
低磷胁迫对不同基因型棉花苗期根系形态及生理特性的影响
2018-02-27 12:18孙淼李鹏程郑苍松刘帅刘爱忠韩慧敏刘敬然董合林
棉花学报 2018年1期
孙淼 ,李鹏程 ,郑苍松 ,刘帅 ,刘爱忠 ,韩慧敏 ,刘敬然 ,董合林 *
(1.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳455000;2.江苏师范大学,江苏徐州221000)
磷是作物体内许多重要化合物的组成成分,在多种代谢过程中起着重要作用,对作物产量和品质均有重要影响[1]。随着现代农业的发展,缺磷耕地面积不断增加,我国有1.07×108hm2耕地,缺磷面积接近0.7×108hm2,其中约有30%耕地面积土壤有效磷含量仅3~5 mg·kg-1[2]。由于土壤中有效磷含量低,对农作物的供应不足,使得施用磷肥成为维持农作物高产的重要措施[3]。但磷肥施入土壤后大部分容易被土壤固定,移动性差,导致磷肥利用率一般只有15%~25%[4]。在农业生产上,土壤中总磷含量高,但能被农作物吸收利用的有效磷含量低,被称为“遗传学缺磷”[5]。长时间增施磷肥,土壤中磷素大量累积,肥料当季利用率持续降低,造成农业生产成本持续增加,已经阻碍农业高效健康发展。因此,研究作物适应土壤低磷和高效吸收利用土壤磷机制,利用作物耐低磷和高效吸收利用磷的基因资源,提升作物对土壤磷的吸收利用率,使土壤中的潜在磷库转化为磷矿替代资源,可以降低作物生产中对磷肥的使用,促进农业可持续发展[6]。
棉花是我国重要的经济作物、大宗农产品和纺织工业原料,在国民经济中的地位举足轻重。磷是棉花生长发育必须的三大营养元素之一,在生育中期能够促进棉花现蕾开花,生育后期促进棉籽成熟,增加铃重,提早吐絮,直接影响籽棉产量和纤维品质。增施磷肥在75~150 kg·hm-2范围内对棉花各器官(除根外)干物质累积和产量有明显促进作用,但继续增加对干物质质量影响不大,甚至有所下降[7]。针对不同叶龄的棉花进行缺磷处理,棉花株高和主茎倒四叶叶面积较正常株分别降低20.8%和32.5%[8]。低磷胁迫下,根系形态的适应性变化是作物高效利用土壤磷的重要生物学基础。已在小麦[9]、玉米[10]、水稻[11]上发现其总根长、根总表面积和侧根长及侧根数在低磷胁迫下均有不同程度增加。当低磷胁迫发生时,不同作物都会形成一套适应机制来应对逆境,关于棉花对低磷胁迫响应的研究尚不多见。因此,本研究选用2种基因型棉花,在不同磷水平下,研究棉花磷吸收利用效率、叶片光合特性和根系形态特征对低磷胁迫的响应,为进一步研究棉花耐低磷胁迫机制提供科学依据。
1 材料与方法
1.1 试验材料和试验设计
选用中棉所79(CCRI 79)和鲁棉研28号(SCRC 28)为试验品种。水培试验于2015年5月在中国农业科学院棉花研究所光照培养室进行,室内昼夜温度分别控制在(28±2)℃和(20±2)℃。选择饱满且胚完整的棉花种子,用2%H2O2消毒30 m in后再清水洗净,在培养箱内进行沙培,待子叶全部展平,洗净沙子后挑选生长一致的棉花,移入装有8L 1/2 Hoagland营养液的避光水培箱(长 32 cm、宽 25 cm、高 14 cm),每天供氧 24 h,7 d后换至全营养液中,分别进行低磷P0(KH2PO41.0×10-5mol·L-1) 和适磷 P1(KH2PO40.5 mol·L-1)处理,每隔 7 d 换 1 次营养液,营养液以霍格兰营养液和阿农微量元素营养液为基液,每个处理设3个重复,每个水培箱定植15株棉花,生长至五叶期取样。
1.2 样品测定和方法
植株全磷测定:将采集的棉花分为根、茎和叶3个部位,于105℃下杀青30 m in,70℃烘干至恒重后称取干物质质量。样品通过H2SO4-H2O2消煮后,采用钼锑抗比色法[12],最后用分光光度计测定。
光合参数测定:利用便携式光合仪LI-6400测定棉花第4片叶光合特性,得到净光合速率Pn、蒸腾速率 Tr、气孔导度 Gs、胞间 CO2浓度 Ci等相关参数。
根系形态扫描与分析:以子叶节为界将棉株分为两部分,用ScanMaker 9800XL plus扫描根系,将扫描的根系图片用Win RHIZO软件分析,得到总根长、根总表面积和根总体积等参数。
根膜透性:采用电导仪法测定根膜透性[13]。表4中液Ⅰ为每克根在去离子水中放置1 h后溶液的电导率,它可以反映根释放出的电解质的量;液Ⅱ为单位质量根在去离子水中煮后放置1小时后溶液的电导率,它可以反映根中电解质总量。前二者比值可以反映根膜透性的大小。
磷外渗率:将棉花移栽于0.5 mol·L-1CaCl2溶液,4 h后测定单位质量根(鲜物质质量)每小时释放出的磷量[13]。
1.3 数据处理与分析
棉花磷吸收效率=磷含量×棉花干物质质量;
棉花磷利用效率=棉株干物质质量/全株磷吸收量;
相对生物量=(低磷处理棉株干物质质量/适磷处理棉株干物质质量)×100%;
相对吸磷量=(低磷处理棉株磷吸收量/适磷处理棉株磷吸收量)×100%;
试验数据用MS Excel 2003进行数据分析,SPSS 17.0软件进行统计分析。
2 结果与分析
2.1 低磷胁迫对不同基因型棉花干物质质量的影响
由表1可知,低磷处理时,中棉所79和鲁棉研28号单株茎、叶和全株干物质质量均有不同程度减少,仅鲁棉研28号单株根干物质质量增加2.53 mg。中棉所79单株根、茎、叶和全株干物质质量分别减少 4.20 mg、13.33 mg、18.73 mg 和36.27 mg,相对生物量分别为81.52%,46.95%,63.22%和63.29%;鲁棉研28号单株茎、叶和全株干物质质量分别减少 6.65 mg、8.97 mg和13.08 mg,相对生物量分别为64.09%,81.23%和85.32%。低磷处理造成2个基因型棉花各器官干物质质量均有不同程度减少,但地上部干物质质量减少幅度明显大于根系,造成根冠比显著高于适磷处理。说明低磷胁迫降低棉花干物质质量,尤其是中棉所79单株茎、叶和全株干物质质量均显著降低,鲁棉研28号仅单株茎干物质质量显著降低,但2个基因型棉花品种的根冠比均显著增加。
2.2 低磷胁迫对不同基因型棉花磷吸收、分配和利用的影响
由表2可知,低磷处理时,中棉所79和鲁棉研28号根、茎、叶和棉株磷含量均显著下降。在相同磷处理时,鲁棉研28号茎、叶磷含量均显著高于中棉所79。表2还说明,低磷处理时,中棉所79和鲁棉研28号根、茎、叶磷吸收效率均显著下降。在相同磷处理时,2个基因型棉花各器官之间磷吸收效率并无显著差异,但鲁棉研28号根、茎、叶和棉株相对吸磷量较中棉所79分别高23.3、11.9、5.3和9.3百分点 (依实际数据计算)。在低磷处理和适磷处理下的根系吸磷量占棉株总吸磷量比例,中棉所79分别为22.95%和19.53%,鲁棉研28号分别为29.98%和18.00%(依实际数据计算),说明低磷胁迫时,鲁棉研28号将更多的磷分配到根系。在低磷处理和适磷处理下的磷利用效率,鲁棉研28号分别为70.56 mg·mg-1和 20.98 mg·mg-1,中棉所 79 均高于鲁棉研 28 号,分别为 90.92 mg·mg-1和 23.09 mg·mg-1,表明中棉所79有较高的磷利用效率,同时,2个基因型低磷处理的磷利用效率均显著高于适磷处理。
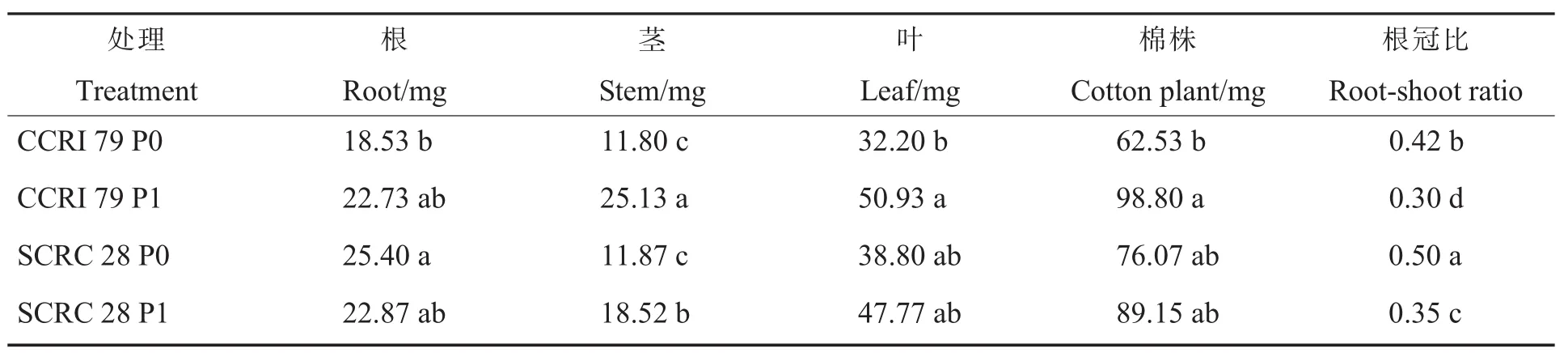
表1 低磷条件下不同基因型棉花的单株干物质质量及根冠比Table 1 Effects of low phosphorus on dry weight and root-shoot ratio of different cotton genotypes
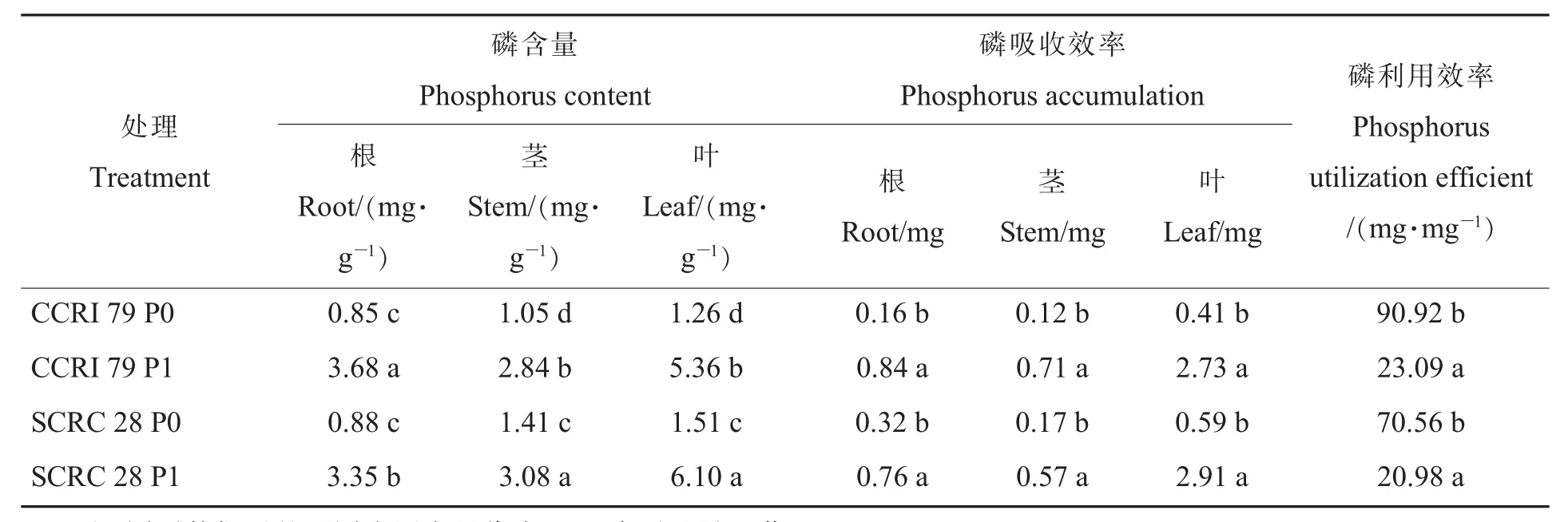
表2 低磷条件下不同基因型棉花对磷的吸收利用Table 2 Effects of low phosphorus on phosphorus accumulation and utilization of different cotton genotypes
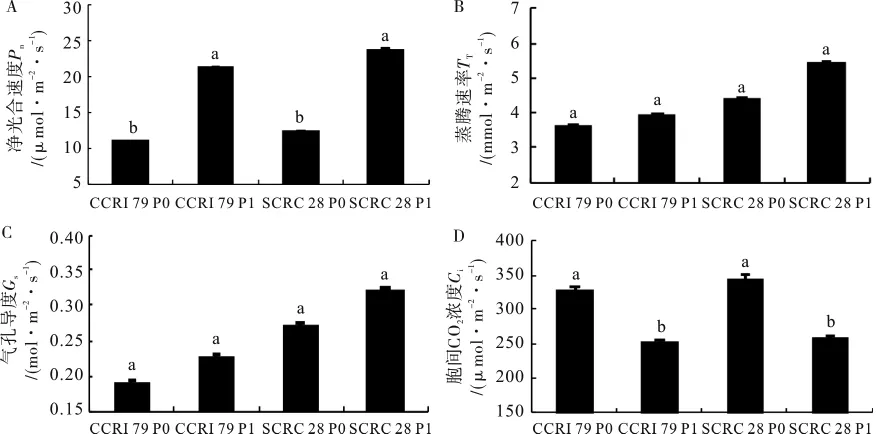
图1 低磷对不同基因型棉花光合特性参数的影响Fig.1 Effects of low phosphorus on photosynthetic characteristic param eter of different cotton genotypes.
2.3 低磷胁迫对不同基因型棉花叶片光合特性的影响
2.3.1 净光合速率。由图1A可知,低磷处理时,中棉所79和鲁棉研28号净光合速率均显著降低,分别减少49.38%和48.68%。鲁棉研28号比中棉所79高13.33%。说明磷营养不足不利于棉花进行光合作用,且不同基因型间有着不同的适应性,本试验中鲁棉研28号表现出较强的耐低磷适应性。
2.3.2 蒸腾速率。蒸腾作用可以降低植物叶面温度,加强对矿质元素的吸收[14]。由图1B可知,低磷处理时,两个品种的蒸腾速率降低,分别减少8.15%和18.95%,但差异均不显著。本试验中,2个基因型通过减少蒸腾速率抑制失水,是棉花适应低磷胁迫的1种自我保护措施。
2.3.3 气孔导度和胞间CO2浓度。气孔作为水分和CO2进出叶片的通道,对光合作用具有重要调节作用。由图1C和1D可知,低磷处理时,2个品种叶片气孔导度分别减少16.81%和15.63%,均未达到显著差异;而胞间CO2浓度分别增加30.06%和33.85%,均差异显著。说明本试验中,低磷胁迫下2个基因型棉花净光合速率下降的主要原因是非气孔因素。
2.4 低磷胁迫对不同基因型棉花根系形态学特征的影响
由表3可知,低磷处理时,中棉所79总根长、根总表面积和根总体积分别降低7.92%,27.97%和26.72%,而鲁棉研28号分别增加13.05%、18.78%和10.50%;鲁棉研28号总根长、根总表面积和根总体积分别比中棉所79高19.83%、41.90%和57.01%。说明磷水平对棉花根系形态学特征有较大影响,且不同基因型棉花的根系形态学特征对低磷胁迫的响应不一致。
2.5 低磷胁迫对不同基因型棉花根膜透性和磷外渗率的影响
由表4可知,在低磷处理和适磷处理下,鲁棉研28号电解质总量 (液Ⅱ)分别比中棉所79高6.15%和9.36%,根膜透性分别低35.87%和18.19%。低磷处理时,中棉所79和鲁棉研28号根膜透性分别降低19.2%和38.38%,磷外渗率表现出相同趋势,分别降低51.08%和57.18%。说明低磷胁迫时,2个基因型棉花根膜透性的变化幅度大于总电解质量的变化幅度,均通过降低根膜透性,减少磷外渗来适应外界缺磷环境,同时,鲁棉研28号根膜透性和磷外渗率均低于中棉所79。
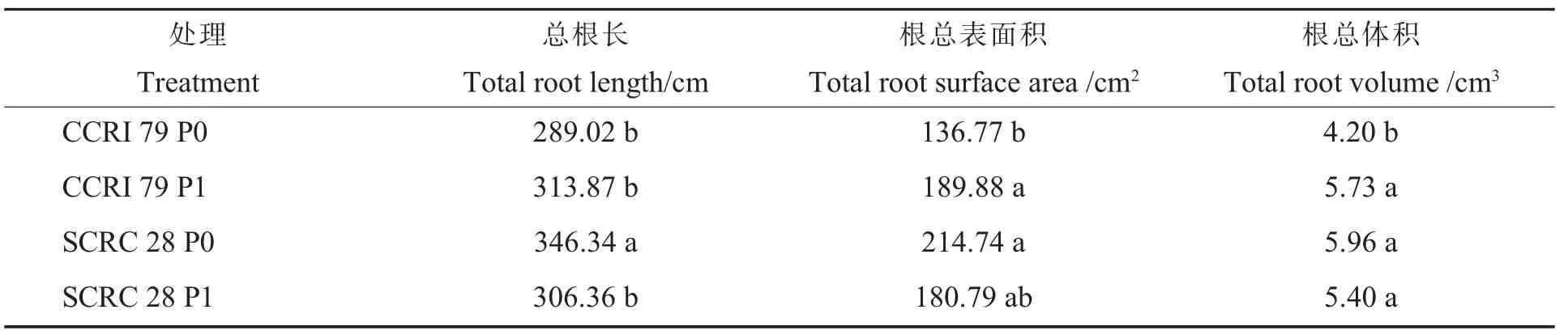
表3 低磷对不同基因型棉花根系形态学参数的影响Table 3 Effects of low phosphorus on root m orphological parameter of different cotton genotypes
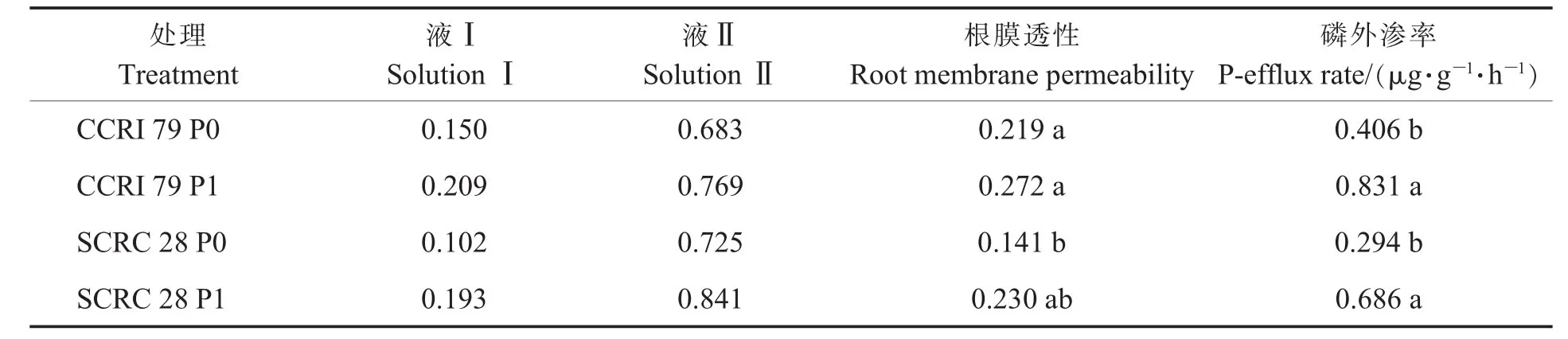
表4 低磷对不同基因型棉花根系生理的影响Table 4 Effects of low phosphorus on root physiology of different cotton genotypes
3 讨论与结论
3.1 低磷胁迫对棉花干物质质量的影响
在棉花整个生育期,干物质的累积以养分吸收为基础,反映了养分吸收利用状况,与棉花产量高低和品质优劣直接相关。本试验结果表明,在低磷胁迫下,棉花单株总干物质质量显著降低[15],与前人关于甜菜[13]和大豆[16]的研究结论一致。有研究表明,棉花各器官干物质质量在低磷胁迫下均呈下降趋势[7]。本研究在棉花不同器官干物质质量对低磷胁迫响应的分析中发现,中棉所79和鲁棉研28号单株茎和叶干物质质量均呈下降趋势,中棉所79单株根干物质质量也呈下降趋势,从0.23 g降到0.19 g,而鲁棉研28号单株根干物质质量从0.23 g增加到0.25 g,表明不同基因型棉花单株根干物质质量对低磷胁迫的响应并不一致。在水培条件下,新陆早13号苗期低磷处理根干物质质量大于适磷处理[6],与鲁棉研28号单株根干物质量的表现一致。低磷胁迫下,不同基因型棉花单株根干物质质量表现不一致,可能是棉花不同基因型间磷效率不一样,也可能由于试验条件(水培、沙培和大田)和取样时期不一样引起,其原因有待进一步研究。本试验研究说明,低磷胁迫显著降低棉花苗期单株茎和全株干物质质量,虽然中棉所79和鲁棉研28号单株根干物质质量表现出不同的变化趋势,但二者的根冠比显著上升,是作物耐低磷胁迫的机制之一,与前人的研究结论一致[17-18]。
3.2 低磷胁迫对棉花磷吸收利用率的影响
已有研究表明,盆栽试验中随着施磷量增加棉花茎叶、根和棉株吸磷量均增加,当施磷量达到960 mg·kg-1时棉花生长受阻且吸磷量呈减少趋势[19]。本研究显示,低磷处理下中棉所79和鲁棉研28号根、茎、叶和棉株磷含量、磷吸收量均显著低于适磷处理,证明磷浓度与棉花磷吸收紧密相关,与前人的研究结论一致[20]。在低磷处理时,鲁棉研28号根、茎、叶和棉株磷含量和磷吸收量均高于中棉所79,但鲁棉研28号磷利用率仅为 70.56 mg·mg-1, 低于中棉所 79的 90.92 mg·mg-1。在棉株不同器官磷分配的分析中发现,中棉所79和鲁棉研28号地上部磷百分数分别为77.05%和70.02%。表明低磷胁迫时,中棉所79将更多的磷分配到地上部分,以此来维持地上部磷的代谢作用,这或许是中棉所79拥有较高磷利用效率的原因之一[6]。本试验研究说明,低磷胁迫显著降低棉花苗期根、茎、叶和棉株磷含量、磷吸收量,同时鲁棉研28号仅拥有较高磷吸收效率,而中棉所79拥有较高磷利用效率。
3.3 低磷胁迫对棉花叶片光合特性的影响
磷是叶绿体及其构成(双层膜、基粒、DNA)和ATP的组成成分,在光合作用物质转化中起重要作用。许多研究证实,包括低磷胁迫在内的诸多逆境胁迫,均会导致光合速率降低[21-22],气孔张开度变小使得气孔导度降低,同时也降低了蒸腾失水[14],这应该是作物面对逆境的1种自我保护机制。本研究中,低磷胁迫下,中棉所79和鲁棉研28号的净光合速率、蒸腾速率和气孔导度均有不同程度降低,与前人研究结论一致;而胞间CO2浓度均显著上升,对此以往的研究并没有一致的结论,甘蓝型油菜[23]和大豆幼苗[24]在低磷处理时胞间CO2浓度均呈下降趋势;熊猫豆[25]和棉花[26]分别在低磷处理和花铃期增温处理时均呈上升趋势;不同棉花品种幼苗在低温胁迫下表现不一致,有上升有降低。导致净光合速率下降的原因包括气孔因素和非气孔因素[27],Jacob[28]研究低磷胁迫对大豆光合作用的影响中发现低磷胁迫导致光合速率降低,并观察到气孔导度降低和叶肉细胞变小,但光合速率下降幅度大于气孔导度的下降幅度,同时胞间CO2浓度增加,与本研究结论相同,说明低磷胁迫显著降低棉花苗期净光合速率,且非气孔因素是光合作用下降的主要原因[29-30]。
3.4 低磷胁迫对棉花根系形态和生理特性的影响
作物在低磷胁迫下经常发生形态学上的变化,最明显的变化就是根冠比增加,根长增加、侧根及根毛增多,扩大根系覆盖面积,以此增加磷吸收来适应低磷环境[31-32]。在本试验中,低磷胁迫下,鲁棉研28号总根长、根总表面积和根总体积均增加,尤其是总根长显著增加,但中棉所79总根长、根总表面积和根总体积均降低。陈波浪等[6]研究低磷胁迫时,新海18号总根长、根总表面积和根总体积分别增加36.0%、145.7%和96.9%,新陆早13号总根长和根总表面积则显著下降,新海18号根系各参数均高于新陆早13号。究其原因,应该是同种作物不同品种根系对低磷胁迫的响应存在特异性[33]。本试验研究说明低磷胁迫时,鲁棉研28号一方面通过增加根长和根表面积增加磷吸收,另一方面通过降低根膜透性减少磷外渗率,能够更有效地满足自身对磷营养的需求,同时鲁棉研28号拥有较好的根系形态学参数也印证了前文中其根干物质质量高于适磷处理和磷吸收效率高于中棉所79的结论,表明鲁棉研28号拥有较强的耐低磷胁迫能力。
[1]宋世佳,孙红春,张永江,等.水培磷胁迫下不同基因型棉花苗期根系形态及叶片光合特性的差异[J].棉花学报,2015,27(3):223-231.Song Shijia,Sun Hongchun,Zhang Yongjiang,et al.Genotypic differences in root morphology and leaf photosynthesis of cotton seedlings treated with phosphorus deficient solutions[J].Cot-ton Science,2015,27(3):223-231.
[2]李卫华.棉花磷素高效利用品种筛选及机理的初步研究[D].乌鲁木齐:新疆农业大学,2010:1-2.Li Weihua.Research on phosphorus efficient genotypes of cotton varieties screening and its mechanism[D].Urumqi:Xinjiang Agricultural University,2010:1-2.
[3]陆欣春,韩晓增,邹文秀.作物高效利用土壤磷素的研究进展[J].土壤与作物,2013,2(4):164-172.Lu Xinchun,Han Xiaozeng,Zou Wenxiu.Advance in the efficient utilization of soil phosphorus by crops[J].Soil and Crop,2013,2(4):164-172.
[4]李志伟,崔力拓.大量施磷对旱地土壤养分、酶活性及作物生长的影响[J].土壤通报,2009,40(4):860-863.Li Zhiwei,Cui Lituo.The effect of applying surplus phosphorus on the soil nutrient enzyme activity and crop grow th of nonirrigated farm land[J].Chinese Journal of Soil Science,2009,40(4):860-863.
[5]张丽梅,贺立源,李建生,等.不同耐低磷基因型玉米磷营养特性研究[J].中国农业科学,2005,38(1):110-115.Zhang Limei,He Liyuan,Li Jiansheng,et al.Phosphorus nutrient characteristics of different maize inbreds with tolerance to low-P stress[J].Scientia Agricultura Sinica,2005,38(1):110-115.
[6]陈波浪,盛建东,蒋平安,等.不同基因型棉花磷效率特征及其根系形态的差异[J].棉花学报,2014,26(6):506-512.Chen Bolang,Sheng Jiandong,Jiang Ping′an,et al.Differences of phosphorus efficiency characteristics and root morphology between two cotton genotypes[J].Cotton Science,2014,26(6):506-512.
[7]戴婷婷,盛建东,陈波浪.磷肥不同用量对棉花干物质及氮磷钾吸收分配的影响[J].棉花学报,2010,22(5):466-470.Dai Tingting,Sheng Jiandong,Chen Bolang.Effect of different phosphorus fertilizer rate on dry matter accumulation and the absorption and distribution of nitrogen,phosphorous,potassium of cotton[J].Cotton Science,2010,22(5):466-470.
[8]陈波浪,盛建东,贾宏涛,等.不同叶龄期氮磷钾缺失对棉花生长及养分积累的影响[J].新疆农业科学,2013,50(2):342-347.Chen Bolang,Sheng Jiandong,Jia Hongtao,et al.Effect of N,P and K elements deficiency on grow th and nutrition accumulation of cotton during different leaf ages[J].Xinjiang Agricultural Sciences,2013,50(2):342-347.
[9]任永哲.低磷胁迫对不同基因型小麦品种苗期性状的影响[J].中国农学通报,2012,28(18):40-44.Ren Yongzhe.Effects of low phosphorus stress on different wheat varieties at seedling stage[J].Chinese Agricultural Science Bulletin,2012,28(18):40-44.
[10]刘存辉,张可炜,张举仁,等.低磷胁迫下磷高效玉米单交种的形态生理特性[J].植物营养与肥料学报,2006,12(3):327-333.Liu Cunhui,Zhang Kewei,Zhang Juren,et al.Morphological and physiological characteristics of P-efficient maize hybrid to stress of phosphorus deficiency[J].Plant Nutrition and Fertilizer Science,2006,12(3):327-333.
[11]明凤,娄玉霞,梁斌,等.低磷胁迫下不同水稻品种根系吸收能力差异的生理与遗传本质[J].应用与环境生物学报,2002,8(5):473-477.M ing Feng,Lou Yuxia,Liang Bin,et al.Genetic and physiological mechanism of P-uptake ability difference between different rice(Oryza sativa L.)genotypes under low-P stress[J].Chinese Journal of Applied and Environmental Biology,2002,8(5):473-477.
[12]鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.Bao Shidan.Analysis of soil and agricultural chemistry[M].Beijing:China Agricultural Press,2000.
[13]刘慧,刘景福,刘武定.不同磷营养油菜品种根系形态及生理特性差异研究[J].植物营养与肥料学报,1999,5(1):40-45.Liu Hui,Liu Jingfu,LiuWuding.Differences of root morphology and physiological characteristics between two rape genotypes with different P-efficiency[J].Journal of Plant Nutrition and Fertilizer,1999,5(1):40-45.
[14]刘菲菲,魏亦农,李志博,等.低温胁迫对棉花幼苗叶片光合特性的影响[J].石河子大学学报(自然科学版),2011,29(1):11-14.Liu Feifei,Wei Yinong,Li Zhibo,et al.Photosynthesis of cotton seedlings under low temperature stress[J].Journal of Shihezi U-niversity(Natural Science),29(1):11-14.
[15]王靓,盛建东,罗佳,等.不同基因型棉花磷效率的差异特征[J].中国土壤与肥料,2015(3):44-49.Wang Liang,Sheng Jiandong,Luo Jia,et al.Difference characteristics of phosphorus efficiency for genotypes cotton[J].Soil and Fertilizer Sciences in China,2015(3):44-49.
[16]乔振江,蔡昆争,骆世明.低磷和干旱胁迫对大豆植株干物质积累及磷效率的影响[J].生态学报,2011,31(19):5578-5587.Qiao Zhenjiang,Cai Kunzeng,Luo Shim ing.Interactive effects of low phosphorus and drought stress on dry matter accumulation and phosphorus efficiency of soybean plants[J].Acta E-cologica Sinica,2011,31(19):5578-5587.
[17]王刚,郑苍松,李鹏程,等.土壤有效磷含量对棉花幼苗干物质积累和碳氮代谢的影响[J].棉花学报,2016,28(6):609-618.Wang Gang,Zheng Cangsong,Li Pengcheng,et al.Effects of soil Olsen-P levels on the dry matter accumulation and carbon and nitrogen metabolism of cotton plant at seedling stage[J].Cotton Science,2016,28(6):609-618.
[18]詹书侠,郑淑霞,王扬,等.羊草的地上―地下功能性状对氮磷施肥梯度的响应及关联[J].植物生态学报,2016,40(1):36-47.Zhan Shuxia,Zheng Shuxia,Wand Yang,et al.Response and correlation of above-and below-ground functional traits of Leymus chinensis to nitrogen and phosphorus additions[J].Chinese Journal of Plant Ecology,2016,40(1):36-47.
[19]崔水利,张炎,王讲利,等.施磷对棉花根系形态及其对磷吸收的影响[J].植物营养与肥料学报,1999,3(3):249-254.Cui Shuili,Zhang Yan,Wang Jiangli,et al.Effect of phosphorus placed in soil on root system morphology of cotton and P uptake[J].Plant Nutrition and Fertilizer Science,1999,3(3):249-254.
[20]曹立为,郭晓双,龚振平,等.磷素营养变化对大豆磷素积累及产量和品质的影响[J].大豆科学,2015,34(3):458-462.Cao Liwei,Guo Xiaoshuang,Gong Zhenping,et al.Changes of phosphorus nutrition on P accumulation,yield and quality of soybean[J].Soybean Science,2015,34(3):458-462.
[21]吴楚,范志强,王政权.磷胁迫对水曲柳幼苗叶绿素合成、光合作用和生物量分配格局的影响[J].应用生态学报,2004,(6):935-940.Wu Chu,Fan Zhiqiang,Wang Zhengquan.Effect of phosphorus stress on chlorophyll biosynthesis,photosynthesis and biomass partitioning pattern of Fraxinus mandchurica seedlings[J].Chinese Journal of Applied Ecology,2004,(6):935-940.
[22]朱隆静,喻景权.不同供磷水平对番茄生长和光合作用的影响[J].浙江农业学报2005,17(3):120-122.Zhu Longjing,Yu Jingquan.Effects of different phosphate levels on grow th and photosynthesis of tomato[J].Acta Agriculturae Zhejiangensis,2005,17(3):120-122.
[23]张海伟,徐芳森.不同磷水平下甘蓝型油菜光合特性的基因型差异研究[J].植物营养与肥料学报,2010,16(5):1196-1202.Zhang Haiwei,Xu Fangsen.Genotypic differences in photosynthetic characteristics in Brassica napus at different phosphorus levels[J].Plant Nutrition and Fertilizer Science,2010,16(5):1196-1202.
[24]原向阳,郭平毅,黄洁,等.缺磷胁迫下草甘膦对抗草甘膦大豆幼苗光合作用和叶绿素荧光参数的影响[J].植物营养与肥料学报,2014,20(1):221-228.Yuan Xiangyang,Guo Pingyi,Huang Jie,et al.Effect of glyphosate on photosynthesis and chlorophyll fluorescence of leaves of glyphosate-resistant soybean[Glycine max(L.)Merr.]seedlings under phosphorus deficiency stress[J].Journal of Plant Nutrition and Fertilizer,2014,20(1):221-228.
[25]廉满红,田宵鸿,曹翠玲.低磷条件下熊猫豆光合特性及碳水化合物累积变化研究[J].干旱地区农业研究,2011,29(5):87-99.Lian Manhong,Tian Xiaohong,Cao Cuiling.Effect of phosphorous stress on photosynthesis characteristics and distribution of carbohydrate of Phaseolus coccineus L.seedlings[J].Agricultural Research in the Arid Areas,2011,29(5):87-99.
[26]孙啸震,张黎妮,戴艳娇,等.花铃期增温对棉花干物重累积的影响及其生理机制[J].作物学报,2012,38(4):683-690.Sun Xiaozhen,Zhang Lini,Dai Yanjiao,et al.Effect of increased canopy temperature on cotton plant dry matter accumulation and its physiological mechanism[J].Acta Agronomica Sinica,2012,38(4):683-690.
[27]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review Plant Physiology,1982(33):317-345.
[28]Jacob J,Law lor D W.Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower,maize and wheat plants[J].Journal of Experimental Botany,1991,42(241):1003-1011.
[29]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.Xu Daquan.Some problems in stomatal limitation analysis of photosynthesis[J].Plant Physiology Communications,1997,33(4):241-244.
[30]Battie-Laclau P,Laclau J P,Beri C,et al.Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment[J].Plant,Cell and Environment,2014,37(1):70-81.
[31]罗佳,候银莹,程军回,等.低磷胁迫下不同磷效率基因型棉花的根系形态特征[J].中国农业科学,2016,49(12):2280-2289.Luo Jia,Hou Yinying,Cheng Junhui,et al.Root morphological characteristics of cotton genotypes with different phosphorus efficiency under phosphorus stress[J].Scientia Agricultura Sinica,2016,49(12):2280-2289.
[32]杨青,张一,周志春,等.低磷胁迫下不同种源马尾松的根构型与磷效率[J].应用生态学报,2012,23(9):2339-2345.Yang Qing,Zhang Yi,Zhou Zhichun,et al.Root architecture and phosphorus efficiency of different provenance Pinus masso-niana under low phosphorous stress[J].Chinese Journal of Applied Ecology,2012,23(9):2339-2345.
[33]高家合,邓碧儿,曾秀成,等.烟草磷效率的基因型差异及其与根系形态构型的关系[J].西北植物学报,2010,30(8):1606-1613.Gao Jiahe,Deng Bi’er,Zeng Xiucheng,et al.Genotypic variation in phosphorus efficiency of tobacco in relation to root morphological characteristics and root architecture[J].Acta Botanica Boreali-Occidentalia Sinica,2010,30(8):1606-1613.
猜你喜欢
南方农业(2022年17期)2022-10-21
农业科技通讯(2022年7期)2022-07-19
安徽农学通报(2022年6期)2022-04-07
农村百事通(2020年9期)2020-05-29
新疆农业科学(2020年4期)2020-04-07
河南农业·综合版(2019年9期)2019-09-10
广西农学报(2019年1期)2019-07-18
浙江农业科学(2019年1期)2019-01-28
中国棉花(2018年3期)2018-04-09
现代农村科技(2018年8期)2018-01-18
