
棉属AD远缘杂种的合成及性状鉴定
2018-04-18 07:10吴玉香李昱樱申状状张磊郑赟
山西农业大学学报(自然科学版) 2018年4期
吴玉香,李昱樱,申状状,张磊,郑赟
(山西农业大学 农学院,山西 太谷 030801)
棉花属双子叶植物(Dicotyledons)、锦葵科(Malvaceae)、棉属(Gossypium)。棉属有51个种,其中46个为二倍体棉种(2n=2X=26),5个为异源四倍体棉种(2n=4X=52)[1]。多年来对棉属四倍体种起源及其供体种问题上,普遍一致的看法是草棉和亚洲棉是四倍体棉种A基因组的祖先供体种,雷蒙德氏棉是D基因组的祖先供体种[2]。这个基因组在棉属进化的早期大约700~1100万年前曾各自经历了单独进化[3,4]。棉属A、D组棉种是研究物种形成过程中生殖隔离机制和基因组互作的良好材料,有关棉属生殖隔离机制的研究对探讨四倍体棉种形成和分化具有重要的科学意义,对棉属远缘杂交亲本选配有重要的参考价值。然而自然条件下由于棉属A、D染色体组棉种之间存在较大的生殖隔离,A组和D组棉种较难杂交成功。
本研究选用A组亚洲棉、草棉为母本,D组雷蒙德氏和瑟伯氏棉为父本进行远缘杂交,旨在获得杂种并从形态学初步鉴定杂种的真实性,探讨A、D组棉种间生殖隔离的先后顺序。进一步探究亲本基因组在杂种中的互作模式,为棉属异源四倍体棉种形成和起源进化提供最直接的证据,为棉花种质创新研究提供有价值的材料。
1 材料和方法
1.1 试验材料
本试验所用材料有:A组草棉(G.herbaceum)(2n=2x=26)、亚洲棉(G.arboreum)(2n=2x=26);D组野生雷蒙德氏棉(G.raimondii)(2n=2x=26)、瑟伯氏棉(G.thurberi)(2n=2x=26)。野生棉材料来源于国家种质三亚野生棉圃,其染色体组及地理分布见表1。

表1 供试材料及其地理分布Table 1 The materials and geographic distribution
1.2 远缘杂交
试验分别以A组亚洲棉和草棉为母本,D组雷蒙德氏棉和瑟伯氏棉为父本进行远缘杂交,共进行了4个组合的杂交,分别是:亚洲棉×雷蒙德氏棉;亚洲棉×瑟伯氏棉;草棉×雷蒙德氏棉;草棉×瑟伯氏棉。每个组合各选500朵母本花进行远缘杂交,具体步骤:母本去雄、父本花粉人工授粉、第二天重复授粉,以保证成功率。亚洲棉花蕾大小为苞叶和花蕾等长且发黄时合适。草棉花蕾变黄,若隐若现时即为合适。
1.3 远缘杂种的形态学观察和鉴定
收获每个组合的杂交种子并播种,以当年长成的杂种植株为材料,获得的远缘杂种从整个植株、叶片、花和父母本做比较,初步从形态学水平观察鉴定杂种的真实性。
2 结果与分析
2.1 远缘杂交结果
对4个组合的远缘杂交结果进行统计,结果见表2。每个组合杂交花数均为500,草棉×雷蒙德氏棉结铃数为11,每个铃有1~3棵种子,一共获得27棵种子,只有3棵饱满种子,播种后全部成苗,结铃率为2.20%,成苗率为11.11%;亚洲棉×雷蒙德氏棉杂交花数为500朵,最终结铃数为43,结铃率为8.60%,获得种子数为34粒,播种后成苗数为4棵,成苗率为11.76%;亚洲棉×瑟伯氏棉杂交花数为500朵,结铃数为28,结铃率为5.60%,获得种子数为36粒,播种后成苗数为3棵,成苗率为8.33%;草棉×瑟伯氏棉杂交花数为500朵,结铃数为4,结铃率为0.80%,没有收到饱满种子,并无一成苗。杂交结果初步表明草瑟组合属于合子前生殖隔离。草雷、亚雷、亚瑟组合生殖隔离是由合子前和合子后生殖隔离共同起作用。
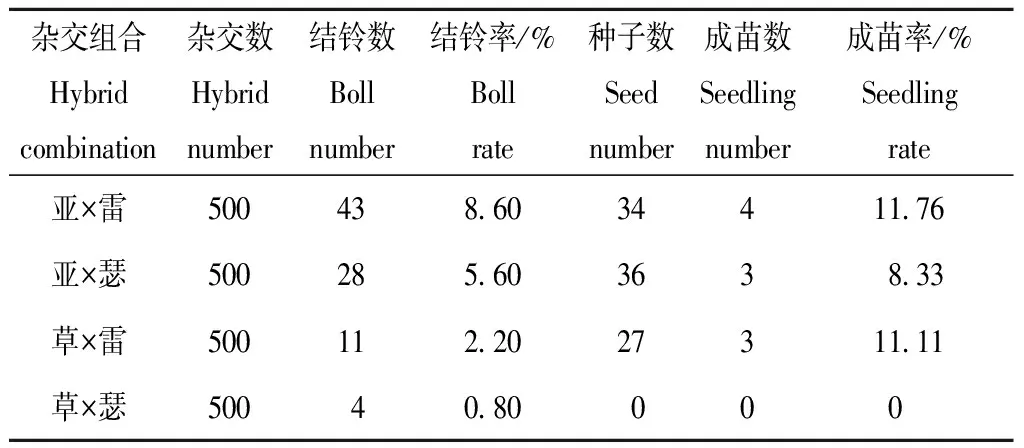
表2 四个组合远缘杂交结果Table 2 The hybrid result of four combination
2.2 草雷杂种形态学观察和鉴定
草雷杂种植株有很强的杂种优势,营养生长旺盛,枝条伸展。播种后45 d现蕾开花,杂种开花时间比父本雷棉早比母本草棉晚,杂种只开花不能结铃。杂种叶形间于双亲之间且比亲本厚,在老的枝条上出现3种叶型:父、母本叶型和中间叶型。杂种花比母本草棉大比父本雷棉小,花冠黄色并有深红色花斑近似于草棉母本;花药黄色近似于草棉母本,但花丝深红像父本雷棉且比母本长。柱头浅黄比草棉长比雷棉短,苞叶联合近似草棉,小苞片数间于双亲之间,花丝平展且存在一丝多药特征,这个特性既不像母本也不像父本,花丝数少于父母本(图1和图2)。

图1 草雷杂种植株以及亲本草棉和雷棉比较Fig.1 Hybrid plant comparison with its parents between G. herbaceum and G. raimondii 注:1:母本草棉,2:杂种,3:父本雷蒙德氏棉Note:1:G. herbaceum 2:Hybrid 3: G. raimondii

图2 草雷杂种花以及亲本草棉和雷棉比较Fig.2 Flower comparison with its parents between G. herbaceum and G. raimondii注:1:母本草棉,2:杂种花,3:父本雷蒙德氏棉Note:1: G. herbaceum 2: Hybrid 3: G. raimondii
草雷叶片密布表皮毛和腺体,有较强抗虫性,近似于雷棉父本。在海南常年进行营养生长和生殖生长,5-10月份主要进行营养生长,但每天都能开花;10-5(第二年)月份开花最多。这是因为母本草棉(来自非洲)抗旱基因的存在,草棉耐热,一年四季开花,加上父本雷棉也是抗旱材料,夏天进行高效率的营养生长。
2.3 亚雷杂种的形态学观察和鉴定
以A组亚洲棉为母本,D组雷蒙德棉为父本进行杂交,获得了杂交种子并已成苗,观察其形态性状:亚雷杂种叶形间于两亲本之间,并且杂种植株呈现三种叶形,两裂片和三裂片以及父本叶形都有,颜色比两亲本深;花瓣大小和花斑大小均间于两亲本之间,花瓣颜色淡黄色像父本,花斑颜色深红趋向于母本亚洲棉,苞叶大小间于父母本之间但形状像父本,花蕊颜色黄色像母本,但花丝深红色且较长像父本,雌蕊柱头淡黄色间于两亲本之间,比母本长比父本短(图3和图4)。

图3 亚雷杂种植株和父母本比较Fig.3 Hybrid plant comparison with its parents between G. arboreum and G. raimondii注:1:母本亚洲棉(左),杂种(右),2:父本雷蒙德氏棉Note:1: G. arboreum (left), Hybrid (right) 2: G. raimondii

图4 亚雷杂种花和父母本比较Fig.4 Flower comparison with its parents between G. arboreum and G. raimondii注:1:母本亚洲棉,2:杂种花,3:父本雷蒙德氏棉Note:1: G. arboreum 2: Hybrid 3: G. raimondii
2.4 亚瑟杂种的形态学观察和鉴定
通过观察亚瑟杂种植株及叶片性状(图5):亚瑟杂种植株生长受阻,植株矮小,但能长侧枝,比父母本都低,杂种植株不现蕾,不开花,不结实,叶形间于双亲之间。另外,杂种的叶脉处有明显的皱缩和不平整现象,可能是因为杂种基因组互作的不协调导致植株出现异常。亚瑟杂种易受高温影响,在海南4-9月生长基本受阻,因为父本瑟伯氏棉的不耐热基因或抗寒基因的存在。杂种长势还在继续观察中,希望采取一些管理措施如喷施叶面肥和激素或嫁接繁殖等恢复其生长。

图5 亚瑟杂种植株以及亲本亚洲棉和瑟伯氏棉Fig.5 Hybrid plant comparison with its parents between G. arboreum and G. thurberi注:1:母本亚洲棉,2:杂种,3:父本瑟伯氏棉Note:1: G. arboreum 2: Hybrid 3: G. thurberi
3 讨论
3.1 远缘杂交生殖隔离机制探讨
生殖隔离是指由于各方面的原因,使亲缘关系接近的类群之间在自然条件下不交配,即使能交配也不能产生后代或不能产生可育后代的隔离机制。生殖隔离最早被划分为合子前隔离和合子后隔离2种形式[5],若隔离发生在受精以前,就称为受精前的生殖隔离,包括地理隔离、生态隔离、季节隔离、生理隔离、形态隔离和行为隔离等;若隔离发生在受精以后,就称为受精后的生殖隔离,包括杂种不活、杂种不育和杂种衰败等。合子前隔离主要受生态和生殖因素的影响;合子后隔离高度依赖于遗传因素,受亲本间相配程度的影响[6]。影响物种间生殖隔离的先后顺序里,那些阻止杂交的隔离将成为限制基因流发生的第一次机会,因此生态地理隔离在一些分类群的总隔离中起到了更重要的作用,激发研究者更大的动力去探究[7]。然而其他一些研究发现合子后障碍对阻止基因交流也非常重要[8~12],因为生殖隔离也常常表现为环境是否强烈影响杂种的行为,即外在的生殖隔离障碍是否依赖于环境,因为杂种要面对自然条件下的强选择压力才能最终存活。
物种形成是一个复杂过程,其实质是生殖隔离
机制的进化,因此研究物种形成主要是研究各种生殖隔离机制形成和进化的过程。本研究结果表明草×瑟组合生殖隔离完全是由于合子前生殖隔离,草×雷、亚×雷、亚×瑟生殖隔离是由合子前和合子后生殖隔离共同起作用。结果还表明D组雷蒙德氏棉能同时和A1、A2组杂交成功并获得了杂种后代,这也进一步验证了雷蒙德氏棉被视为四倍体棉种祖先的直接证据。
3.2 远缘杂种中亲本基因组互作探讨
对于进行有性生殖的生物来说,杂种新个体生物学上的适应需要父母本基因组成功协调的互作,而这些互作表现在从DNA到外在表型的不同阶段发生。杂交后的基因组互作探讨也将极大的促进物种形成生物学的发展[13]。因此我们期望将两组分化百万年的A、D基因组棉种杂交到F1中产生新的表现型。远缘杂种的基因组互作和协调是杂种稳定和存活的关键,基因组互作能够很好解释种间杂交导致的极端后果如杂种优势和互作不亲和导致的杂种坏死。本研究4个组合远缘杂交,只有3个组合获得杂种植株,杂种大多数性状间于双亲之间。只有草雷、亚雷2个组合杂种植株能现蕾开花,杂种基因组互作亲和且有较强生长势。亚瑟杂种植株矮小,不现蕾不开花,但能长侧枝,究其原因是亲本基因组互作不亲和的结果。转录组测序技术提供了解析来自父母本基因组互作和表达模式的强大信息,来自不同亲本的杂种后代转录组学比较,可以阐明基因组互作在转录亲和中起到的关键作用。本研究获得的3个组合杂种的基因组互作模式有待于进一步从转录表达水平来验证。
4 结论
草瑟属于合子前生殖隔离;草雷、亚雷、亚瑟生殖隔离是由合子前和合子后生殖隔离共同起作用。且草雷、亚雷杂种基因组互作亲和,而亚瑟杂种基因组互作不亲和。D组雷蒙德氏棉能同时和A1、A2组杂交成功并获得杂种后代,进一步验证了雷蒙德氏棉被视为四倍体棉种祖先的直接证据。
[1]Fryxell P A. A revised taxonomic interpretation ofGossypiumL. (Malvaceae)[J],Pheedea,1992,2: 108-165.
[2]Wendel JF, Cronn RC. Polyploidy and the evolutionary history of cotton[J]. Advances in Agronomy, 2003, 78(2):139-186.
[3]Seelanan T, Schnabel A, Wendel JF. Congruence and consensus in the cotton tribe (Malvaceae), Syst Bot, 1997, 22(2):259-290.
[4]Cronn RC, Small RL, Haselkorn T ,et al. Rapid diversification of the cotton genus (Gossypium: Malvaceae) revealed by analysis of sixteen nuclear and chloroplast genes[J]. American journal of botany. 2002, 89(4):707-725.
[5]Snow AA. Postpollination selection and male fitness in plants[J]. Amcrucan naturaliat, 1994, 144 (2): 69-83.
[6]Moyle LC, Olson MS, Tiffin P. Patterns of reproductive isolation in three angiosperm genera[J]. Evolution, 2004, 58 (6): 1195-1208.
[7]Sobel JM, Ecogeographic Isolation and Speciation in the Genus Mimulus[J]. American naturalist, 2014,184(5): 565-579.
[8]Kozak GM, Rudolph AB, Colon BL ,et al. Postzygotic isolation evolves before prezygotic isolation between fresh and salt water populations of the rainwater killifish, Lucania parva[J]. Journal of evolutionary biology, 2012:1-11.
[9]Sambatti JBM, Strasburg JL, Ortiz-Barrientos D, et al. Reconciling extremely strong barriers with high levels of gene exchange in annual sunflowers[J]. Evolution, 2012, 66(5):1459-1473.
[10]Scopece G, Croce A, Lexer C, et al. Components ofreproductive isolation between Orchis mascula and Orchis pauciflora[J]. Evolution, 2013, 67(7): 2083-2093.
[11]Briscoe Runquist RD, Chu E, Iverson JL, et al. Rapid evolution of reproductive isolation between incipient out-crossing and selfing Clarkia species[J]. Evolution, 2014, 68:2885-2900.
[12]Kao JY, Lymer S, Hwang SH, et al. Postmating reproductive barriers contribute to the incipient sexual isolation of the United States and Caribbean Drosophila melanogaster[J]. Evolution, 2015, 5:3171-3182.
[13]Payseur BA, Rieseber LH. A genomic perspective on hybridization and speciation[J] Molecular Ecology, 2016, 25(11): 2337-2360.
猜你喜欢
林业科学研究(2022年4期)2022-08-03
麦类作物学报(2022年5期)2022-05-20
中国农业大学学报(2022年6期)2022-05-16
中国种业(2021年2期)2021-03-01
新世纪智能(英语备考)(2019年12期)2020-01-13
高中生之友(中旬刊)(2019年5期)2019-01-10
种业导刊(2019年5期)2019-01-04
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
上海农业学报(2017年3期)2017-04-10
种子科技(2015年10期)2015-01-22
