
不同生态因子对岩扇贝幼贝耗氧率和排氨率的影响
2017-06-19 18:56曹善茂梁伟锋刘钢李晓雨大连海洋大学辽宁省贝类良种繁育工程技术研究中心辽宁大连116023
大连海洋大学学报 2017年3期
曹善茂,梁伟锋,刘钢,李晓雨(大连海洋大学辽宁省贝类良种繁育工程技术研究中心,辽宁大连116023)
不同生态因子对岩扇贝幼贝耗氧率和排氨率的影响
曹善茂,梁伟锋,刘钢,李晓雨
(大连海洋大学辽宁省贝类良种繁育工程技术研究中心,辽宁大连116023)
为研究岩扇贝Crassadoma gigante的代谢生理状况,为其人工育苗和规模化养殖提供依据,采用不同生态条件对岩扇贝幼贝耗氧率和排氨率进行了相关研究。结果表明:温度、盐度、pH和体质量对岩扇贝幼贝耗氧率、排氨率均有极显著影响(P<0.01);在试验温度为6~22益时,岩扇贝幼贝单位体质量的耗氧率、排氨率随温度的升高而增大;经回归分析,体质量(X)与耗氧率或排氨率(Y)均呈负相关幂指数关系,岩扇贝幼贝单位体质量的耗氧率、排氨率随体质量的增大而降低;岩扇贝幼贝的耗氧率与排氨率先随盐度(24~30)的升高而增大,在盐度为30时达到最大值,而后随盐度(30~36)的升高而降低;岩扇贝幼贝的耗氧率与排氨率先随pH(7.0~8.0)的升高而增大,在盐度为8.0时达到最大值,而后随pH(8.0~9.0)的升高而降低;岩扇贝幼贝一昼夜的耗氧率及排氨率最高峰均在13:00,耗氧率和排氨率最低峰均在1:00。本研究结果可为岩扇贝的生理生态学研究提供理论依据。
岩扇贝幼贝;耗氧率;排氨率;昼夜节律
岩扇贝Crassadoma gigantea隶属于软体动物门Mollusca、瓣鳃纲Lamellibranchia、珍珠贝目Pteri-oida、扇贝科Pectinidae。该扇贝广泛分布于北美太平洋沿岸,具有个体大、闭壳肌大、生长快、味道鲜美等特点,是优良的扇贝品种,当前因过度采捕致其资源量不断下降,因而在国外市场供不应求[1]。目前,国外学者对岩扇贝的营养成分[1]、口味[1]、冷藏肉质的稳定性[2]、野生种群的生长速率比较[3]、排精产卵机制[4]、幼贝的饵料配比[5]、附着生理[6]等方面进行了研究,而国内学者对岩扇贝的研究较少,自2013年引进中国以来,主要进行了其人工育苗技术[7]、闭壳肌营养成分[8]和幼贝摄食生理的研究[9],尚未见对其代谢生理的相关研究报道。贝类的代谢生理是研究其生物学、生理生态学的重要内容之一,研究贝类耗氧率、排氨率对掌握其呼吸与排泄的特点,了解其生理状态、最适生存条件、对环境的适应情况,以及其生活习性、增养殖等具有重要的指导意义。本试验中,研究了不同体质量的岩扇贝幼贝在不同温度、盐度、pH条件下,以及昼夜不同时段的耗氧率和排氨率,以期为岩扇贝的人工育苗及规模化养殖提供理论依据。
1 材料与方法
1.1 材料
试验用贝为从加拿大温哥华岛引进的岩扇贝亲贝,经人工育苗后获得的子一代幼贝,取720枚待用。试验用海水均为大连市黑石礁海域沙滤自然海水。
1.2 方法
挑选状态良好、无损伤的个体,洗刷去除体表附着物,暂养于控温循环水族箱中,水温为(14.0依0.3)益,试验期间定时投饵换水。试验用饵料有新月菱形藻Nitzschia closterium、湛江等鞭金藻Isochrysis zhanjiangensis、扁藻Platymonus spp.、小球藻Chlorella spp.,均为实验室培养所得的新鲜藻液。日投饵量逸25伊104cells/mL(以新月菱形藻计),每两天换水1/3~1/2,主投新月菱形藻,辅投湛江等鞭金藻、扁藻、小球藻,每天测定水化指标。暂养期间,海水盐度为31.6~32.3,pH为7.93~8.33,溶解氧>5 mg/L。
1.2.1 试验设计岩扇贝幼贝暂养20 d后开始试验,挑选体质量差异较小的幼贝进行温度、盐度、pH和昼夜节律试验,体质量试验挑选小、中、大3种规格的幼贝。试验开始时间均为9:00。
温度试验设6、10、14、18、22益共5个温度组,从14益开始,每日升高或降低1益,试验盐度为31.6,pH为8.06。
盐度试验设24、27、30、33、36共5个盐度组,从31.6开始,每日升高或降低1.4~3.0。盐度用海水晶和无氨纯水调节,试验温度为18益, pH为8.06。
pH试验设7.0、7.5、8.0、8.5、9.0共5个pH组,从8.06开始,每日升高或降低0.06~ 0.50。pH用1 mol/L盐酸和氢氧化钠调节,试验温度为18益,盐度为31.6。
昼夜节律试验设9:00~13:00、13:00~ 17:00、17:00~21:00、21:00~1:00、1:00~5:00、5:00~9:00共6个时间段。试验时对试验装置进行遮光处理以保持全程黑暗,试验温度为18益,盐度为31.6,pH为8.06。从9:00开始试验,连续测定13:00、17:00、21:00、1:00、5:00、9:00共6个时间点的耗氧率与排氨率。
试验中用橡皮胶管通过虹吸的方法从同一个水槽往呼吸瓶中加水,连续不间断注入约3倍呼吸瓶体积的水,最后注满瓶子并消除附着在瓶壁的气泡,放入试验贝后封口。上一时间段试验结束后,即时测定呼吸瓶中水样的耗氧率与排氨率,并将试验贝移入预先准备好的下一组呼吸瓶中继续下一时段试验。
以上试验在达到试验条件后,稳定48 h,试验前停止投饵24 h,以避免其排便对试验结果产生影响。以500 mL锥形瓶为呼吸瓶,每瓶放5个幼贝,空白对照组不放贝,以平整玻璃片封口,试验时间2 h,试验期间杜绝任何惊扰。试验结束时,匀呼吸瓶使水样溶解氧、氨氮均匀分布,测定试验前后水样的溶解氧(DO)和氨氮(NH+4-N)含量。每组设3个平行,另设1个空白对照。试验结束后,用游标卡尺测量试验贝的壳长、壳高,然后解剖分离贝壳与软体部,均置于烘箱(70益)中烘干至恒重。试验用岩扇贝幼贝规格参数见表1。
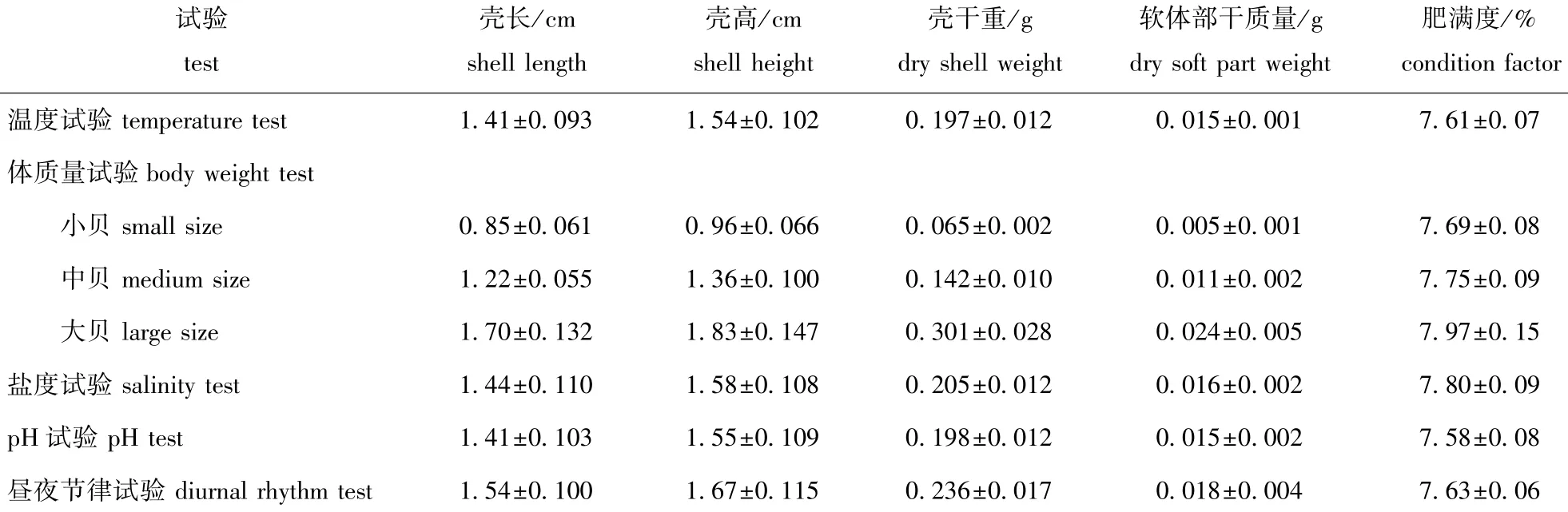
表1 试验用岩扇贝幼贝规格Tab.1Specifications of juvenile rock scallop Crassadoma gigantea used in the experiment
1.2.2 指标的测定与计算采用Winkler碘量法测定溶解氧含量,采用次溴酸钠氧化法测定氨氮含量。
耗氧率与排氨率的计算公式为

其中:OR为单位体质量的耗氧率[mg/(g· h)];DO0和DOt分别为平行组试验开始和结束时水中的DO含量(mg/L);DO0忆和DOt忆分别为对照组试验开始和结束时水中的DO含量(mg/L); NR为单位体质量的排氨率[mg/(g·h)];N0和Nt分别为平行组试验开始和结束时水中的NH+4-N含量(mg/L);N0忆和Nt忆分别为对照组试验开始和结束时水中的NH+4-N含量(mg/L);V为呼吸瓶中水的体积(L),本试验均为0.59 L;W为试验贝软体部干质量(g);t为试验时间(h),除昼夜节律试验外均为2 h;Q10为温度对贝类代谢的影响强度;M1和M2分别为t1和t2温度下贝类的代谢率。
1.3 数据处理
采用SPSS 19.0软件对耗氧率、排氨率进行单因子方差分析(One-way ANOVA)及组间多重比较分析(LSD),极显著性水平设为0.01,显著性水平设为0.05。
2 结果与分析
2.1 不同温度下岩扇贝幼贝耗氧率和排氨率
从图1可知,岩扇贝幼贝耗氧率、排氨率与温度的变化呈正相关关系。在18益时上升幅度突然变大,在18益后上升幅度有所减缓,而排氨率仍大幅升高。方差分析和组间多重比较分析表明,温度对岩扇贝幼贝耗氧率、排氨率均有显著影响(P<0.01),各温度组间均有显著性差异(P< 0.05)。
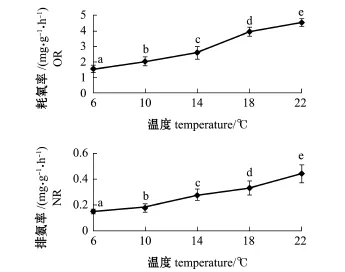
图1 温度对岩扇贝幼贝耗氧率和排氨率的影响Fig.1Effects of temperature on oxygen consumption rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea
从表2可见,岩扇贝幼贝的耗氧率Q10值为1.40~2.76,在14~18益时Q10值最高。

表2 水温变化对岩扇贝幼贝代谢率的影响Tab.2Influence of tem perature on metabolism rate of ju-venile rock scallop Crassadoma gigantea
2.2 不同体质量岩扇贝幼贝的耗氧率和排氨率
在温度为18益、盐度为31.6、pH为8.06的条件下,体质量对岩扇贝幼贝耗氧率、排氨率的影响见图2。从图2可知,岩扇贝幼贝单位体质量的耗氧率、排氨率均随软体部干质量的增大而降低。经回归分析表明,耗氧率(Y1)、排氨率(Y2)与软体部干质量(X1、X2)间呈负相关幂函数关系,回归方程分别为
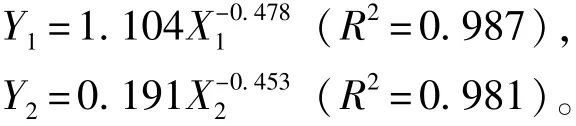
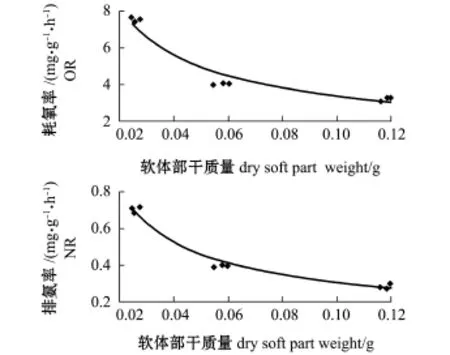
图2 体质量对岩扇贝幼贝耗氧率和排氨率的影响Fig.2Effect of body weight on oxygen consumption rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea
2.3 不同盐度下岩扇贝幼贝的耗氧率和排氨率
从图3可知,盐度为24~36时,岩扇贝幼贝的耗氧率、排氨率均随盐度的增大呈先增大后减小的趋势,均在盐度为30时达到最大值。方差分析表明,盐度对岩扇贝幼贝的耗氧率、排氨率均有极显著影响(P<0.01)。组间多重比较分析表明:盐度30和33组间的耗氧率无显著性差异(P> 0.05),其余盐度间的耗氧率均有显著性差异(P< 0.05);盐度24和36组间的排氨率无显著性差异(P>0.05),其余盐度间的排氨率均有显著性差异(P<0.05)。
2.4 不同pH下岩扇贝幼贝的耗氧率和排氨率
从图4可知,pH为7.0~9.0时,岩扇贝幼贝的耗氧率、排氨率均随pH的增大而呈先增大后减小的趋势,均在pH为8.0时达到最大值。方差分析表明,pH对岩扇贝幼贝耗氧率、排氨率有极显著影响(P<0.01)。组间多重比较分析表明,pH为7.5、8.5、9.0时,3组间的耗氧率无显著性差异(P>0.05),其余pH组间的耗氧率均有显著性差异(P<0.05);pH为7.5和9.0时,两组间的排氨率无显著性差异(P>0.05),其余pH组间的排氨率均有显著性差异(P<0.05)。
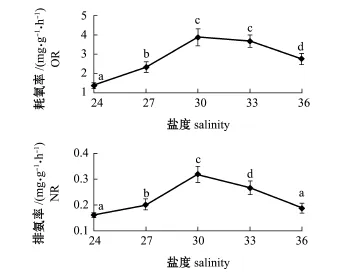
图3 盐度对岩扇贝幼贝耗氧率和排氨率的影响Fig.3Effects of salinity on oxygen consum p tion rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea
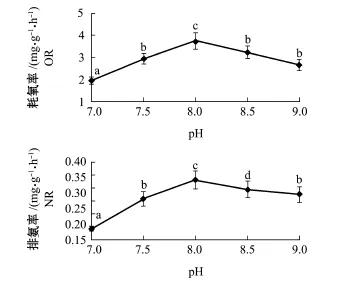
图4pH对岩扇贝幼贝耗氧率和排氨率的影响Fig.4Effect of pH on oxygen consumption rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea
2.5 岩扇贝幼贝耗氧率和排氨率的昼夜变化
从图5可见,岩扇贝幼贝耗氧率有两个高峰期,分别为9:00~13:00、1:00~5:00,耗氧率分别为5.209、3.895 mg/(g·h),在24 h内基本呈现出“高—低—高冶昼夜变化规律。岩扇贝幼贝的排氨率也有两个高峰期,分别为9:00~ 13:00和1:00~5:00,排氨率分别达到0.648、0.417 mg/(g·h),在24 h内排氨率变化规律也呈现“高—低—高冶的昼夜变化规律。
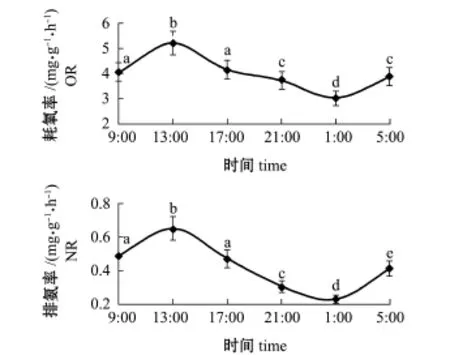
图5 岩扇贝幼贝耗氧率和排氨率的昼夜变化Fig.5Diurnal variation in oxygen consumption rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea
2.6 岩扇贝幼贝在不同条件下的氧氮比
如表3所示,当温度为6~22益时,岩扇贝幼贝O/N值为9.45~11.77,说明岩扇贝幼贝主要是蛋白质供能,其次是脂肪和碳水化合物。在温度为18益的自然海水中,无论何种规格的岩扇贝幼贝的代谢物结构均一致,均以蛋白质供能为主,脂肪和碳水化合物次之。不同规格的岩扇贝幼贝其O/N值为10.20~11.27,表明幼贝在此阶段主要以蛋白质供能,脂肪和碳水化合物供能占比例较低。当盐度为24~36时,岩扇贝幼贝的O/N值为8.76~ 14.70,且随盐度增大而增大,说明盐度升高会使岩扇贝幼贝蛋白质的代谢水平下降,蛋白质供能比重降低,脂肪和碳水化合物供能比重增加。当pH为7.0~9.0时,岩扇贝幼贝的O/N值为8.92~ 10.92,表明在适宜pH范围内,岩扇贝幼贝蛋白质代谢水平较高,但随着pH的进一步降低或升高,O/N值减小,蛋白质代谢水平降低。试验期间,岩扇贝幼贝的O/N值有明显的昼夜变化,最小值(8.03)在9:00~13:00,最大值(12.99)在21:00~1:00,而一昼夜中,9:00~13:00是岩扇贝幼贝耗氧率与排氨率的最高峰,21:00~ 1:00是耗氧率与排氨率的最低峰,由此可见,当岩扇贝幼贝代谢水平升高时,蛋白质供能比重增加,脂肪和碳水化合物比重降低;而代谢水平降低时,脂肪和碳水化合物供能比重增加,蛋白质比重降低。
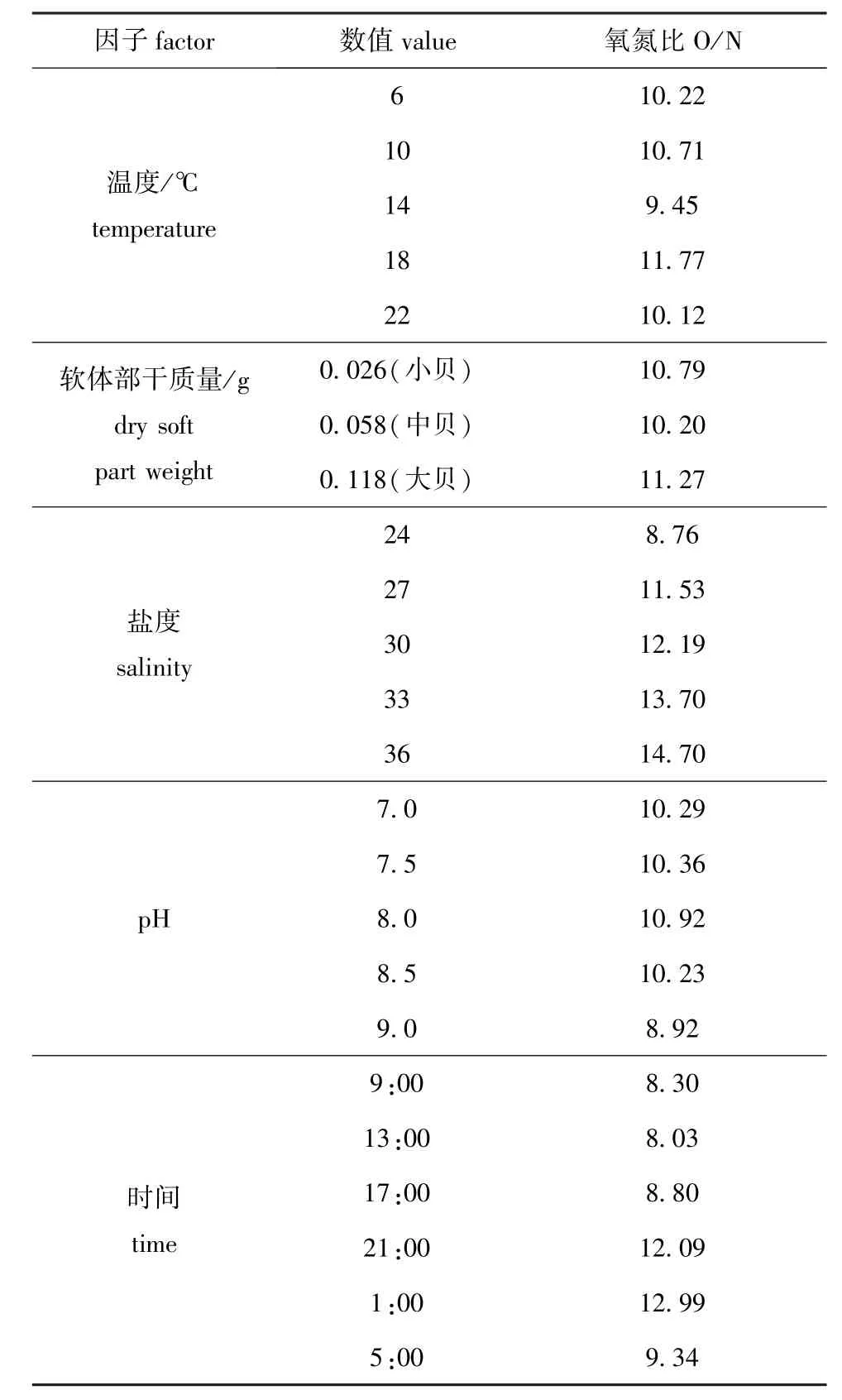
表3 岩扇贝幼贝在不同条件下的氧氮比Tab.3The oxygen-nitrogen ratio(O/N)of juvenile rock scallop Crassadoma gigantea under different condi-tions
3 讨论
3.1 温度对岩扇贝幼贝耗氧率和排氨率的影响
温度是影响水生生物代谢水平的重要因素。温度过低或过高均会导致生物体细胞理化性质和活性的改变,从而影响机体的代谢。樊甄姣等[10]研究表明,低温会导致水生生物体液的冻结,引起死亡;温度过高时,会使体内的一些帮助代谢的蛋白酶失活。毕远溥等[11]研究温度对皱纹盘鲍耗氧率和排氨率的影响时发现,温度为12~20益时,皱纹盘鲍耗氧率和排氨率均呈上升趋势,温度为20~ 28益时,耗氧率和排氨率均不断下降。杨红生等[12]对墨西哥湾扇贝的研究表明,墨西哥湾扇贝的耗氧率和排氨率均随温度升高而上升,当温度超过其耐受范围时,其耗氧率和排氨率则降低。本试验结果表明,当温度为6~22益时,岩扇贝幼贝耗氧率与排氨率均呈上升趋势,表明岩扇贝幼贝随着水温的升高,其代谢水平也不断升高,也说明本试验设定的温度区间仍在岩扇贝幼贝的适宜温度内,这与原产地岩扇贝的适宜水温(6~20益)接近。
另外,水温变化对生物代谢的直接影响可从Q10值上反映出来。Q10为温度系数,即每升高10益代谢反应变化的程度。Q10值越高,表明动物对温度变化的反应越敏感[13],Q10值越低,表明动物对温度变化越迟钝。据报道,双壳类的耗氧率Q10值为1.0~2.5,平均值为2.0[14]。本试验结果表明,岩扇贝幼贝的耗氧率Q10值为1.40~2.76,在14~18益时Q10值最高,其他区间稍低,说明岩扇贝幼贝在此区间对温度较为敏感,代谢旺盛,但温度略上升或下降也会引起耗氧率的明显变化,这可能是因为此区间是岩扇贝的最适生长水温,水温的变化对代谢影响更明显所致。
3.2 体质量对岩扇贝幼贝耗氧率和排氨率的影响
一般来说,生物体单位个体的耗氧率随体质量的增大而增大,单位体质量的耗氧率随体质量的增加而降低,其主要原因是因为个体小的生物体(体质量轻)正处在生长发育过程中,代谢活动要比个体大的生物体(体质量重)旺盛。本试验中,岩扇贝幼贝也符合这一规律,说明岩扇贝幼贝正值快速生长发育期,其耗氧率、排氨率与软体部干质量的关系也符合Y=a Wb模型。最初此模型由Win-berg提出,主要应用于鱼类代谢率(M)与体质量(W)的相关关系为M=aWb,b值的大小反映了体质量对代谢影响的程度和方式[15],随后被国内外的研究者广泛应用于多种水生动物中。
3.3 盐度对岩扇贝幼贝耗氧率和排氨率的影响
盐度是海水养殖中的主要环境因子,对生物体的排氨率有重要影响,Navarro[16]研究发现,在盐度为18~30时,Choramytilus chorus的排氨率随盐度的升高而升高。范德朋等[17]对缢蛏耗氧率及排氨率的研究表明,盐度对缢蛏耗氧率和排氨率均有极显著影响,在盐度为22时均达到最大值,大于或小于22时则呈下降趋势。王芳等[18]对二倍体和三倍体长牡蛎呼吸和排泄的研究也有相似结果,在试验盐度范围内,二倍体和三倍体长牡蛎的耗氧率、排氨率均随盐度的增大而增大。这与本试验的研究结果相一致,本试验中观察了不同盐度下岩扇贝幼贝的表观生理活动,在较低盐度(24~27)和较高盐度(33~36)环境中,岩扇贝幼贝外套膜及触手伸展幅度有所减小,张壳程度也较正常时小。这可能是因低盐环境迫使贝类机体渗透压发生改变,导致贝类主动关闭贝壳,将机体与低盐环境隔离,保护机体免受低盐环境的侵害,此现象是贝类长期适应自然环境而进化出的一种生理性保护措施。但关闭贝壳的同时减少了贝类与外界环境的水交换,通过外界环境获得的氧气减少,机体必须降低生理代谢水平以保护动物体免于窒息,同时,水体中盐度的升高会伴随着水体溶解氧饱和度的降低也是不可忽视的原因。岩扇贝是广温性种类,分布范围广对盐度的变化有良好的适应性,从本试验结果也可知,岩扇贝幼贝在所设定的5个盐度组内均有一定的呼吸代谢活动。
3.4 pH对岩扇贝幼贝耗氧率和排氨率的影响
Harris等[19]研究表明,当pH为6.08~6.72时,鲍鱼Haliotis rubra的耗氧率明显低于pH为8.45时,鲍鱼的耗氧率与pH呈正相关关系,当pH>8.45时,耗氧率与pH呈负相关关系;吴常文等[20]在研究杂交鲟Hybrid stureno时也发现,在适宜的pH范围内,杂交鲟的耗氧率随pH的升高而升高,超出适宜范围后,耗氧率显著下降。本试验结果与上述研究有类似结果,岩扇贝耗氧率、排氨率在pH值为8.0时最高,低于或高于8.0时,耗氧率与排氨率均下降。另外,根据水化学原理,水中pH的变化还与二氧化碳浓度、水温有关。水温越高,水中溶解的二氧化碳越少,pH越大。向水中充氧也会使二氧化碳溶解度降低而使pH增大。
3.5 岩扇贝幼贝耗氧率和排氨率的昼夜节律
昼夜变化又叫昼夜节律(day-night rhythm),也叫日节律(daily rhythm)。地球上的光、温度、湿度等生态因子均具有明显的周期性,其中最重要的是日周期、月周期、潮汐周期、季周期和年周期。日周期是由地球的自转所引起的,它形成白昼与黑夜的交替,生物在具有这种周期性的地球上进化和发展。因而,在生物的生命活动中形成各种节律——生物节律(biologcial rhythm)是必然的。动物的昼夜活动节律是一种复杂的生物学现象,它是对各种环境条件和昼夜变化的一种综合性的适应,包括对各种光、温度、湿度等非生物条件和食物条件,以及种内社群关系和天敌等种间关系这些生物因素的适应。因此,各种生物的昼夜活动节律均具有其本身的特点,以及各自对外界环境条件综合适应的特点。
对贝类昼夜呼吸的研究表明,白天的呼吸强度高于晚上[15]。Navarro[16]认为,这种生物的节律是由盐度、温度、摄食和光周期等日变化所控制的。而Childress等[21]则认为,海洋环境中,很多环境条件较为稳定,动物代谢的昼夜变化主要与水体中的溶解氧的日变化及动物为消除溶氧竞争而采取的生存策略有关。刘其根等[22]研究表明,河蚬耗氧率的高峰恰好出现在水体中溶氧的高峰。作者认为,贝类作为较为低等的海洋生物,环境中的各种理化因子的变化均会引起贝类自身生理活动的变化,若理化因子呈规律性的周期变化,则贝类的生理活动也呈规律性的周期变化。本试验全程保持黑暗,其他因子不变,岩扇贝幼贝一昼夜的耗氧率与排氨率略有变化,结合笔者已做研究的昼夜摄食情况分析[9],摄食率较大的时间段,也恰好是耗氧率、排氨率较高的时间段。
[1]Phleger C F,Holtz R B,Grimes PW,et al.Chemical and sensory analysis of the purple-hinge rock scallo Hinnitesmultirugosus Gale [J].Journal of Food Science,1978,43(6):1793-1796.
[2]Maxwell-Miller G,Josephson R V,Spindler A A,et al.Chilled(5益)and frozen(-18益)storage stability of the purple-hinge rock scallop,Hinnitesmultirugosus Gale[J].Journal of Food Sci-ence,1982,47(5):1654-1661.
[3]Monical Jr JB.Comparative studies on growth of the purple-hinge rock scallop Hinnitesmultirugosus(GALE)in southern California [J].Journal of the World Aquaculture Society,1979,10(1-4): 648.
[4]Laur佴n D J.Oogenesis and protandry in the purple-hinge rock scallop,Hinnitesgiganteus,in upper Puget Sound,Washington,U. S.A.[J].Canadian Journal of Zoology,1982,60(10):2333-2336.
[5]Cary SC,Leighton D L,Phleger C F.Food and feeding strategies in culture of larval and early juvenile purple-hinge rock scallops, Hinnitesmultirugosus(GALE)[J].Journal of theWorld Aquacul-ture Society,1981,12(1):156-169.
[6]Culver C S,Richards JB,Page H M.Plasticity of attachment in the purple-hinge rock scallop,Crassadoma gigantea:implications for commercial culture[J].Aquaculture,2006,254(1-4):361-369.
[7]曹善茂,汪健,王谦,等.岩扇贝人工育苗的初步研究[J].大连海洋大学学报,2017,32(1):1-6.
[8]曹善茂,王昊,陈炜,等.岩扇贝闭壳肌营养成分的分析及与中国3种扇贝的比较[J].大连海洋大学学报,2016,31(5):544-550.
[9]曹善茂,梁伟锋,汪健,等.岩扇贝幼贝滤食率的基础研究[J].大连海洋大学学报,2016,31(6):612-617.
[10]樊甄姣,吴文常,皇甫淑燕.温度、盐度、pH对角蝾螺排氨率的影响[J].渔业现代化,2007,34(3):10-12.
[11]毕远溥,蒋双,刘海映,等.温度、体重对皱纹盘鲍耗氧量和排氨量的影响[J].应用与环境生物学报,2000,6(5):444-446.
[12]杨红生,张涛,王萍,等.温度对墨西哥湾扇贝耗氧率及排泄率的影响[J].海洋学报,1998,20(4):91-96.
[13]Criffiths C L,Criffiths R J.Bivalvia[M]//Pandian T J,Vernberg F J.Animal Energetics.San Diego:Academic Press,1987,2:2-88.
[14]W llbur A E,Hilbish T J.Physiological energetics of the ribbed mussel Geukensia demissa(Dillwyn)in response to increased temperature[J].Journal of Experimental Marine Biology and E-cology,1989,131(2):161-170.
[15]刘勇,施坤涛,张少华,等.双壳贝类呼吸代谢的研究进展[J].南方水产,2007,3(4):65-69.
[16]Navarro JM.The effects of salinity on the physiological ecology of Choromytilus chorus(Molina,1782)(Bivalvia:Mytilidae)[J]. Journal of Experimental Marine Biology and Ecology,1988,122 (1):19-33.
[17]范德朋,潘鲁青,马牲,等.盐度和pH对缢蛏耗氧率及排氨率的影响[J].中国水产科学,2002,9(3):234-238.
[18]王芳,王昭萍,董双林,等.盐度对二倍体和三倍体长牡蛎呼吸和排泄的影响[J].海洋科学,2003,27(6):73-76.
[19]Harris JO,Maguire G B,Edwards S J.Effect of pH on growth rate,oxygen consumption rate,and histopathology of gill and kid-ney tissue for juvenile greenlip abalone,Haliotis laevigata Dono-van and blacklip abalone,Haliotis rubra Leach[J].Journal of Shellfish Research,1981,18(2):611-619.
[20]吴常文,朱爱意,赵向炯.海水养殖杂交鲟对环境变化耐受性的试验研究[J].水产科学,2005,24(9):1-4.
[21]Childress JJ,Taylor SM,Cailliet G M,et al.Patterns of growth, energy utilization and reproduction in some meso-and bathype-lagic fishes off Southern California[J].Marine Biology,1980,61 (1):27-40.
[22]刘其根,沈和定,周洪琪,等.河蚬的耗氧率和排氨率[J].上海海洋大学学报,1999,8(4):298-303.
Oxygen consum ption rate and ammonia excretion rate of juvenile rock scallop Crassadoma gigantea in different ecological factors
CAO Shan-mao,LIANGWei-feng,LIU Gang,LIXiao-yu
(Engineering Research Center of Shellfish Culture and Breeding in Liaoning Province,Dalian Ocean University 116023,China)
In this study,the effects of temperature,salinity,pH,and body weighton oxygen consumption rate and ammonia excretion rate were studied in rock scallop Crassadoma gigantea introduced in 2013 to provide the basis for study of the physiological ecology of the scallop and the guidance to production practice.The results showed that oxygen consumption rate and ammonia excretion rate in juvenile rock scallop were significantly affected by tempera-ture,salinity,pH,and body weight(P<0.01),increased with the rise of temperature within 6-22益.The regres-sion analysis revealed that relationship between body weight(X)and oxygen consumption rate or ammonia excretion rate(Y)was expressed as negative exponential relationship.The oxygen consumption rate and ammonia excretion rate per unitbody weightwere decreased with the increase in bodyweight,and increased with the increase in salin-ity from 24 to 30,themaximum at salinity of30,and then decreased with the increase in salinity from 30 to 36;in-creased with the increase in pH from 7.0 to 8.0,themaximum at salinity of 8.0,and then decreased with the in-crease in salinity from 8.0 to 9.0.The peak of oxygen consumption rate and ammonia excretion rate were observed at13:00,theminimal oxygen consumption rate at 21:00 and the minimal ammonia excretion rate at 1:00.The findings provide reference with physiological and ecological research in rock scallop.
juvenile Crassadoma gigantea;oxygen consumption rate;ammonia excretion rate;diurnal rhythm
S917.4
A
10.16535/j.cnki.dlhyxb.2017.03.005
2095-1388(2017)03-0280-07
2016-12-28
辽宁省海洋与渔业厅项目(201214);大连市海洋与渔业局项目(2012011)
曹善茂(1960—),男,副教授。E-mail:caoshm@dlou.edu.cn
猜你喜欢
家教世界·创新阅读(2019年11期)2019-12-10
家教世界(2019年31期)2019-12-05
水产科学(2018年3期)2018-06-04
人生与伴侣·共同关注(2018年8期)2018-02-16
河北渔业(2015年7期)2015-07-21
中国新技术新产品(2015年3期)2015-07-19
中国洗涤用品工业(2015年5期)2015-02-28
应用海洋学学报(2014年1期)2014-11-22
河北渔业(2014年3期)2014-07-08
浙江海洋大学学报(自然科学版)(2014年6期)2014-03-13
