
菲律宾蛤仔闭壳肌组织学及其蛋白特性
2017-06-19 18:56张龙刘俊荣田元勇姜梦云刘慧慧郑尧宋扬刘金洋大连海洋大学食品科学与工程学院辽宁大连116023
大连海洋大学学报 2017年3期
张龙,刘俊荣,田元勇,姜梦云,刘慧慧,郑尧,宋扬,刘金洋(大连海洋大学食品科学与工程学院,辽宁大连116023)
菲律宾蛤仔闭壳肌组织学及其蛋白特性
张龙,刘俊荣,田元勇,姜梦云,刘慧慧,郑尧,宋扬,刘金洋
(大连海洋大学食品科学与工程学院,辽宁大连116023)
为探讨菲律宾蛤仔Ruditapes philippinarum的加工特性与肌肉蛋白质特性的关系,对菲律宾蛤仔闭壳肌的组织形态、肌原纤维蛋白(Myofibrillar,Mf)的分布特性,以及与其低温凝胶化相关的特性等进行了分析。结果表明:闭壳肌肌纤维具有高度同向性,横纹肌呈直线柱状,平滑肌呈螺旋柱状;利用SDSPAGE电泳对两种类型肌肉的蛋白分布分析显示,横纹肌肌浆蛋白组分中存在相对分子质量为23 000的差异蛋白,且溶出更多的粗丝蛋白,平滑肌的Mf组分中存在标志性蛋白Myorod,且副肌球蛋白含量高,肌球和原肌球蛋白含量低,肌动蛋白含量接近;整个闭壳肌Mf的溶解性依赖于离子强度和有无三磷酸腺苷(Adenosine Triphosphate,ATP)的添加;Ca2+-ATPase活性在中等离子强度(KCl浓度为0.30 mol/L)时最高,偏离中等离子强度后活性逐渐降低;参与交联的蛋白包括肌球蛋白、副肌球蛋白、原肌球蛋白,离子强度越高交联作用越显著。本研究结果可为探究经济贝类软体可食部位蛋白特性提供参考。
菲律宾蛤仔;横纹肌;平滑肌;肌原纤维蛋白;蛋白特性
水产无脊椎动物肌肉组织及蛋白特性不同于水产脊椎动物,贝类肌肉组织除横纹肌外,还包括平滑肌,且副肌球蛋白含量较高。王帅等[1]将栉孔扇贝闭壳肌蛋白添加到鲢肌肉中制成蛋白凝胶,显著提高了产品的凝胶强度;Fukuda等[2]研究发现,副肌球蛋白的存在对肌肉凝胶特性具有显著影响,含量更高的平滑肌形成的凝胶硬而脆;对扇贝横纹闭壳肌盐溶性蛋白的热凝胶性研究发现,相比90益下加热,低温(25益)凝胶化过程对蛋白凝胶发挥了更重要的作用,这是由于谷氨酰胺转移酶TGase的交联作用[3]。鱼糜凝胶是鱼糜加盐擂溃使肌动球蛋白溶出后,发生蛋白的变性和聚集,进而形成三维网络结构[4-5]。
菲律宾蛤仔Ruditapes philippinarum是中国四大养殖贝类之一,也是辽宁省最主要的滩涂贝类养殖品种[6]。菲律宾蛤仔作为重要的水产蛋白源,其食品原料学属性却被较少关注。笔者在已往研究中,对菲律宾蛤仔软体部位含氮物的组成及分布进行了剖析[6],本研究中,对闭壳肌进行组织学观察,以肌原纤维蛋白(Myofibrillar,Mf)为主线,重点考察Mf的分布特性及其在低温凝胶化过程中的相关特性,包括溶解性、Ca2+-ATPase活性、TGase作用下的交联特性,旨在为探究经济贝类软体可食部位蛋白特性提供参考。
1 材料与方法
1.1 材料
试验用菲律宾蛤仔购自大连市长兴水产品市场(2015年9—12月),规格为25~30个/kg。
主要仪器设备有GL-21M高速冷冻离心机(湖南湘仪实验仪器开发有限公司),Synergy H1全功能酶标仪(美国伯腾仪器有限公司),Bio-Rad垂直平板电泳槽(北京伯乐生命科学发展有限公司),PHS-3C精密pH计(上海精密科学有限公司),Leica DMI6000倒置荧光显微镜(北京中仪光科科技发展有限公司)。
主要试剂有SDS-聚丙烯酰胺凝胶电泳试剂, TGase(美国Sigma公司),蛋白标准品(美国BioLabs公司),KCl、CaCl2、NaCl(中国天津市科密欧化学试剂有限公司),ATP(中国上海华蓝化学科技有限公司),钼酸铵(中国上海晶纯生化科技股份有限公司),TRIS、EGTA、EDTA(中国北京索莱宝科技有限公司),各试剂均为分析纯或化学纯。
1.2 方法
1.2.1 肌浆蛋白和肌原纤维蛋白的提取用解剖刀将新鲜原料的闭壳肌切断,除去体腔液,以便观察其软体组织结构。进一步对闭壳肌进行分割,细分为横纹闭壳肌和平滑闭壳肌。参照Hashimoto等[4]的方法进行肌浆蛋白和肌原纤维蛋白的分离提取。
1.2.2 组织学分析参照Mizuta等[5,7]和Lee等[8]的方法,分别取横纹闭壳肌和平滑闭壳肌组织块,置于卡诺氏液中固定,用常规石蜡进行包埋,切片,用苏木精-伊红染色后,利用光学显微镜对其进行组织学观察。
1.2.3SDS-PAGE及电泳谱图定量分析在所有蛋白样品中加入电泳上样液[含有2%SDS、8 mol/L尿素、2%茁-巯基乙醇、50 mmol/L Tris-HCl(pH 8.0)]使其溶解,在100益下加热3 min,用于分析交联特性的样品用磁力搅拌至完全溶解。采用SDS-聚丙烯酰胺凝胶电泳进行蛋白质组分分子量的测定,其中浓缩胶质量分数为5%,分离胶质量分数为7.5%,采用R-250考马斯亮蓝染色法检验,用醋酸甲醇溶液进行脱色。扫描胶片后得到电泳图谱,并使用Multi Gauge 2.0软件对图谱进行定量分析。
1.2.4 溶解性的测定将提取的闭壳肌Mf溶液分别调至NaCl终浓度为0.05、0.10、0.20、0.30、0.40、0.50 mol/L,蛋白质终浓度为5 mg/mL,4益条件下放置40 min后,于12 000 g下离心20 min,收集上清液,采用双缩脲法测定蛋白质含量,溶解度表示为上清液中蛋白质含量与总蛋白质含量的百分比。同时,设定1组含5 mmol/L(表示终浓度,下同)ATP、4 mmol/L EGTA、4 mmol/LMgCl2的试验组,放置10 min,其他操作同上。
1.2.5Ca2+-ATPase活性的测定将提取的闭壳肌Mf溶液与ATP反应液[1 mg/mL Mf、1 mmol/L ATP、5 mmol/L CaCl2、25mmol/L Tris-马来酸(pH 7.0)]混合,其中KCI设定6个浓度,分别为0.05、0.10、0.20、0.30、0.40、0.50 mol/L。20益条件下分别反应0、5、10、15、20、25、30 min,在1 mL反应液中加入0.5 mL 15%HClO4溶液终止反应。采用钼酸铵法[9]测定反应释放的无机磷(Pi)含量。Mf的Ca2+-ATPase活性单位是滋mol Pi/ (mg·min),表示1 mg肌原纤维蛋白在1 min时间内分解ATP生成无机磷酸的微摩尔数。
1.2.6 交联特性分析在提取的闭壳肌Mf溶液中添加CaCl2和TGase作为试验组,终浓度分别为10 mmol/L和0.4%,以添加EGTA作为空白对照组,终浓度为6 mmol/L。将对照组与试验组中的NaCl设为0.05、0.10、0.20、0.30、0.40、0.50 mol/L 6个浓度组,蛋白质终浓度为5 mg/mL,置于25益条件下反应不同时间,添加EDTA终止反应后立即在100益下加热2 min,加入上述电泳上样液后用磁力搅拌过夜至完全溶解。
1.3 数据处理
所有试验组均设3个平行,使用3个不同批次的菲律宾蛤仔进行试验。试验数据用平均值依标准差表示,用SPSS 17.0软件进行单因素方差分析,采用Duncan氏新复极差检验法进行显著性差异分析,显著性水平设为0.05。
2 结果与讨论
2.1 横纹肌与平滑肌的组织学结构
如图1所示,菲律宾蛤仔整个软体部位由闭壳肌、足肌、外套膜、水管、鳃和内脏团6个组织构成,其中闭壳肌可分为横纹闭壳肌和平滑闭壳肌,两者间无明显分界线,这一结构区别于扇贝闭壳肌。菲律宾蛤仔各组织主要以平滑肌为主,本研究中选取横纹闭壳肌和平滑闭壳肌作为两种类型肌肉的代表。在光学显微镜(100倍和400倍)下分别对横纹肌与平滑肌的横纵切面进行观察,从图2可见:胶原纤维呈浅红色,肌纤维呈亮红色,细胞核和核糖体呈蓝色;以胶原蛋白为主的结缔组织存在形式由大到小依次为肌膜、肌束膜、肌内膜;闭壳肌肌纤维具有高度同向性,横纹肌呈直线柱状,平滑肌呈螺旋柱状。
横纹闭壳肌与平滑闭壳肌肌纤维在形态和分布上存在明显差异,这与肌原纤维蛋白的组成和含量密不可分,因而影响食用品质。此外,肌纤维具有动态变化性,这对跟踪表征经济贝类流通过程中的品质变化具有参考价值。
2.2 横纹肌与平滑肌的蛋白分布特性
对菲律宾蛤仔横纹闭壳肌与平滑闭壳肌的全蛋白、肌浆蛋白和肌原纤维蛋白进行SDS-PAGE电泳分析(图3),并对肌原纤维蛋白的电泳谱图进行定量分析(表1)。
两种类型肌肉的不同组分蛋白的电泳图谱有所差异。首先,从全蛋白条带图3-A可知,两种类型肌肉蛋白种类丰富,组成相似,收缩蛋白组分(粗、细丝蛋白)清晰可见。肌浆蛋白包括肌红蛋白和大量的酶类,主要是糖酵解酶系,另外还有磷酸戊糖途径酶系和肌酸激酶、AMP-脱氨酶等[10],这些蛋白可溶于水或低离子强度的中性盐溶液中,其占闭壳肌含氮物的12.55%[6]。从图3-B可见,与平滑肌相比,横纹肌肌浆蛋白组分中出现1条含量丰富的蛋白条带,相对分子质量为23 000,可能是蛋白酶或肌钙蛋白等。部分肌原纤维蛋白均出现在两种肌肉的肌浆蛋白组分中,且横纹肌中溶出的粗丝蛋白更多,说明肌原纤维蛋白在低离子强度下具有高的溶解性。这不同于鱼类,鱼类肌原纤维蛋白只能在高离子强度下溶出。鱼糜加工过程中会有水洗过程,如果将传统水洗方式应用于菲律宾蛤仔以获得肌原纤维蛋白,很可能因其高溶解性而有所损失。研究发现,扇贝[11]和鱿鱼[12]肌肉中的肌动球蛋白在低离子强度下即可溶出。
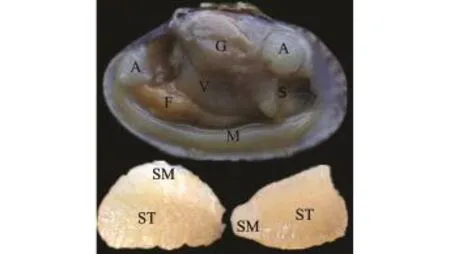
图1 菲律宾蛤仔软体部位的解剖图Fig.1Anatomy of soft part of M anila clam Ruditapes philippinarum
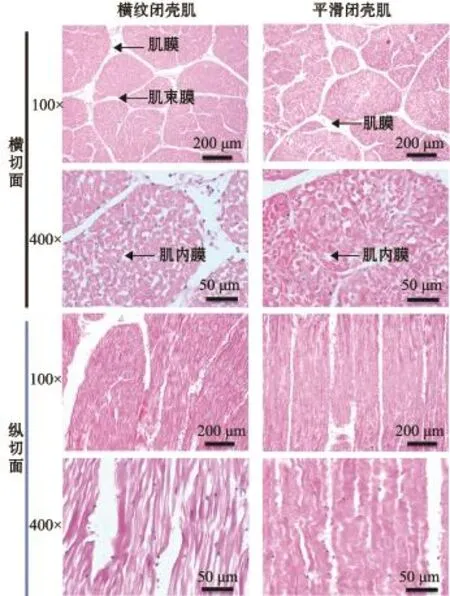
图2 菲律宾蛤仔横纹肌与平滑肌的光学显微结构Fig.2Lightm icroscopical features of the striated mus-cle and smooth muscle in M anila clam Ruditapes philippinarum
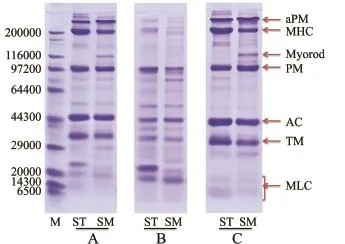
图3 菲律宾蛤仔横纹肌与平滑肌的蛋白电泳图谱Fig.3SDS-PAGE electrophoresis patterns of the stri-ated muscle and smooth muscle in M anila clam Ruditapes philippinarum
肌原纤维蛋白组分(包括粗丝蛋白和细丝蛋白)为肌肉的主要含氮物质,占闭壳肌含氮物的50.40%[6]。软体动物的粗丝蛋白包括形成核心的副肌球蛋白(Paramyoosin,PM),以及分布于表面的肌球蛋白(Myosin,MY)、Myorod(MR)[13-15]、颤搐蛋白(Twitchin)[16-18];细丝蛋白包括肌动蛋白(Actin,AC)、原肌球蛋白(Tropomyosin,TM)、肌钙蛋白(Troponin,TN)或钙调节蛋白(Calpo-nin)[19-20]。如图3-C所示,Mf中主要蛋白的相对分子质量如下:肌球蛋白重链(Myosin hight chain, MHC)为200 000,Myorod为116 000,PM为100 000,AC为45 000,TM为35 000,肌球蛋白轻链(Myosin light chain,MLC)<20 000,副肌球蛋白多聚体(Aggregated paramyosin,aPM)[16,21]> 200 000。平滑肌中存在一标志性蛋白Myorod(横纹肌中不存在),占肌原纤维蛋白的5.39%。
由Mf的电泳谱图进行定量分析(表1)得知,横纹肌与平滑肌中主要蛋白含量分别为:MHC为19.13%和12.24%,副肌球蛋白(含PM和aPM)为25.93%和36.89%,AC为23.14%和22.57%, TM为22.97%和17.38%,MLC为8.84%和5.53%,Myorod为0和5.39%。与横纹肌相比,平滑肌中副肌球蛋白含量高,肌球和原肌球蛋白含量低,肌动蛋白含量接近。
有研究报道,副肌球蛋白在扇贝、鱿鱼和牡蛎横纹肌肌原纤维蛋白中分别约占3%、14%、19%,在牡蛎平滑肌肌原纤维蛋白中约占38%[22]。因此,无脊椎动物平滑肌中副肌球蛋白的含量相对较高。笔者发现,在菲律宾蛤仔闭壳肌中含有丰富的副肌球蛋白多聚体,而相关贝类的研究报道中尚未明确指出。菲律宾蛤仔平滑闭壳肌肌肉中还存在标志性蛋白Myorod,Shelud蒺ko等[14]研究表明, Myorod相对分子质量为110 000~130 000,肌肉中含量为5%~6%,扇贝、牡蛎、帘蛤和蚶蜊中只有1条多肽链,贻贝中含有的相对分子质量接近,含量相同的两条链(琢和茁链)。
2.3 闭壳肌肌原纤维蛋白的溶解性
离子强度和ATP对闭壳肌肌原纤维蛋白溶解性的影响如图4所示,无ATP添加时,肌原纤维蛋白的溶解度随离子强度(NaCl浓度为0.05~0.50 mol/L)的增加而明显上升,从14.27%迅速上升到72.92%。离子强度的增加使蛋白表面吸附更多的盐离子,带电表层使蛋白质分子间排斥作用增强,蛋白分子与水分子间的水化作用增大,因而溶解性提高[23]。添加5 mmol/L ATP后,肌原纤维蛋白的溶解度随着离子强度(NaCl浓度为0.05~0.50 mol/L)的增加而缓慢增加,其溶解度为61.82%~ 91.94%,始终处于较高水平。
综上所述,肌原纤维蛋白溶解度随ATP(决定粗丝和细丝解离程度)和NaCl浓度(改变蛋白质分子周围的离子氛)的增加而增大,肌原纤维蛋白溶解度提高主要是由粗丝蛋白和细丝蛋白解离引起的,其中ATP比NaCl发挥了更重要的作用。Niki等[24]发现,扇贝横纹肌中ATP含量越高,低盐溶性蛋白的溶出率就越高,相应的凝胶性就越好。这说明原料越新鲜,Mf溶解状态越好,越有利于凝胶的形成。
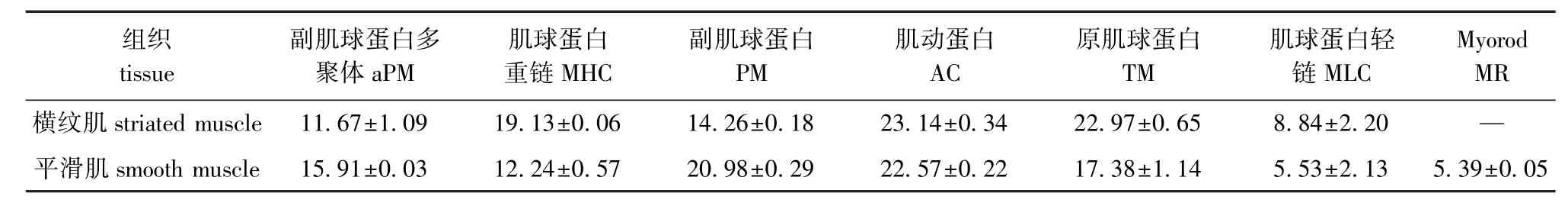
表1 菲律宾蛤仔横纹肌与平滑肌中肌原纤维蛋白的分布T ab.1M yofibrillar proteins distribution in the striated muscle and smooth muscle of M anila clam Ruditapes philippinarum %
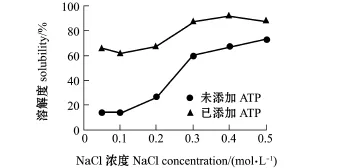
图4 离子强度和ATP对菲律宾蛤仔闭壳肌肌原纤维蛋白溶解性的影响Fig.4Influence of ionic strength on solubilities ofmyo-fibrillar proteins in adductor of Manila clam Ruditapes philippinarum
2.4 闭壳肌肌原纤维蛋白的Ca2+-ATPase活性
试验中以Ca2+-ATPase活性为肌球蛋白变性指示参数,在不同离子强度下,分析了闭壳肌肌原纤维蛋白稳定性的变化规律(表2)。结果表明,闭壳肌肌原纤维蛋白Ca2+-ATPase活性为0.20~0.30滋mol Pi/(min·mg);在中等离子强度(0.30 mol/L KCl)时Ca2+-ATPase活性最高,偏离中等离子强度后活性逐渐降低。因此,离子强度会影响肌球蛋白分子结构的稳定性,高或低离子强度均可导致肌球蛋白头部结构发生较大变化,引起Ca2+-ATPase活性下降。
在对虾夷扇贝闭壳肌Mf[25]、栉孔扇贝闭壳肌肌球蛋白[26]、鲤Mf[27]等的相关研究中也发现, Ca2+-ATPase最大活性出现在中等离子强度范围内。与鱼类相比,菲律宾蛤仔闭壳肌Mf的Ca2+-ATPase活性(0.50 mol/L KCl)时是鲢Mf的2.6倍[28]。Hayashi等[29]研究表明,高离子强度对鲤Mf中Ca2+-ATPase活性有两种抑制方式,即高离子强度抑制S-1片段(胰蛋白酶切后肌球蛋白头部)活性和降低F-actin的保护作用。
2.5 闭壳肌肌原纤维蛋白的交联特性
TGase具有Ca2+依赖性,能够促进着-(酌-谷酰基)赖氨酸的交联[30-31],添加EGTA为对照组,由于电泳上样液含有尿素、SDS和茁-巯基乙醇,能打开蛋白分子间除非二硫共价键外的所有化学键,所以,本研究中只考察由TGase引起的着-(酌-谷酰基)赖氨酸的交联。通过设定不同离子强度和放置时间,分析Mf各组分在交联过程中的参与情况及含量变化。
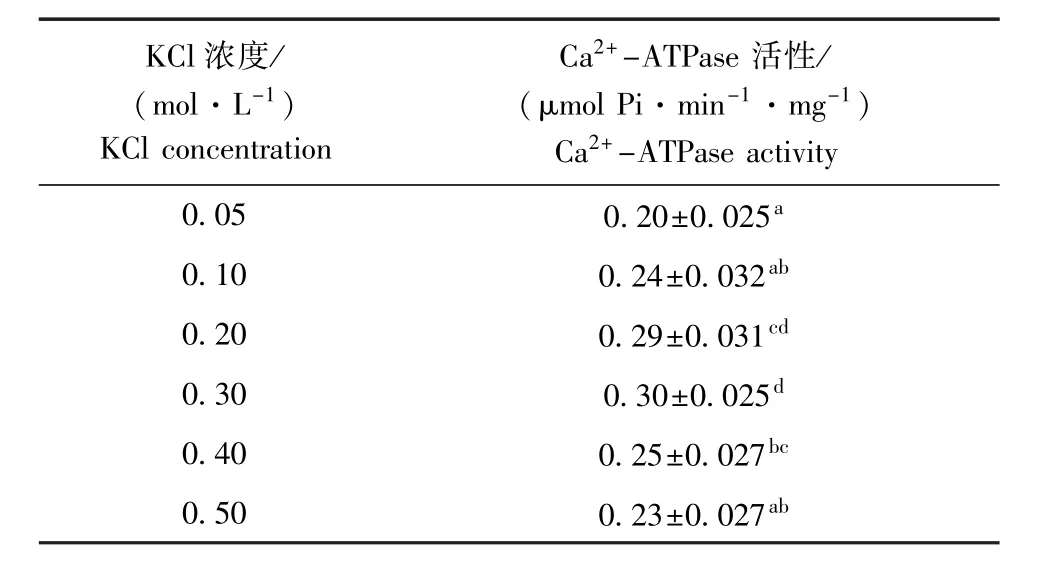
表2 离子强度对菲律宾蛤仔闭壳肌肌原纤维蛋白Ca2+-ATPase活性的影响Tab.2Influence of ionic strength on Ca2+-ATPase activity ofm yofibrillar proteins in adductor of M anila clam Ruditapes philippinarum
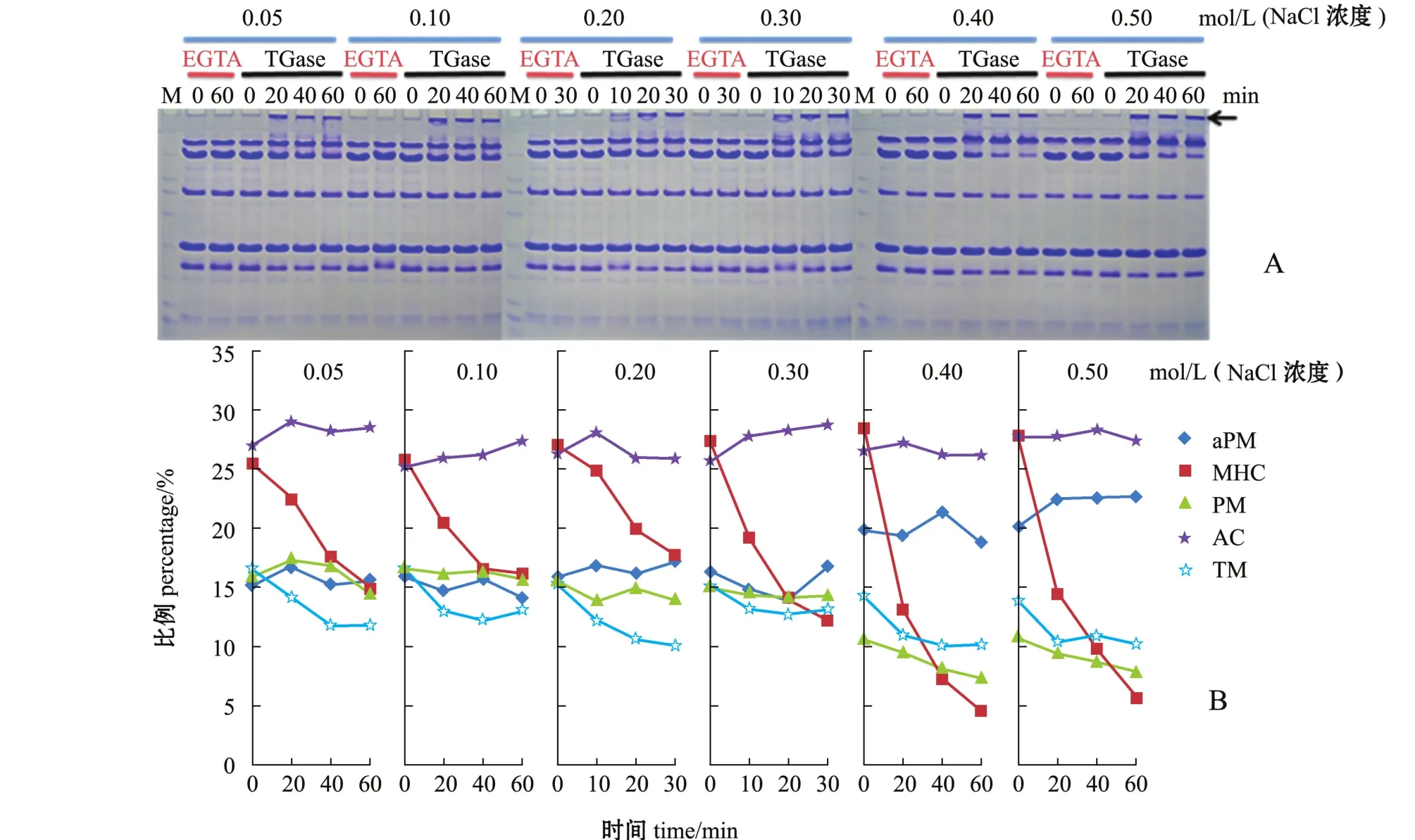
图5 离子强度和TGase对菲律宾蛤仔闭壳肌肌原纤维蛋白交联特性的影响Fig.5Influence of ionic strength and TGase on crosslinking feature ofm yofibrillar proteins in adductor muscle of M anila clam Ruditapes philippinarum
本试验中对放置不同时间的肌原纤维蛋白进行SDS-PAGE电泳及定量分析(图5),结果表明,随着离子强度的增加,AC含量无明显变化,MHC含量消失速率加快,相应的MHC多聚体(箭头所示)明显增多。表明TGase催化MHC形成了着-(酌-谷酰基)赖氨酸键,而对肌动蛋白未有影响。当离子强度达到0.40、0.50 mol/L时,PM含量才出现下降趋势,同时aPM含量处于较高水平,说明PM在此条件下参与了交联过程,且交联程度较小。此外,TM含量在各个离子强度下均随时间增加而略有下降,MHC消失的同时,其相对分子质量有所增加,可推测TM通过与MHC结合而发生交联。综合有关低温凝胶化相关特性可知,高离子强度下,Mf的溶解状态好,变性程度大,TGase作用下的交联程度强,更有利于其低温凝胶化过程。
Fukuda等[2]通过动态流变学特性研究了扇贝平滑肌中副肌球蛋白的热诱导凝胶,副肌球蛋白的存在对肌肉凝胶特性有显著影响,含量高的平滑肌形成的凝胶硬而脆;陆海霞[32]运用聚丙烯酰胺凝胶和环境扫描电子显微镜等技术研究发现,头足类生物特有的副肌球蛋白也参与热诱导凝胶的形成。徐辉[33]通过SDS-PAGE分析凝胶蛋白质的变化发现,随着TGase浓度的增加PM带强度均会减小,表明TGase催化体系中副肌球蛋白的交联促进了蛋白质的聚合,从而提高了凝胶的硬度和弹性。其中副肌球蛋白参与交联过程的结论与本研究结果相同。然而,Yoshida等[3]对扇贝横纹闭壳肌盐溶性蛋白热凝胶特性的研究显示:添加Ca2+并在25益下放置4 h,可激活其内源TGase使凝胶性显著增强,其中促进了肌球蛋白重链的交联,而肌动蛋白和副肌球蛋白无变化;Ca2+的添加未能影响副肌球蛋白的热凝胶特性,TGase很难交联副肌球蛋白分子。Park等[34]研究了日本鱿鱼外套膜肌肉凝胶形成能力及副肌球蛋白的凝胶机制,结果发现,在0.50 mol/L NaCl时,副肌球蛋白和肌动蛋白未参与低温凝胶化过程的交联,肌球蛋白是参与交联的主要成分。其中副肌球蛋白未参与交联过程的结论与本研究结果不同。
从以上关于无脊椎动物肌肉凝胶性的报道可见,无脊椎动物肌肉的凝胶形成能力和热凝胶质地不同于鱼类,本质差异在于其肌肉组织类型和蛋白组成及特性的不同,例如副肌球蛋白和原肌球蛋白的参与,肌球蛋白与副肌球蛋白的含量,蛋白的热稳定性等。目前,尚未见原肌球蛋白参与交联的报道,此外,副肌球蛋白参与凝胶形成也需一定条件,例如内源TGase对Ca2+和离子强度的依赖性、温度和pH等。
综上所述,菲律宾蛤仔横纹闭壳肌与平滑闭壳肌的组织形态及肌原纤维蛋白分布特性存在明显差异,这种差异与肌肉的食用品质密切相关;在不同离子强度下,整个闭壳肌肌原纤维蛋白低温凝胶化相关的特性,呈现了较强的规律性;高离子强度下肌原纤维蛋白溶解状态好,变性程度大,TGase作用下交联程度强,更有利于其低温凝胶化过程。值得关注的是,闭壳肌肌肉中含有丰富的副肌球蛋白多聚体,且副肌球蛋白和原肌球蛋白均参与了TGase作用下的交联过程。
[1]王帅,刘俊荣,辛草,等.扇贝闭壳肌蛋白对鲢鱼肌肉蛋白凝胶的修饰[J].水产科学,2011,30(11):659-665.
[2]Fukuda N,Fujiura M,Kimura M,et al.Thermally induced gelation of paramyosin from scallop adductormuscle[J].Fisheries Science, 2006,72(6):1261-1268.
[3]Yoshida W,Kunimi O,Fujiura M,et al.Thermal gelation of salted paste from scallop striated adductormuscle[J].Fisheries Science, 2003,69(5):1017-1025.
[4]Hashimoto K,Watabe S,Kono M,etal.Muscle protein composition of sardine and mackerel[J].Bulletin of the Japanese Society of Scientific Fisheries,1979,45(11):1435-1441.
[5]Mizuta S,Miyagi T,Nishimiya T,et al.Partial characterization of collagen in several bivalvemolluscs[J].Food Chemistry,2004,87 (1):83-88.
[6]张龙,刘俊荣,田元勇,等.菲律宾蛤仔(Ruditapes philippina-rum)软体部位含氮物的组成及其分布[J].食品科学,2015,36 (24):95-100.
[7]Mizuta S,Miyagi T,Nishimiya T,et al.Partial characterization of collagen inmantle and adductor of pearl oyster(Pinctada fucata) [J].Food Chemistry,2002,79(3):319-325.
[8]Lee JS,Lee Y G,Park J J,et al.Microanatomy and ultrastructure of the footof the infaunal bivalve Tegillarca granosa(Bivalvia:Ar-cidae)[J].Tissue and Cell,2012,44(5):316-324.
[9]Fiske C H,Subbarow Y.The colorimetric determination of phos-phorus[J].The Journal of Biological Chemistry,1925,66(2): 375-400.
[10]Foegeding E A,Lanier T C,Hultin H O.Characteristics of edible muscle tissues[M]//Fennema O R.Food Chemistry.New York: Marcel Dekker,1996:879-942.
[11]Matsumoto JJ,DyerW J,Dingle JR,et al.Protein in extracts of prerigor and postrigor scallop striated muscle[J].Journal of the Fisheries Research Board of Canada,1967,24(4):873-882.
[12]De Jesus Encinas-Arzate J,Ezquerra-Brauer J M,Oca觡o-Higuera V M,et al.Effectof ionic strength on soluble protein re-moval from giant squid mantle(Dosidicusgigas)and functional evaluation of protein recovery[J].Food Science and Biotechnol-ogy,2014,23(2):401-407.
[13]B佴jar P,Villamar侏n JA.Catalytic subunit of cAMP-dependent protein kinase from a catchmuscle of the bivalvemollusk Mytilus galloprovincialis:purification,characterization,and phosphoryla-tion of muscle proteins[J].Archives of Biochemistry and Bio-physics,2006,450(2):133-140.
[14]Shelud蒺ko N S,Tuturova K F,Permyakova T V,et al.A novel thick filament protein in smoothmuscles of bivalvemolluscs[J]. Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,1999,122(3):277-285.
[15]Shelud蒺ko N,Permjakova T,Tuturova K,etal.Myorod,a thick fil-ament protein inmolluscan smoothmuscles:isolation,polymeriza-tion and interaction with myosin[J].Journal of Muscle Research &Cell Motility,2001,22(1):91-100.
[16]Shelud蒺ko N S,Matusovsky O S,Permyakova T V,et al.“Twitchin-actin linkage hypothesis冶for the catch mechanism in mol-luscanmuscles:evidence that twitchin interacts withmyosin,my-orod,and paramyosin core and affects properties of actomyosin [J].Archives of Biochemistry and Biophysics,2007,466(1): 125-135.
[17]Shelud蒺ko N S,Matusovskaya G G,Permyakova T V,et al.
Twitchin,a thick-filament protein from molluscan catch muscle, interacts with F-actin in a phosphorylation-dependentway[J].Archives of Biochemistry and Biophysics,2004,432(2):269-277.
[18]Tsutsui Y,Yoshio M,Oiwa K,etal.Striatedmuscle twitchin ofbi-valves has“catchability冶,the ability to bind thick filaments tight-ly to thin filaments,representing the catch state[J].Journal of Molecular Biology,2007,365(2):325-332.
[19]Funabara D,Nakaya M,Watabe S.Isolation and characterization of a novel 45 kDa calponin-like protein from anterior byssus re-tractormuscle of themussel Mytilus galloprovincialis[J].Fisher-ies Science,2001,67(3):511-517.
[20]Dobrzhanskaya A V,Matusovskaya G G,Matusovsky O S,et al. Thin filaments of bivalve smooth musclemay contain a calponinlike protein[J].Biophysics,2010,55(5):703-706.
[21]Watabe S,Iwasaki K,Funabara D,et al.Complete amino acid se-quence of Mytilus anterior byssus retractor paramyosin and its pu-tative phosphorylation site[J].Journal of Experimental Zoology, 2000,286(1):24-35.
[22]Sano T,Fnoguchi S,Tsuchiya T,et al.Contribution of paramyosin tomarinemeat gel characteristics[J].Journal of Food Science, 1986,51(4):946-950.
[23]杨志敏,蒋立科.生物化学[M].北京:高等教育出版社, 2005:69-70.
[24]Niki T,Kato Y,Nozawa H,et al.Gelation of low salt soluble pro-teins from scallop adductormuscle in relation to its freshness[J]. Fisheries Science,2002,68(3):688-693.
[25]吴忠,刘俊荣,田元勇.虾夷扇贝闭壳肌和外套膜肌原纤维蛋白的特性分析[J].水产学报,2015,39(11):1640-1649.
[26]Mori K,Ojima T,Nishita K.Comparative studies on ATPase activ- ity and filament assembly of akazara and vertebratemyosins[J]. Bulletin of the Japanese Society of Scientific Fisheries(Japan), 1989,55(10):1851-1856.
[27]Koseki S,Nomura K,Konno K.Development ofmyofibrillar AT-Pase assay system on pH stat[J].Fisheries Science,2005,71 (2):380-387.
[28]Zheng Liang,Yu Kefeng,Yuan Chunhong,et al.Characterization ofmyosin subfragment-1 of summer and winter silver carp(Hy-pophthalmichthysmolitrix)muscle[J].Journal of Food Science, 2012,77(9):C914-C920.
[29]Hayashi K,Konno K.Stabilization ofmyosin by ionic compounds as affected by F-actin[J].Fisheries Science,2006,72(6): 1306-1312.
[30]Jia Dan,You Juan,Hu Yang,et al.Effect of CaCl2on denatur-ation and aggregation of silver carp myosin during setting[J]. Food Chemistry,2015,185:212-218.
[31]姜梦云,刘俊荣,田元勇,等.蛋白质交联与食品蛋白配料功能性的研究进展[J].大连海洋大学学报,2016,31(5):576-582.
[32]陆海霞.秘鲁鱿鱼肌原纤维蛋白质凝胶特性的研究[D].杭州:浙江工商大学,2009.
[33]徐辉.谷氨酰胺转氨酶对秘鲁鱿鱼肌原纤维蛋白凝胶特性的影响[D].杭州:浙江工商大学,2007.
[34]Park S,Cho S,Yoshioka T,et al.Influence of endogenous protea-ses and transglutaminase on thermal gelation of salted squid mus-cle paste[J].Journal of Food Science,2003,68(8):2473-2478.
Characteristics of histology and protein in adductor of M anila clam Ruditapes philippinarum
ZHANG Long,LIU Jun-rong,TIAN Yuan-yong,JIANG Meng-yun,LIU Hui-hui, ZHENG Yao,SONG Yang,LIU Jin-yang
(College of Food Science and Engineering,Dalian Ocean University,Dalian 116023,China)
In this study,the histomorphology,protein profile and myofibrillar(Mf)gelation at low temperature were analyzed in muscle of Manila clam Ruditapes philippinarum for exploring the relationship between processing characteristics and properties ofmuscle protein.The results showed that adductormuscle fiberswere highly homon-ymous in direction,aswell as,striated and smoothmusclewere linear and helical columnar,respectively.Analysis of protein distribution in two types ofmuscle by SDS-PAGE electrophoresis revealed that the sarcoplasmic protein was focused on 23 000 presented in striated muscle,while the thick filamentwas dissolved.Compared with striated muscle,more level ofmyorod was only found in smooth muscle,in which there was higher content of paramyosin, and lower content ofmyosin and tropomyosin,an approximate content of actin.The solubility of Mf from whole ad-ductor was depended on ionic strength and ATP.Themaximum value of Ca2+-ATPase activity was observed in the middle ionic strength(KCl concentration 0.30 mol/L),and the activity was reduced gradually as the deviation from themiddle ionic strength.Proteins involved in crosslinking process contained myosin,paramyosin and tropo-myosin,with the increase in ionic strength,themore significanteffectof crosslinking.The findings provide a scien-tific reference for the further study on the protein properties of edible parts from economically important shellfish.
Ruditapes philippinarum;striated muscle;smooth muscle;myofibrillar protein;protein profile
S912
A
10.16535/j.cnki.dlhyxb.2017.03.016
2095-1388(2017)03-0356-07
2016-06-14
国家自然科学基金面上项目(041314030)
张龙(1989—),男,硕士研究生。E-mail:shipinzl@126.com
刘俊荣(1963—),女,博士,教授。E-mail:ljunrong@dlou.edu.cn
猜你喜欢
食品工业科技(2022年17期)2022-08-27
现代食品科技(2022年4期)2022-04-28
智慧农业导刊(2022年6期)2022-04-16
新农业(2022年6期)2022-04-13
水产科学(2021年4期)2021-07-21
食品工业科技(2021年3期)2021-06-16
食品科学(2020年11期)2020-07-13
郑州大学学报(医学版)(2015年2期)2015-02-27
心血管病学进展(2015年4期)2015-02-22
河北渔业(2014年5期)2014-07-08
