植酸酶和酶学特性研究
2017-01-12 08:13韩红娟孙百慧彭日荷姚泉洪
上海农业学报 2016年6期
陈 磊,许 晶,韩红娟,邱 瑾,孙百慧,彭日荷,姚泉洪
(1上海海洋大学食品学院,上海 201306;2上海市农业科学院生物技术研究所,上海 201106)
毕赤酵母表达罗氏耶尔森氏菌(Yersinia rohdei)植酸酶和酶学特性研究
陈 磊1,2,许 晶2,韩红娟2,邱 瑾1,2,孙百慧1,2,彭日荷2,姚泉洪2
(1上海海洋大学食品学院,上海 201306;2上海市农业科学院生物技术研究所,上海 201106)
按毕赤酵母偏爱密码子,将来自Yersinia rohdei的植酸酶基因(YrAPPA)进行了化学合成,并成功在毕赤酵母GS-115中异源分泌表达,重组植酸酶分泌量能达到72 μg/mL。酶学特性研究结果表明:酶比活达到1 500 U/mg,重组植酸酶的最适反应温度和pH分别为55℃和4.5;纯化的重组植酸酶能耐受60℃高温,且pH 3—7.5范围内重组植酸酶有良好的稳定性;Cu2+、Zn2+、Fe3+对植酸酶活性有抑制作用;重组植酸酶有抗胃蛋白酶和胰蛋白酶水解特性。
植酸酶;酶学特性;毕赤酵母;电击转化;异源表达
植酸即肌醇六磷酸及其盐类,广泛存在于植物组织和相应的粮食产品中,占其总量的1%—3%,其中磷含量占植物总磷的60%—90%,是磷和肌醇的主要储存形式[1-2]。植酸盐也可与蛋白质形成难溶的配合物,植酸形式的磷不但不能被单胃动物所消化利用,而且还是一种抗营养因子,会抑制多种营养成分的吸收和利用[3-6]。
植酸酶(E.C.3.1.3.8.肌醇六磷酸水解酶)是催化植酸(六磷酸环己六醇)及植酸盐水解成肌醇与磷酸(或磷酸盐)的一类酶的总称,属于磷酸单酯水解酶,是组氨酸酸性磷酸酶家族的重要成员,它广泛存在于动物、植物和微生物中[7]。植酸酶可使植物性饲料中磷的利用率提高60%,粪便中磷排出量减少40%,在饲料中添加植酸酶不仅可提高植物性饲料中磷的利用并减少粪便中磷的排放[8-10],而且能够通过降低植酸盐的抗营养作用,增加动物对蛋白质和一些矿物质元素的吸收能力[11-12]。因此对提高畜牧生产效率及降低磷对环境的污染有重要意义。
近年来,植酸酶在饲料工业中应用更加广泛,但各种不同的商业化植酸酶均存在一定缺陷,仍然有一些难题需要攻关。有必要开展新的植酸酶基因资源挖掘和功能研究,并最终利用毕赤酵母作为生物反应器,进行大规模低成本生产[13-16]。本研究设计并化学合成了来自罗氏耶尔森氏菌的植酸酶基因,通过电击转化法导入毕赤酵母(GS-115)细胞,实现异源高水平分泌表达[17],并对表达的植酸酶进行相关酶学特性研究。
1 材料与方法
1.1 试验材料
1.1.1 菌株和载体
毕赤酵母GS-115(His-Mut+)菌株,大肠杆菌(DH5α),pPIC9K酵母表达载体购自Invitrogen公司;毕赤酵母GS-115细胞感受态,重组表达载体pYPX88(GenBank登录号:AY178045)均为本实验室准备。
1.1.2 生化酶和试剂
Taq DNA聚合酶,T4 DNA连接酶,限制性核酸内切酶购自日本Takara公司;内切糖苷酶购于New England Biolab公司;其他所用药品均为分析纯,购自上海生工生物工程有限公司。
1.1.3 主要仪器
电击仪购自Bio-Rad公司;多功能酶标仪购自TECAN infinite M200瑞士帝肯公司。
1.2 方法
1.2.1 YrAPPA基因合成及表达载体的构建
源自NCBI数据库的YrAPPA植酸酶基因序列(GenBank登录号:ABU98779),在不改变植酸酶氨基酸序列的前提下,按照毕赤酵母密码子偏爱优化:选择在毕赤酵母表达概率高的密码子替换表达概率低的密码子。YrAPPA基因序列优化后,根据PTDS法[18](PCR-based two-step DNA synthesis method,PTDS)进行化学合成,设计大量带有重叠序列的引物(表1),每个引物由60个寡核苷酸组成,相邻引物之间存在20 bp的互补序列,第一步通过连续重叠延伸PCR法合成500 bp左右的中间片段;第二步将第一步合成的全部中间片段等比例混合并用作模板,使用基因最外侧的一对引物再次进行PCR扩增获得全长基因(图1)。

表1 YrAPPA基因合成的引物Table 1 Primers of YrAPPA gene synthesis

(续表1)

图1 YrAPPA基因的PCR扩增合成策略Fig.1 Strategy of PCR amplification and synthesis of YrAPPA gene
使用1.5%的琼脂糖凝胶电泳分离产物,DNA胶回收试剂盒回收。回收产物克隆到pMD18-T载体上进行DNA测序,获得正确的带有植酸酶合成基因YrAPPA的重组质粒。重组质粒经BamH I和Sac I双酶切,与经过同样双酶切的pPIC9K载体连接,得到重组表达载体pYPX88-YrAPPA。
1.2.2 表达载体pYPX88-YrAPPA电击转化
挑取毕赤酵母单菌落接种于5 mL YPD液体培养基(酵母提取物10 g/L,胰化蛋白胨20 g/L,pH 5.5,用前加入2%的葡萄糖)30℃过夜摇培活化毕赤酵母菌。取1 mL过夜菌接种于50 mL YPD,30℃,200 g/min揺菌至OD600=1.6—1.8,然后根据Invitrogen手册的方法制备毕赤酵母感受态细胞。取2 μL Bgl II酶切后的pYPX88-YrAPPA载体和80 μL毕赤酵母感受态细胞,混匀后冰浴5 min,在电压1.75 kV,电阻400 Ω条件下电击转化,点击后立即加入800 μL预冷的0.6 mol/L山梨醇溶液。将混合液离心弃上清,用100 μL无菌水悬浮菌体,并涂布于SD-山梨醇平板(补充材料),置于30℃恒温培养箱中培养72 h。
1.2.3 阳性毕赤酵母菌株诱导表达
挑取阳性菌落接种于100 mL BMGY培养基(500 mL/L 2SD,10 g/L酵母提取物,20 g/L胰化蛋白胨,5 μL/L 4×106倍生物素,pH 5.8,用前添加1%甘油),30℃摇床连续培养约2 d,达到指数生长期(OD600约为5.0)时,离心收集菌体,加入适量无菌水清洗沉淀,清除残留的甘油。加入20 mL BMMY培养基(2SD培养基和蒸馏水1∶1混合,pH 5.8,用前添加1%甲醇)重新悬浮菌体,30℃连续诱导培养3 d,每隔24 h添加1%甲醇诱导分泌表达植酸酶。培养至第3天,将菌液加入50 mL离心管中,置于4℃离心机中12 000 r/min离心15 min,取上清液即粗酶液,4℃冰箱保存。
1.2.4 植酸酶分离纯化和SDS-PAGE分析
取1 mL粗酶液加入已经用平衡缓冲液(pH 7.4;20 mmol/L HEPES,10 mmol/L咪唑,100 mmol/L NaCl)清洗的Ni2+-NTA琼脂糖亲和柱中,粗酶液完全流经镍柱后,加入两次1 mL洗涤缓冲液(pH 7.4;20 mmol/L HEPES,30 mmol/L咪唑,500 mmol/L NaCl)清洗挂柱的杂蛋白,最后加入3次500 μL洗脱缓冲液(pH 7.4;20 mmol/L HEPES,250 mmol/L咪唑,500 mmol/L NaCl),洗脱挂柱的重组植酸酶蛋白。
通过去糖基化酶(Endo H)处理粗酶液:16 μL粗酶液和1.6 μL糖蛋白变性液混合后100℃变性10 min,再加入2 μL G5缓冲液和1 μL内切糖苷酶H混匀,在37℃温育2 h;将纯化后的粗酶液和去糖基化后的酶液采用SDS-PAGE(12%分离胶和5%积层胶)进行鉴定,并使用Bradford法测定酶液蛋白含量。
1.3 酶活分析
根据钒-钼黄分光光度法[19],5 μL的粗酶液和100 μL的5 mmol/L植酸钠(溶于0.25 mol/L的乙酸缓冲液)混合后,在37℃反应30 min后加入100 μL的显色液(100 g/L的钼酸铵溶液和2.35 g/L的偏钒酸铵溶液按1∶1混合使用)和200 μL硝酸溶液(硝酸和水1∶2)终止反应。在415 nm波长下测定吸光值,每个数据有3个重复。每分钟内从浓度为5.0 mmol/L植酸钠溶液中释放1 μmol无机磷,即为一个植酸酶活性单位(1 U)。
配置50 mmol/L的基准磷酸二氢钾(称取0.6804 g磷酸二氢钾,用pH 5.5的乙酸缓冲液定容到100 mL),再分别用蒸馏水将其稀释为1.5625 μmol/mL、3.1250 μmol/mL、6.2500 μmol/mL、12.500 μmol/mL、25.000 μmol/mL五个浓度梯度。每个浓度梯度分别做1个空白对照(用蒸馏水代替磷酸二氢钾)和3组重复试验,在标准差范围内取平均值表示。
1.3.1 最适温度和热稳定性
用0.2 mol/L NaAc-HAc缓冲液(pH 4.0)稀释好的粗酶液,与植酸钠混合(反应体系如1.3)后在不同温度下(10—85℃,每间隔5℃)反应30 min,根据钒-钼黄分光光度法后测OD415计算酶的相对酶活,以酶最高相对活性所对应的温度为最适温度。
分别在3个不同温度(60℃,65℃和70℃)下温育粗酶液,测定重组植酸酶的热稳定性。每隔5 min(0—30 min)取出酶液,在最适条件下(55℃,pH 4.5)反应30 min后根据钒-钼黄分光光度法测得OD415计算酶的相对酶活。以上均利用变性失活的粗酶液作为空白对照,每组数据均有3个重复数据。
1.3.2 最适pH值和pH稳定性
将植酸钠分别溶解在不同pH值缓冲液中:0.2 mol/L Gly-HCl(pH 1.0—3.5),0.2 mol/L NaAc-HAc(pH 4.0—6.5),0.2 mol/L Tris-HCl(pH 7.0—8.5)和Gly-NaOH(pH 9.0—10.0),在55℃下反应30 min后,根据钒-钼黄分光光度法测OD415计算酶的相对酶活,以酶最高相对活性所对应的pH值为最适pH。
pH稳定性研究是将粗酶液加入不同缓冲液(同上)中,37℃处理30 h后,在最适条件下(55℃,pH 4.5)反应30 min,根据钒-钼黄分光光度法测得OD415,并计算酶的相对酶活。以变性失活的粗酶液作为空白对照,每组数据均有3个重复数据。
1.3.3 不同金属离子对酶活力的影响
0.2 mol/L NaAc-HAc(pH 4.5)缓冲液分别溶解不同金属离子溶液[KNO3、NaCl、Ca(NO3)2、(NH4)2SO4、MgSO4、ZnSO4、AlCl3、NiCl2、CoCl2、FeCl3、LiCl、MnCl2、CrCl3、CuSO4、BaCl2],离子最终浓度为2 mmol/L。在37℃温育30 min后在最适条件下(55℃,pH 4.5)反应30 min,根据钒-钼黄分光光度法测得重组植酸酶的剩余酶活性,以未加入金属离子处理的重组植酸酶活为空白对照。
1.3.4 蛋白酶对重组植酸酶酶活的影响
用0.2 mol/L Gly-HCl(pH 2.0)和0.2 mol/L Tris-HCl(pH 7.5)缓冲液分别稀释好胃蛋白酶和胰蛋白酶,将稀释好的消化蛋白酶液与植酸酶按一定重量比例[蛋白酶/植酸酶(w/w):500/1、400/1、200/1、100/1、50/1、1/1、1/50、1/100、1/200、1/400、1/500]混合。在37℃处理5 h后,最适条件下(55℃,pH 4.5)反应30 min,根据钒-钼黄分光光度法测得不同质量比消化酶液处理下重组植酸酶的剩余酶活,以未经过消化酶液处理的重组植酸酶酶活作空白对照。
2 结果与分析
2.1 YrAPPA基因合成
为了实现YrAPPA基因在毕赤酵母中高效分泌表达,本研究根据毕赤酵母偏爱密码子对基因进行了化学合成和结构优化,在基因3’端引入了编码6×His的核苷酸序列,便于表达后蛋白的分离纯化;消除了野生型基因中编码N端23个氨基酸的信号肽基因序列(如图2所示下划线序列);分别在基因5’和3’末端添加BamH I和Sac I限制性酶切位点(如图2中方框序列所示);合成基因与野生型基因的核苷酸序列同源性为71.17%(图2)。

图2 YrAPPA合成基因(YrAPPA-OP)和野生型基因(YrAPPA-WT)序列比对Fig.2 Sequence alignment of synthetic gene(YrAPPA-OP)and wild gene(YrAPPA-WT)

图3 SDS-PAGE检测植酸酶Fig.3 Detection of phytase from P.pastoris by SDS-PAGE
2.2 SDS-PAGE分析
在经过72 h诱导培养后,重组植酸酶分泌表达量达到了72 μg/mL,根据酶比活定义计算得到重组植酸酶的比活为1 500 U/mg。经SDS-PAGE分析,图3中粗酶液泳道中蛋白条带单一且没有杂蛋白条带的出现,重组植酸酶经过内切糖苷酶H去糖基化后与未去糖基化的分子量均为47 kDa,而且与生物信息学预测的分子量理论值相当,这些结果都表明重组植酸酶并没有被糖基化修饰。
2.3 最适温度和热稳定性
如图4所示:重组植酸酶的最适温度在55℃,而且在25—60℃范围内酶活比较稳定,相对酶活均在50%以上;图5所示酶的热稳定性研究,设定酶液在55℃下的相对酶活为100%。结果显示,置于60℃温育10—30 min,植酸酶的相对酶活都比较稳定,均在40%左右,而在65℃和70℃下,相对酶活仅20%左右;因此60℃条件下植酸酶的热稳定性优于65℃和70℃。

图4 植酸酶的最适温度Fig.4 Optimum temperature for phytase

图5 植酸酶的热稳定性Fig.5 Thermal stability of phytase
2.4 最适pH和pH稳定性
通过在不同的pH条件下测定酶的相对酶活,图6结果显示,植酸酶的最适pH为4.5,在pH为2—6的范围内,植酸酶相对酶活均在50%以上,说明酶在酸性条件下有很好的活性;在pH 3—7.5范围内相对酶活均在60%以上。以上结果说明植酸酶的pH适宜范围比较宽,在偏酸和弱碱性区域有很好的适应性。
2.5 金属离子对重组植酸酶活性的影响
通过2 mmol/L的不同金属离子处理,0.5 h后分析对植酸酶酶活的影响。图7结果显示,Cu2+、Fe3+和Zn2+对重组植酸酶有很强的抑制作用,剩余相对酶活分别为14.77%、35.59%和38.04%,对重组植酸酶的抑制均超过50%,尤其是Cu2+的抑制作用最大。其他的如Ni2+、Mg2+、Cr3+、Co2+等其他离子对酶的抑制仅有百分之十几,对酶活影响较小。

图6 植酸酶的最适pH值和pH稳定性Fig.6 Optimum pH and pH stability of phytase
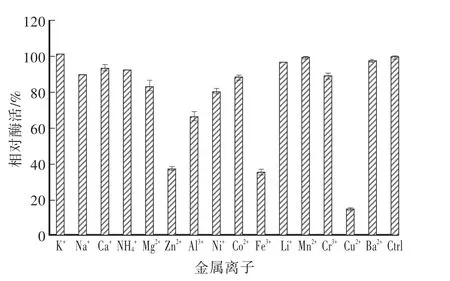
图7 金属离子对植酸酶的影响Fig.7 Effects of metal ions on phytase activity
2.6 蛋白酶对YrAPPA植酸酶活性的影响
图8显示,胃蛋白酶液处理重组植酸酶5 h后,在w/w为1/500—400/1,重组植酸酶的剩余酶活大于65%,即使在抑制作用最强(w/w:500/1)的情况下,重组植酸酶酶活还剩余53.72%。但是胰蛋白酶液处理重组植酸酶5 h后,胰酶/植酸酶(w/w)在1/400—1/1间植酸酶的剩余活性均在100%以上,在质量比为50/1时植酸酶的剩余酶活降为100%以下;这说明了重组植酸酶对于胰蛋白酶和胃蛋白酶均具有有良好的抗水解的能力。
2.7 重组植酸酶反应动力学参数
通过双倒数作图法:以反应速率的倒数为纵坐标,以底物浓度的倒数为横坐标作图,如图9所示,根据回归方程计算得到米氏常数Km=0.37 μmol/mL,最大反应速率Vm=0.89 μmol/(mL·min)。
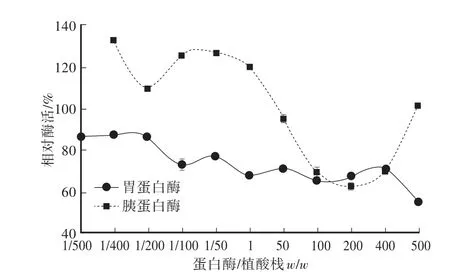
图8 胃蛋白酶和胰蛋白酶对植酸酶的影响Fig.8 Effects of pepsin and trypsin on phytase activity

图9 双倒数图Fig.9 The Lineweaver-Burk plot
3 讨论
本研究对源自罗氏耶尔森氏菌的植酸酶YrAPPA基因进行了优化,包括:在不改变氨基酸序列的前提下,按宿主细胞的偏爱密码子进行优化,提高基因翻译效率;消除基因内部的常用限制性内切酶识别位点,便于载体构建;消除逆向重复序列、茎环结构和转录终止信号,使基因内部的GC/AT均衡,提高RNA的稳定性;优化mRNA二级结构自由能,以提高基因表达效率。在完成基因设计的基础上,用本实验室建立的PTDS法对YrAPPA基因进行了化学合成,并且成功在毕赤酵母中实现了高水平的异源表达。PTDS法操作简便,不需要DNA模板,引物也不需要磷酸化,通过一系列带有互补序列的引物合成外源基因。作为真核表达系统,毕赤酵母具有较少外分泌蛋白干扰、高水平表达和蛋白翻译后修饰等优点。本研究中,优化后的YrAPPA基因在毕赤酵母中获得高水平分泌表达,且重组植酸酶具有很高的比活力(1 500 U/mg)。如果将该重组植酸酶应用于大规模工业化生产,有望大幅度降低生产成本。
YrAPPA植酸酶的活性受很多金属离子的调节,如Cu2+、Fe3+和Zn2+金属离子抑制植酸酶活性60%以上。而Ni2+、Mg2+、Cr3+等离子对重组植酸酶的酶活只有百分十几的抑制作用,这与植酸可与多价的金属离子形成难溶的螯合物[12],降低了底物的有效浓度从而使相对酶活有些下降有关。
用胃蛋白酶(w/w:500/1)液处理5 h后,植酸酶剩余酶活为53.72%;但是经1/400—1/1(w/w)胰蛋白酶液处理5 h后,重组植酸酶的酶活有所提升,表现出抗胰化蛋白酶水解的特点。在其他的植酸酶研究中也发现经过胰蛋白酶处理后植酸酶的酶活会有所增加[20-22],推测可能是被胰蛋白酶水解生成的植酸酶小片段中有活性更高的抗胰蛋白酶多肽存在,从而增加了植酸酶的活性。
在目前商业化应用中,对于重组植酸酶作为动物饲料添加剂主要存在三个关键性的难题,包括:胃液中酸性环境对酶活性的抑制,热稳定性差影响植酸酶造粒以及动物肠胃中胃蛋白酶对于植酸酶的水解而降低活性[23-24]。而本研究所得到的重组植酸酶有明显的优势:首先,重组植酸酶有比较好的耐热性,在60℃温育30 min后还有40%以上的剩余酶活,能够较好地解决植酸酶在饲料生产过程中因高温制粒导致剩余酶活偏低的问题;其次,该酶有比较稳定和广泛的pH范围,在pH 3—7.5温育30 h后重组酶还剩余60%以上酶活,说明该酶在消化道内比较复杂的酸度环境下有比较好的耐受能力;对重组植酸酶的抗消化酶能力的研究发现,该酶具有良好的抗胃蛋白酶和胰蛋白酶水解能力,尤其是抗胃蛋白酶的能力很强,胃蛋白酶在1/100—400/1(w/w)之间重组植酸酶的剩余酶活比较稳定,即使在抑制作用最强(w/w:500/1)时,重组植酸酶还剩余50%以上酶活。本研究所得到的YrAPPA植酸酶的相关酶学特性的数据,尤其是蛋白酶对于酶活的影响,为实际应用和后续研究提供了重要的参考。
[1]杨平平,王燕,史宝军,等.植酸酶酶活测定方法的研究[J].饲料工业,2003,24(8):34-36.
[2]彭日荷,熊爱生,李贤,等.应用毕氏酵母高效表达耐高温植酸酶[J].生物化学与生物物理学报,2002,1(6):725-730.
[3]柯晓静.黑曲霉WP1植酸酶基因的克隆及在毕赤酵母中的表达[D].保定:河北农业大学,2007.
[4]付大伟.Yersinia spp.来源植酸酶的酶学性质及结构与功能研究[D].北京:中国农业科学院,2010.
[5]李晓龙,杨合同,扈进冬,等.植酸酶的多样性及其分类[J].微生物学通报,2010,37(5):738-747.
[6]张民扬,李吕木,唐倩,等.植酸酶在畜禽生产中的应用研究进展[J].添加剂世界,2015(10):44-48.
[7]赵冬敏.植酸酶基因phyA的克隆及在毕赤酵母中的高效表达[D].泰安:山东农业大学,2007.
[8]NELSON T S,SHIEL T R,WODZINSKI R J,et al.The availability of phytase phosphorusin soybean meal before and after treatment with a mold phytase[J].Poult Sci,1968,47(6):1842-1848.
[9]蒋守群,吴天星.植酸酶的研究进展[J].动物营养学报,1999,11(3):1-11.
[10]NELSON T S.The utilization of phytate phosphorus by poultry:A review[J].Poultry Science,1967,46(4):862-871.
[11]WYSS M,PASAMONTES L,FRIEDLEIN A,et al.Biophysical characterization of fungal phytases(myo-inositol hexakisphosphate phosphydrolases):Molecular size,glycosylation pattern,and engineering of proteolytic resistance[J].Appl Environ Microbio,1999,65(2):359-366.
[12]贺建华.植酸磷和植酸酶研究进展[J].动物营养学报,2005,17(1):1-6.
[13]ZHAO W,XIONG A S,FU X Y,et al.High level expression of an acid-stable phytase from Citrobacter freundii in Pichia pastoris[J].Appl Biochem Biotechnol,2010,162(8):2157-2165.
[14]HUANG H Q,LUO H Y,WANG Y R,et al.A novel phytase from Yersinia rohdei with high phytate hydrolysis activity under low pH and strong pepsin conditions[J].Appl Microbiol Biotechnol,2008,80(3):417-426.
[15]SHIVANGE A V,ANNEGRET S,ALEXANDER D,et al.Directed evolution of a highly activeYersinia mollaretii phytase[J].Appl Microbiol Biotechnol,2012,95(2):405-418.
[16]姚斌,张春义,王建华,等.高效表达具有生物学活性的植酸酶的毕赤酵母[J].中国科学,1998,1(3):237-243.
[17]姚斌,张春义,王建华,等.产植酸酶的黑曲霉菌株筛选及其植酸酶基因克隆[J].农业生物技术学报,1998,1(1):1-6.
[18]XIONG A S,YAO Q H,PENG R H,et al.A simple,rapid,high-fidelity and cost-effective PCR-based two-step DNA synthesis method for long gene sequences[J].Nucleic Acids Research,2003,32(12):437-442.
[19]蒋建军.植酸含量的测定:钒钼黄比色法的方法探讨[J].陕西化工,1994(4):38-40.
[20]李平,易弋,石征宇,等.嗜热地衣芽孢杆菌植酸酶的原核表达及酶学性质研究[J].食品工业科技,2015,37(8):1-7.
[21]ZHU W F,QIAO D Q,HUANG M,et al.Modifying thermostability of appA from Escherichia coli[J].Curr Microbiol,2010,61(4):267-273.
[22]RODRIGUEZ E,PORRES J M,HAN Y M,et al.Different sensitivity of recombinant Aspergillus niger phytase(r-PhyA)and Escherichia coli pH 2.5 acid phosphatase(r-AppA)to trypsin and pepsin in vitro[J].Archives of Biochemistry and Biophysics,1999,365(2):262-267.
[23]马玺.植物植酸酶应用特性的研究[D].哈尔滨:东北农业大学,2001.
[24]王祖鹏,许伟,邵荣,等.基于定点突变改善中性植酸酶催化特性及结构效应分析[J].食品科学,2015,36(19):118-123.
(责任编辑:程智强)
Expression of phytase from Yersinia rohdei in Pichia pastoris and its enzymatic characteristics
CHEN Lei1,2,XU Jing2,HAN Hong-juan2,QIU Jin1,2,SUN Bai-hui1,2,PENG Ri-he2,YAO Quan-hong2*
(1College of Food Science and Technology,Shanghai Ocean University,Shanghai 201306,China;2Biotechnology Research Institute,Shanghai Academy of Agricultural Sciences,Shanghai 201106,China)
Yersinia rohdei phytase gene(YrAPPA)was chemically synthesized according to the codon usage of Pichia pastoris and successfully expressed in P.pastoris GS-115,with the secretion yield of recombinant phytase being up to about 72 μg/mL.The enzymatic characterization showed that the specific activity of enzyme is 1 500 U/mg,and the optimal reacting temperature and pH of recombinant phytase are respectively 55℃and 4.5;The purified recombinant phytase can tolerate high temperature at 60℃and has a good stability within pH 3—7.5;The phytase whose activity is inhibited by Cu2+,Zn2+and Fe3+has characteristics resistant to trypsin and pepsin.
Phytase;Enzymatic property;Pichia pastosis;Electroporation;Heterologous expression
Q78
A
1000-3924(2016)06-018-08
2015-12-07
上海市科委农业成果转化项目(143919NO300);上海市种业发展项目[沪农科种字(2016)第1-2号]
陈磊(1990—),男,在读硕士,研究方向:生物化学与分子生物学。Tel:13262988682,E-mail:chenleihefei@163.com
*通信作者:姚泉洪(1965—),男,研究员,研究方向:植物和微生物基因工程。E-mail:yao.quanhong65@yahoo.com
猜你喜欢
中国饲料(2022年13期)2022-07-07
食品与发酵工业(2022年6期)2022-04-01
中国畜牧杂志(2020年11期)2020-11-18
国外畜牧学(猪与禽)(2020年8期)2020-03-04
生物技术通报(2019年7期)2019-02-21
中国畜禽种业(2018年1期)2018-01-18
广东饲料(2016年1期)2016-12-01
工业微生物(2016年5期)2016-11-11
化学与生物工程(2016年10期)2016-11-10
畜牧兽医科技信息(2015年5期)2015-12-27
