
盐旱交互对燕麦种子萌发及幼苗生理特性的影响
2018-05-22 03:14张宇君赵丽丽王普昶陈超
草业学报 2018年5期
张宇君,赵丽丽*,王普昶,陈超
(1.贵州大学动物科学学院草业科学系,贵州 贵阳550025;2.贵州省农业科学院草业研究所,贵州 贵阳 550006)
燕麦(Avenasativa)为禾本科燕麦属一年生粮饲兼用作物,是饲养家畜的优选饲粮,其本身含有较为丰富的膳食纤维、β-葡聚糖、蛋白质、淀粉等,同时具有较高的医用价值,长期食用能明显抑制血糖、血脂和血压的升高[1],还能促进肠道益生菌繁殖,增强人体免疫力,在我国西南地区种植面积较大[2]。西南地区是全球岩溶三大区之一,广泛分布着以石灰土为主的非地带性土壤,其基岩中多为可溶性盐类且极具富钙[3-4],同时由于岩溶地区土层浅薄和岩石的高渗透性使得土壤持水力减弱,加之全球气候变暖、干旱频繁等多方面因素导致干旱和钙盐已成为岩溶地区鲜明的生境特征[5-7],这使得燕麦在该环境下生长时常会遭受钙盐和干旱的双重胁迫,尤其在耐盐性较低的萌发期遭遇干旱少雨的春季返盐阶段后,会直接导致燕麦单产低及总产不稳定的发生[8],这是制约燕麦生产的重要限制性因素之一。在大规模种植生产、粮改饲等工程实施中,燕麦因适应性强常作为重要的经济作物被采用,因此,研究钙盐、干旱交互胁迫下燕麦种子的萌发状况及幼苗的生理生化适应机理,对西南岩溶地区的燕麦生产具有重要理论意义和实际生产价值。
目前有关燕麦的研究主要侧重于单一PEG[9]、NaCl[10]、水分[11]胁迫因子或是盐碱交互胁迫[12-13]、氯化钠与干旱交互胁迫[14]对其生长发育的影响。但针对高钙、干旱岩溶环境下燕麦的适应性研究缺少相关报道。因此,本试验设置了CaCl2、PEG单一胁迫及CaCl2+PEG交互胁迫试验,对钙盐、干旱交互胁迫下燕麦种子萌发及幼苗的生理变化进行系统研究,比较燕麦对单一胁迫与交互胁迫之间的适应差异,分析不同胁迫处理下燕麦种子的萌发参数和幼苗生理指标的变化情况,并结合隶属函数评价各胁迫处理对燕麦的影响,筛选出最佳的胁迫处理浓度。本实验结果可为燕麦在岩溶地区的引种栽培及抗逆育种研究提供理论参考依据。
1 材料与方法
1.1 试验材料
供试材料燕麦Avena-23由贵州大学草业科学饲草育种课题组提供。
1.2 试验方法
试验采用双因素析因设计,针对西南岩溶地区富钙干旱的特点,选取CaCl2作为盐胁迫渗透物质,设置5个胁迫梯度分别为:0(CK)、50、100、150、200 mmol·L-1,标记为A0、A1、A2、A3、A4。利用PEG-6000配成不同水势的高渗溶液模拟干旱,设置6个梯度:0(CK)、-0.2、-0.4、-0.6、-0.8、-1.2 MPa,标记为B0、B1、B2、B3、B4、B5。2个因素胁迫共计30个处理,如表1所示。
根据文献[15]进行发芽试验。选取大小均匀一致、籽粒饱满的燕麦种子,用0.5% KMnO4溶液浸种3 min消毒后用蒸馏水冲洗3遍,最后用滤纸吸干种子表面水分,置于直径为15 cm、垫有双层滤纸的培养皿中,每培养皿100粒种子。1个培养皿为1次重复,3次重复为一个处理。分别向每一培养皿中加入预先配制好的10 mL等体积的不同浓度胁迫溶液,随后放置于(25±2) ℃下恒温培养箱内培养发芽。
1.2.1萌发指标测定 本试验于2017年7月22日开始,每日早上8:30观察记录发芽数,以芽长不短于种子长的1/2,胚根至少与种子等长作为发芽标准,并用称重法每日补充蒸发的水分。培养至第14天时,从各处理的3个重复中各随机挑选10株健壮且生长一致的幼苗,用滤纸吸干幼苗表面附着水分后使用0.1 mg 电子天平称量其苗重,然后剪下胚根、胚芽,分别测定胚根长(RL,length of root)和胚芽长(SL,length of shoot),计算发芽势(GP,germination potentia)、发芽率(GR,germination rate)、发芽指数(GI,germination index)和种子活力指数(VI,vigour index)。计算公式如下:
GP=(前4 d种子发芽数/种子总数)×100%
GR=(14 d种子发芽数/种子总数)×100%
GI=∑(Gt/Dt)
式中:Gt为第t天的发芽数;Dt为萌发第t天。
VI=GI×苗重
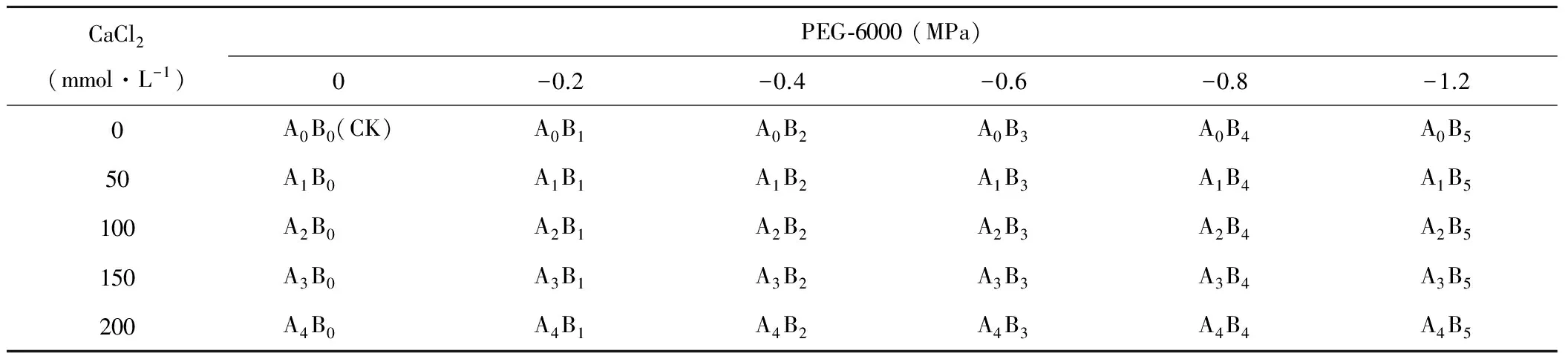
表1 试验处理方案Table 1 Treatments of the experiment
1.2.2粗酶液的制备 胁迫结束后,称取不同处理的燕麦幼苗叶片各0.1 g,用蒸馏水洗净,加入预冷的提取液缓冲液9 mL,冰浴充分研磨,以7500 r·min-1冷冻离心15 min,取上清液装入预先编号的收集管中,用于生理指标测定。
1.2.3生理指标的测定 可溶性蛋白含量的测定采用考马斯蓝亮G-250染色法[16];丙二醛(MDA)含量的测定参照硫代巴比妥酸(TBA)检测法[17];超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)以及谷胱甘肽(GSH)的测定使用试剂盒法,按照试剂盒说明书操作、比色,并运用试剂盒提供的公式计算出活力值,所有试剂盒均由南京建成生物有限公司提供。以上指标均使用美国BioTek 公司生产的多功能酶标仪(PMT49984)进行测定。
1.3 数据分析
使用Microsoft Excel 2007软件对试验数据进行整理和统计分析,SPSS 18.0软件进行单因素方差分析,采用Sigma Polt 10.0绘图。利用隶属函数法[18]对CaCl2、PEG单一胁迫以及交互胁迫下燕麦耐受性进行综合评价。每个样品各项指标的具体隶属函数值计算公式为:
Xu=(X-Xmin)/(Xmax-Xmin)
(1)
Xu=1-(X-Xmin)/(Xmax-Xmin)
(2)
式中:X为参试样品某一抗性指标的测定值,Xmax、Xmin分别为所有试样中该指标的最大值和最小值,若所测指标与燕麦的耐受性呈正相关,则使用(1)式计算隶属值,负相关则用(2)式。最后把每个试样的各项指标隶属函数值进行累加,取其平均值。平均值越大,抗性越强,反之越弱。
2 结果与分析
2.1 单一胁迫及交互胁迫处理下燕麦种子的萌发特征
由表2可见,CaCl2单一胁迫下燕麦的发芽率、发芽势、胚根长、发芽指数、活力指数均随着浓度增大呈先增后减趋势,在50、100 mmol·L-1CaCl2处理下的发芽率、发芽势、发芽指数、活力指数均显著高于对照CK(P<0.05);CaCl2浓度为150 mmol·L-1时,发芽率与对照差异不显著(P>0.05),发芽势显著高于对照;当浓度达到200 mmol·L-1时,发芽势和发芽率显著低于对照(P<0.05),这说明适宜中低浓度的CaCl2可促进种子萌发,而高浓度的CaCl2则会抑制种子萌发。胚芽长随着CaCl2浓度增大呈逐渐增加趋势,说明CaCl2能够促进燕麦胚芽的生长。
在PEG单一胁迫下(表2),燕麦的发芽率、发芽势、发芽指数、活力指数均随着胁迫浓度增大表现为先增后减趋势,发芽率、发芽势、发芽指数、活力指数在-0.4、-0.6 MPa PEG处理下均显著高于对照(P<0.05)且活力指数和发芽指数在-0.4 MPa时达到最大值,发芽率和发芽势在-0.6 MPa时达到最大值;当渗透势增大到-1.2 MPa时,发芽率、发芽势、发芽指数、活力指数均显著低于对照。表明适宜的PEG胁迫能促进燕麦种子的萌发。胚根长随胁迫浓度增大逐渐缩短,胚芽长随胁迫浓度增大逐渐伸长,这说明PEG处理会抑制胚根生长,但能促进胚芽生长。
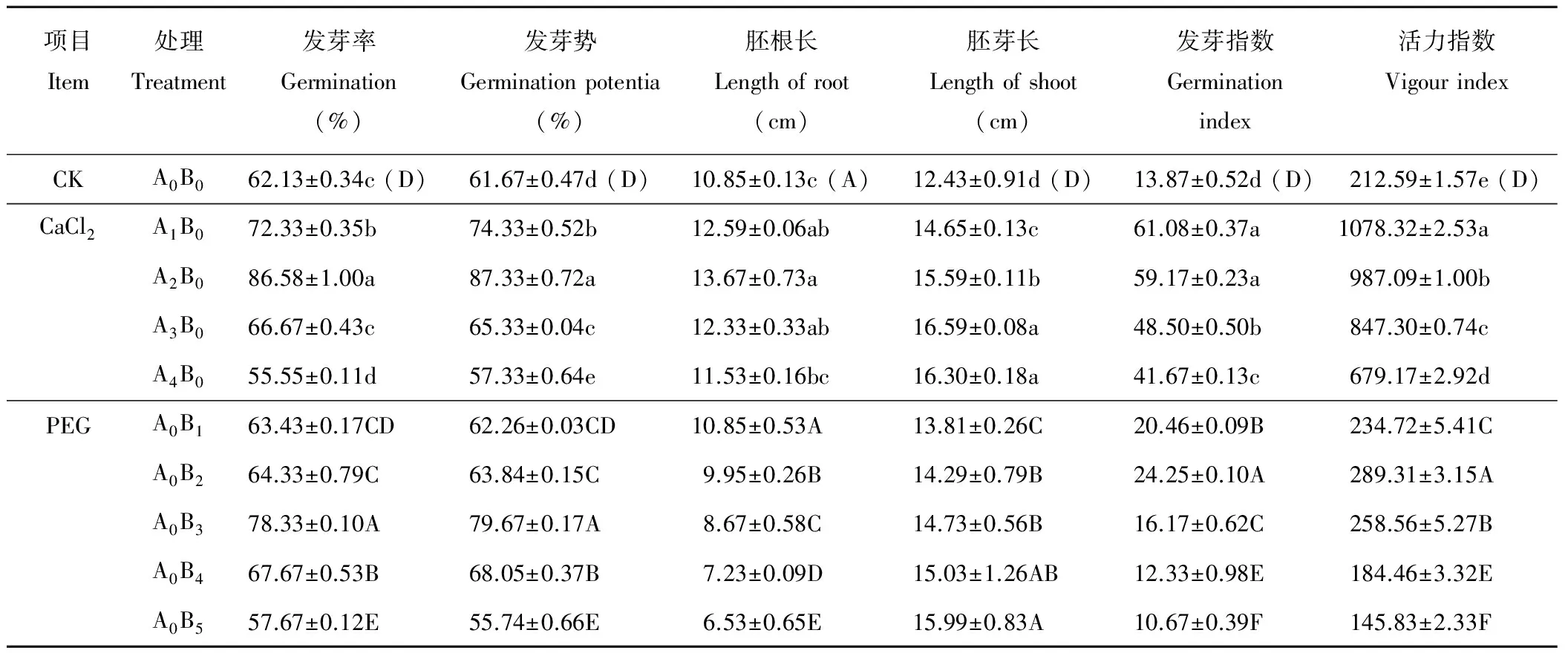
表2 单一胁迫对燕麦种子萌发参数的影响Table 2 Effects of single stress on oat seed germination
中低浓度交互作用对干旱胁迫下燕麦种子的发芽率和发芽势存在促进作用。由表3可见,当PEG渗透势为-0.2,-0.4 MPa时,A0B1、A0B2组的发芽率、发芽势与对照差异均不显著,经50、100 mmol·L-1CaCl2处理后,发芽率、发芽势显著提高;150 mmol·L-1时,发芽率、发芽势与对照差异不显著(P>0.05);当CaCl2浓度达到200 mmol·L-1时,发芽率和发芽势显著低于对照。PEG渗透势为-0.6 MPa时,A0B3组发芽率、发芽势显著高出对照26.07%、29.29%,经50、100 mmol·L-1CaCl2处理后,A1B3、A2B3组的发芽率、发芽势分别高出对照31.45%、30.81%,42.22%、41.61%。当PEG渗透势达到-0.8 MPa时,仅A0B4组高于对照,其余处理组均显著低于对照(P<0.05)。同一PEG处理下,CaCl2浓度的增大不再促进种子萌发。同一PEG渗透势处理下,适宜中低浓度的CaCl2可以明显增大种子的发芽指数和活力指数,但过高的CaCl2浓度会降低种子的发芽指数和活力指数。同一PEG渗透势处理下,胚根长随着CaCl2浓度的增大呈逐渐减小趋势;胚芽长在-0.2至-0.6 MPa PEG处理下随CaCl2浓度的增大呈逐渐伸长趋势,在-0.8至-1.2 MPa PEG处理下则随着CaCl2浓度的增大呈逐渐缩短趋势。以上表明在特定的干旱环境下,一定浓度的CaCl2能增强燕麦种子对干旱环境的适应并促进萌发;而CaCl2浓度过高则会产生相反的效应。
2.2 单一胁迫及交互胁迫处理下燕麦幼苗的生理特性
在CaCl2单一胁迫下,燕麦幼苗的可溶性蛋白含量、POD、CAT、SOD、GSH活性随胁迫浓度的增大呈先升后降趋势(图1),这5个指标均在50 mmol·L-1CaCl2时开始显著高于对照(P<0.05),并且在100 mmol·L-1CaCl2时达到最大值,说明适宜浓度的CaCl2可以增加燕麦可溶性蛋白含量并提高抗氧化酶活性;150 mmol·L-1CaCl2时SOD活性与对照差异不显著(P>0.05);当继续增大胁迫浓度到200 mmol·L-1时,4个抗氧化酶活性均显著低于对照(P<0.05),这可能是该胁迫浓度已超过了燕麦过氧化防御系统的阈值导致的。MDA含量随CaCl2胁迫浓度的增大呈逐渐上升趋势,在50 mmol·L-1时与CK差异不显著(P>0.05),这可能是因为该胁迫浓度还未对燕麦造成伤害,没有发生膜脂过氧化,而随着胁迫浓度增大对燕麦幼苗的伤害加重则产生了更多的MDA,在200 mmol·L-1时达到最大值6.63 μmol·g-1。
在PEG单一胁迫下,可溶性蛋白含量、POD、CAT、SOD、GSH活性也随胁迫浓度增大表现为先升后降趋势(图2),且均在-0.6 MPa 时显著高于其余处理组(P<0.05);抗氧化酶活性在-0.8 MPa PEG胁迫时骤降且显著低于-0.4和-0.6 MPa 处理组。MDA含量随胁迫浓度增大呈逐渐上升趋势,在-1.2 MPa PEG胁迫时达到最大值12.05 μmol·g-1且显著高于其余处理组(P<0.05)。
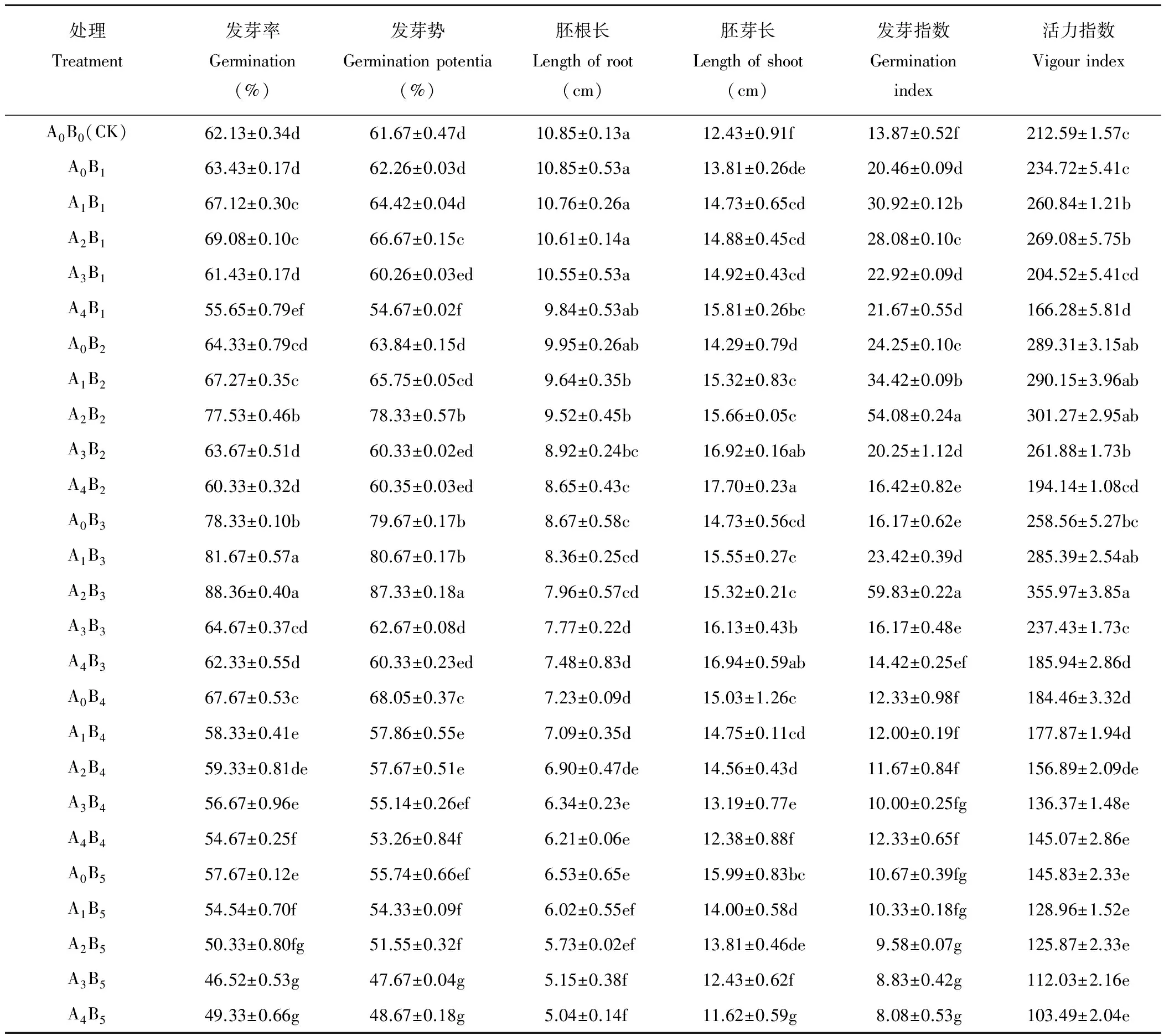
表3 交互胁迫对燕麦种子萌发参数的影响Table 3 Effects of multiple stresses on oat seed germination
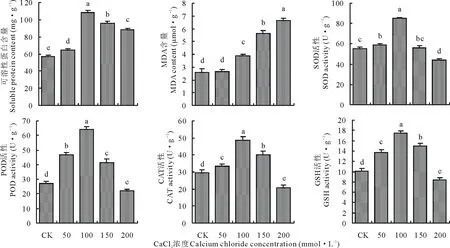
图1 CaCl2单一胁迫对燕麦幼苗生理指标的影响Fig.1 Effects of a single stress (CaCl2) on physiological indices of oat seedlings
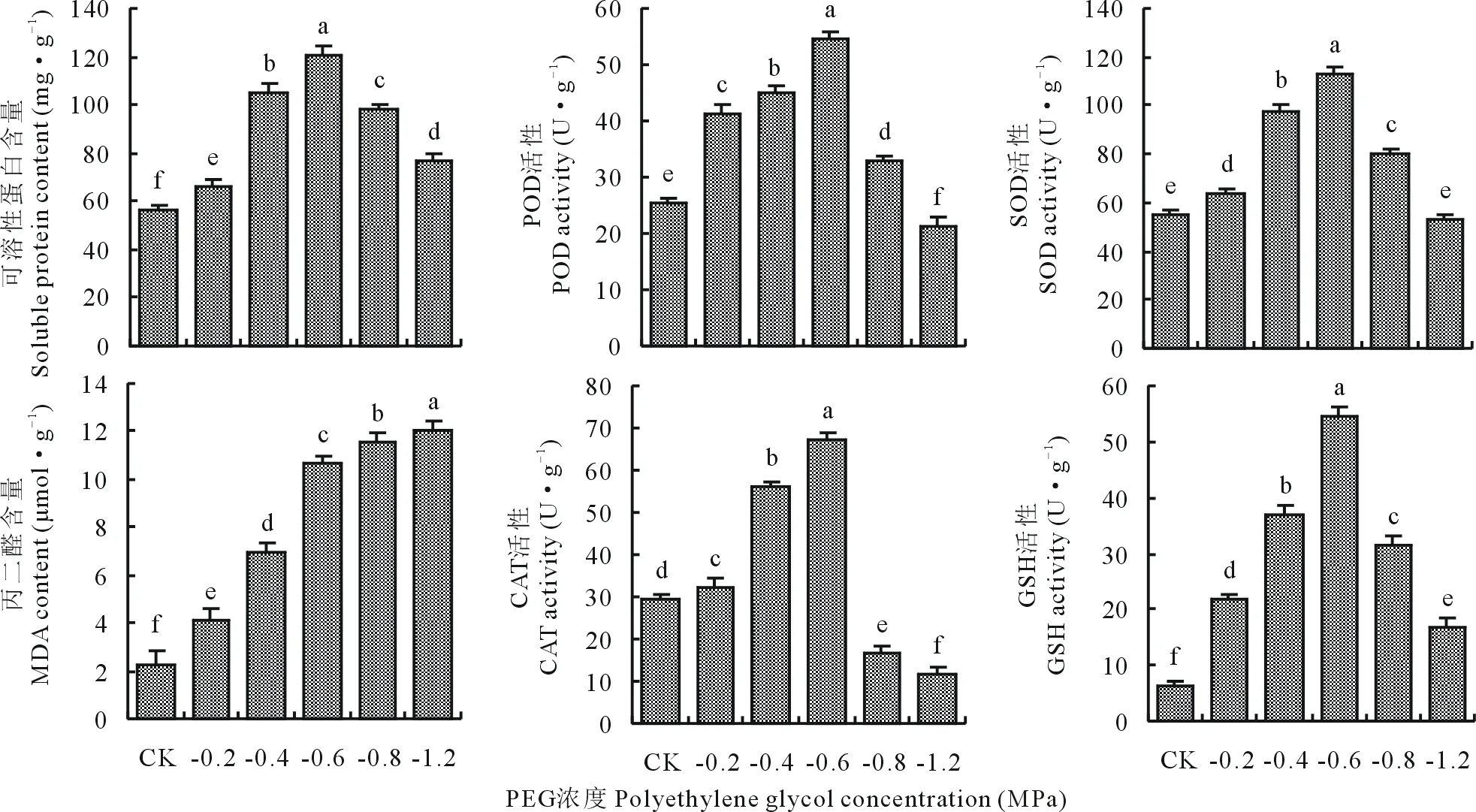
图2 PEG单一胁迫对燕麦幼苗生理指标的影响Fig.2 Effects of a single stress (PEG) on physiological indices of oat seedlings
由图3可知,可溶性蛋白含量与加入CaCl2处理和PEG单一处理时的变化趋势相似,但变化幅度存在差异。-0.6 MPa PEG+50mmol·L-1CaCl2和-0.6 MPa PEG+100 mmol·L-1CaCl2处理下的可溶性蛋白含量为125.18和136.47 mg·g-1,显著高出对照120.12%、139.97%(P<0.05);而-1.2 MPa PEG+200 mmol·L-1CaCl2处理下的可溶性蛋白含量仅为46.12 mg·g-1,显著低于对照。
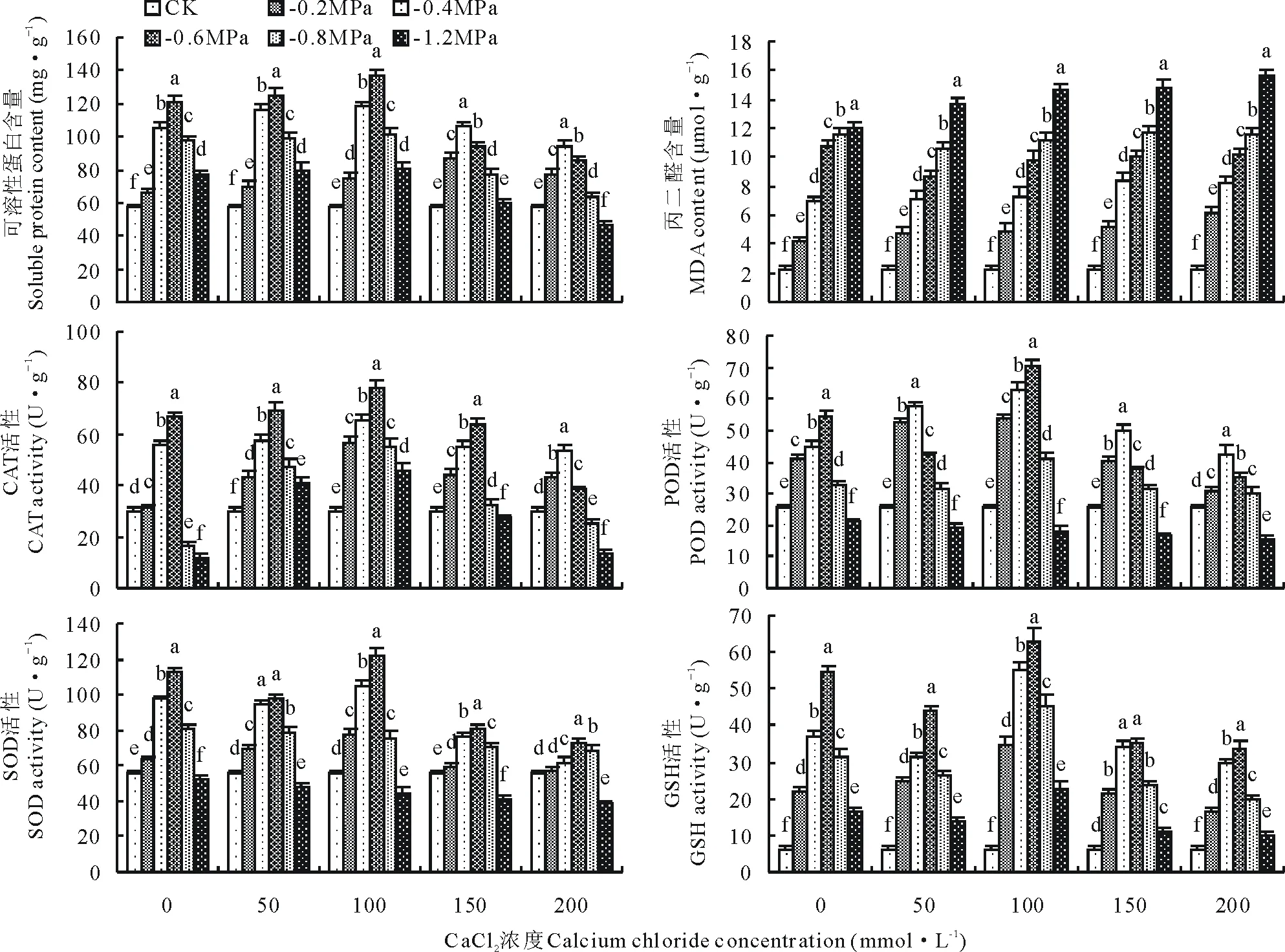
图3 交互胁迫对燕麦幼苗生理指标的影响Fig.3 Effects of multiple stresses on physiological indices of oat seedlings
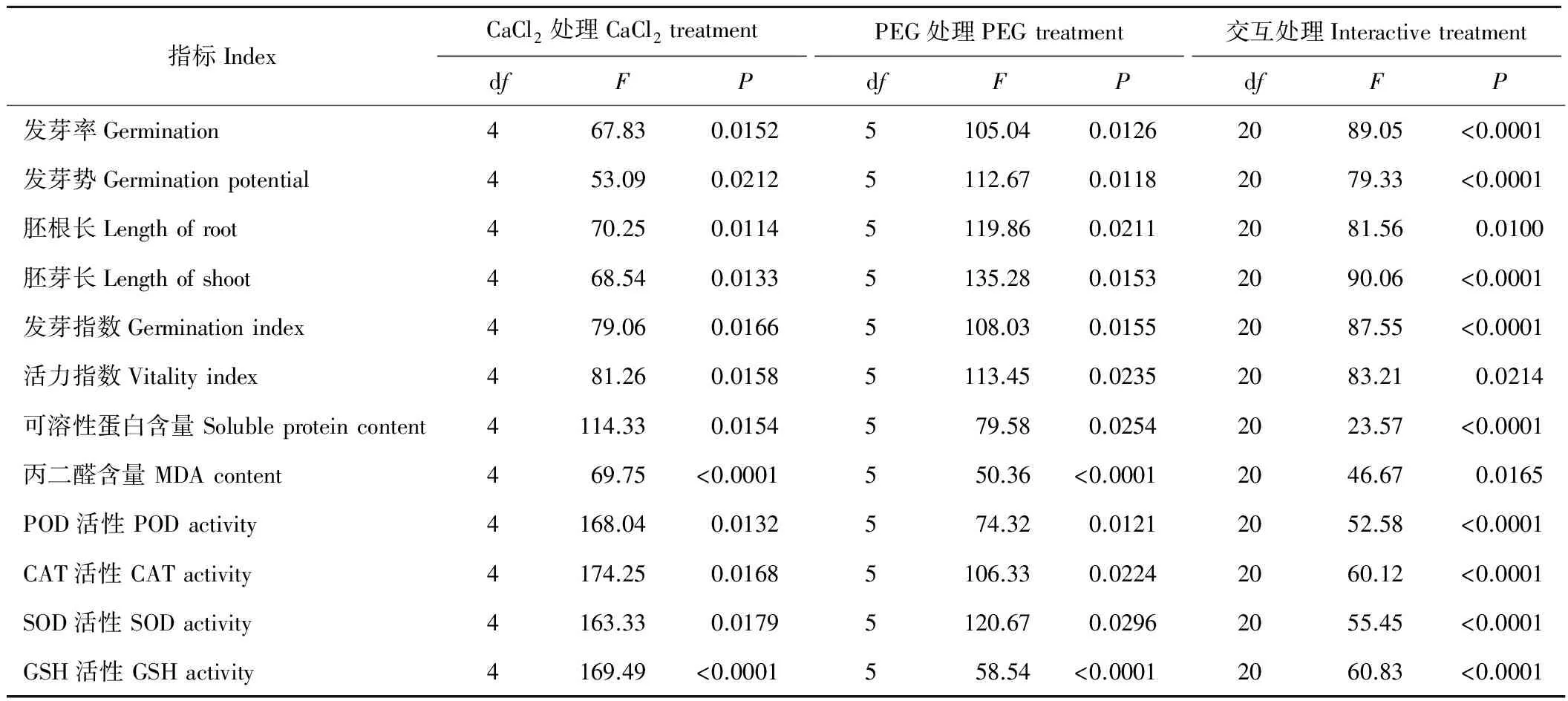
表4 CaCl2和PEG交互处理对燕麦种子萌发和生理指标影响的双因素方差分析Table 4 The two-way analysis of variance evaluating the effects of CaCl2 and PEG interactions on oat seed germination and physiological indices
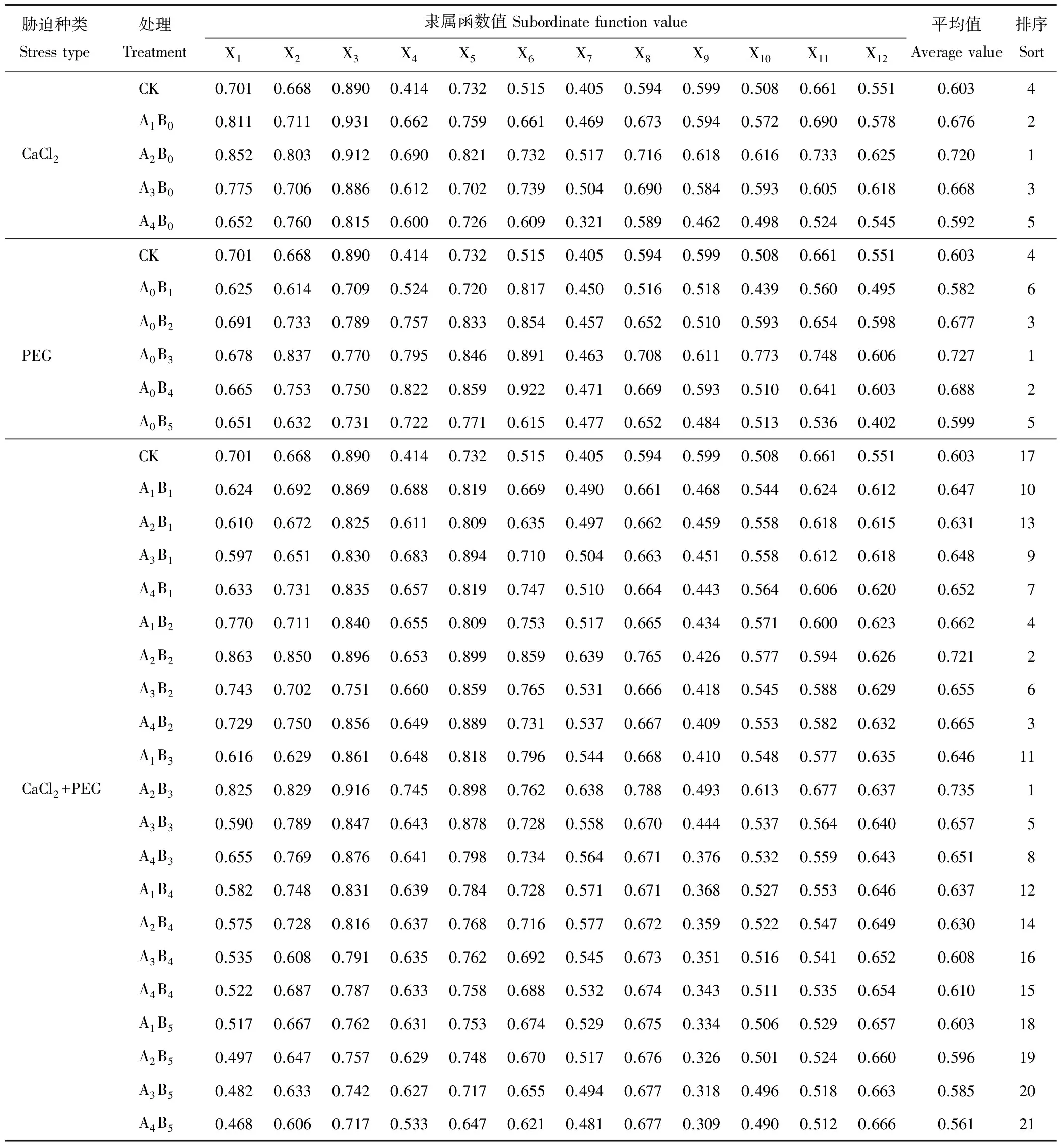
表5 CaCl2、PEG单一及交互处理下燕麦耐受性综合评价Table 5 The tolerance evaluation of single CaCl2, PEG and interactive treatment in oat germination
在不同CaCl2浓度处理下,燕麦幼苗的MDA含量均随PEG渗透势增大呈逐渐上升趋势,并且在同一CaCl2浓度下不同PEG处理间差异显著(P<0.05),-1.2 MPa PEG+200 mmol·L-1CaCl2时达到最大值15.57 μmol·g-1。
POD、SOD、CAT、GSH是植物在逆境胁迫下为抵御外界伤害产生的抗氧化酶类。在不同CaCl2浓度下(-0.4 MPa PEG+50 mmol·L-1CaCl2处理除外),抗氧化酶活性随PEG渗透势增大到-0.6 MPa时均表现为随胁迫加重活性增强,并且均在-0.6 MPa PEG+100 mmol·L-1CaCl2处理时达到最大值;当PEG渗透势超过-0.6 MPa 之后则表现为随着胁迫增大活性降低。其中,POD、SOD、CAT活性在-1.2 MPa PEG+150 mmol·L-1CaCl2、-1.2 MPa PEG+200 mmol·L-1CaCl2处理下均显著低于对照(P<0.05)。以上说明适当中低浓度的交互胁迫可以增加幼苗的可溶性蛋白含量和提高抗氧化酶活性,但胁迫过重则会起反作用。
2.3 单一胁迫与交互胁迫差异比较及耐受性综合评价
交互胁迫对燕麦种子萌发参数和生理指标影响的双因素方差分析结果见表4。由表4可知,燕麦的发芽率、发芽势、胚根长、胚芽长、发芽指数、活力指数在CaCl2、PEG单因素处理和两者交互处理下的表现存在差异,其中单因素处理下各个指标间差异显著(P<0.05);交互处理下,仅胚根长、活力指数两者差异显著(P<0.05),发芽率、发芽势、胚芽长、发芽指数均差异极显著(P<0.01)。燕麦幼苗的可溶性蛋白含量、POD、CAT、SOD活性在单因素处理下差异显著(P<0.05),而在交互处理下差异极显著(P<0.01);丙二醛含量恰好相反:在单因素处理下差异极显著(P<0.01),交互处理下差异显著(P<0.05);GSH活性在不同处理下均表现为差异极显著(P<0.01)。综上可知,交互胁迫对燕麦种子萌发和幼苗生理的影响比单一胁迫更具优势。
通过以上种子萌发情况及幼苗生理指标的变化分析发现,燕麦在单一及交互胁迫下存在一定的适应机制,在不同胁迫下均存在着胁迫临界浓度,当超过该胁迫浓度后燕麦对此逆境的适应能力降低。本研究为探究燕麦对不同胁迫条件的耐受临界点,采用隶属函数法分别对CaCl2、PEG单一胁迫及交互胁迫处理下的耐受性进行全面、综合的评价。隶属函数结果分析表明(表5),在CaCl2单一处理下,A2B0处理的隶属函数均值最大,燕麦反应最为敏感,即100 mmol·L-1CaCl2为燕麦抵御钙盐胁迫的临界浓度;在PEG单一处理下,A0B3处理时隶属函数值最大,即-0.6 MPa PEG为燕麦耐旱性的临界浓度;在CaCl2+PEG交互胁迫处理下,根据交互胁迫处理下各指标的隶属函数值,评价出燕麦对各个处理的耐受性强弱依次为:A2B3>A2B2>A4B2>A1B2>A3B3>A3B2>A4B1>A4B3>A3B1>A1B1>A1B3>A1B4>A2B1>A2B4>A4B4>A3B4>CK>A1B5>A2B5>A3B5>A4B5。其中A2B3(100 mmol·L-1CaCl2+-0.6 MPa PEG)处理效果较突出,为交互胁迫的临界浓度,且其隶属函数值高于单一处理下表现最佳的A2B0和A0B3。
3 讨论
种子萌发是高等植物生长发育的起点,是植物生命史的关键阶段[19]。种子萌发阶段的耐盐、耐旱情况可在一定程度上反映该物种的耐盐、旱程度[20],而发芽率、发芽势、发芽指数和活力指数是检测种子质量的重要指标。赵丽丽等[21]对二色胡枝子(Lespedezabicolor)的研究指出,在低盐、低旱环境下,蛋白含量高、吸水能力较强的种子在萌发过程中吸胀损伤得以缓解,从而促进种子萌发,起到提高种子萌发耐盐、旱的作用。本研究中,单一及交互胁迫下燕麦种子的发芽率、发芽势、发芽指数以及活力指数在中低浓度胁迫时均呈现出随胁迫加重促进萌发的趋势,与此研究结论相同。然而,随着单一和交互胁迫浓度的持续增大,种子内部膜系统遭到破坏,燕麦种子萌发受到抑制,说明当胁迫浓度增加到一定程度时会对种子造成损伤并导致原本的促进作用转变为抑制作用,这与蔡喜悦等[22]对复羽叶栾树(Koelreuteriabipinnata)的研究结果一致。本研究中,种子胚根长在PEG单一处理及交互处理下均随干旱胁迫程度的加重呈逐渐缩短趋势,这与周敏等[23]对伞花木(Eurycorymbuscavaleriei)和马缨杜鹃(Rhododendrondelavayi)的研究结果类似,表明种子胚根长主要受干旱胁迫的影响。
植物体内的可溶性蛋白大多是参与各种代谢的酶类,其含量是了解植物总代谢的一个重要指标[24]。相关研究[25-26]表明,低浓度干旱或钙盐胁迫可提高蛋白酶活性,促进贮藏蛋白水解为可溶性蛋白,增加叶片中可溶性蛋白含量;而当胁迫浓度过高时则会破坏质膜内外的离子平衡,同时扰乱细胞质中的Ca2+状态,破坏以磷酸为基础的能量代谢和质膜离子平衡[27],导致可溶性蛋白含量下降。本研究中,可溶性蛋白含量在单一胁迫及交互胁迫时均表现为随胁迫浓度增大呈先增后减趋势,与前人研究结论相似。
植物在生长代谢过程中会产生超氧化物自由基等活性氧[28],钙盐、干旱胁迫不仅能提高细胞的活性氧水平,还能诱导植物防御体系的建立,从而避免或缓解活性氧对植物的伤害[29]。在正常生理条件下,这些活性氧自由基的产生和消除存在一个动态平衡,对植物本身没有任何伤害;而在钙盐、干旱胁迫或其他逆境胁迫条件下,若大量产生的活性氧自由基不能被及时清除时这种平衡将会被破坏[30]。本研究中,在单一及交互胁迫下,燕麦幼苗的超氧化物歧化酶、过氧化氢酶、过氧化物酶以及谷胱甘肽活性均在中低浓度胁迫时随浓度增大而逐渐增加,这些酶活性的增强有利于及时清除活性氧自由基,保持平衡;而随着胁迫浓度的进一步增大,这几种抗氧化酶已不能完全清除存在的活性氧自由基,平衡被打破,因此活性降低。
MDA含量是植物受逆境胁迫时发生膜脂过氧化的最终产物,其含量的多少直接反映植物细胞发生膜脂过氧化强弱的程度[31]。在不同胁迫处理条件下MDA含量均随胁迫浓度的增大逐渐增加,这是细胞受到严重伤害的一种表现,在交互胁迫时MDA含量较单一胁迫产生更多,究其原因可能是CaCl2对植物的胁迫效应主要为渗透胁迫和离子胁迫,PEG仅为渗透胁迫,两者交互处理时,渗透胁迫伤害效应叠加[32],加重燕麦幼苗受损程度,表现为更多的MDA累积,这与蒋明敏等[33]对石蒜(Lycorisradiata)的研究结论相似。
本研究从植物对钙盐、干旱互作机制的角度分析了燕麦对钙盐、干旱岩溶生境下种子萌发及幼苗的生理生化适应机制,发现中低浓度的单一胁迫及交互胁迫均有利于促进种子萌发和增强幼苗的抗氧化酶活性,根据隶属函数法综合评价得知,CaCl2单一胁迫的临界浓度为100 mmol·L-1,PEG单一胁迫的临界浓度为-0.6 MPa,交互胁迫在100 mmol·L-1CaCl2+-0.6 MPa PEG浓度时的处理效果明显优于单一胁迫处理下的最佳效果;相比于单一胁迫,交互胁迫下的隶属函数最大值更大,并且在交互胁迫下植物能通过钙盐、干旱交互增效作用来提高自身对环境的适应性,表现出更强的耐受性,从而具有相对较高的萌发率和较好的生理适应性。萌发指标和生理指标的双因素方差分析结果中:种子的发芽率、发芽势、胚根长、胚芽长、发芽指数、活力指数在单一胁迫处理时差异显著,而盐旱交互处理时,发芽率、发芽势、胚芽长、发芽指数差异极显著;幼苗的可溶性蛋白、POD、CAT、SOD活性在单一胁迫处理时差异显著,而在交互处理下差异极显著的表现证实了这一结论的准确性。
4 结论
在岩溶地区燕麦种植生产应用中选择中低浓度盐旱交互胁迫区域进行种植可以促进种子的萌发和幼苗生长。但在自然环境条件下,植物生长过程中受到的外界胁迫并不单一,胁迫程度也不尽相同,尤其在以钙盐、干旱为生境特征的岩溶地区,富钙干旱环境对植物的生长发育影响较大,因而,岩溶生境下植物的逆境响应机制还有待于后期进行更深入细致的研究。
参考文献References:
[1] Liu J X, Wang J C, Wang R J,etal. Effects of drought stress on physiological characteristics of oat seedlings. Agricultural Research in Arid Areas, 2014, 32(3): 24-28.
刘建新, 王金成, 王瑞娟, 等. 旱盐交叉胁迫对燕麦幼苗叶片生理特性的影响. 干旱地区农业研究, 2014, 32(3): 24-28.
[2] Lin W J, Wu G F, Li C H,etal. Effects of cultivar and environment on nutritional quality of chinese naked oats. Acta Agronomica Sinica, 2011, 37(6): 1087-1092.
林伟静, 吴广枫, 李春红, 等. 品种与环境对我国裸燕麦营养品质的影响. 作物学报, 2011, 37(6): 1087-1092.
[3] Chen Y, Wang P C, Zhao L L,etal. Effects of exogenous calcium on the physiological characters ofCajanuscajanunder drought stress. Acta Agrestia Sinica, 2014, 22(5): 1051-1055.
陈莹, 王普昶, 赵丽丽, 等. 外源钙对干旱胁迫下木豆种苗生理特性的影响. 草地学报, 2014, 22(5): 1051-1055.
[4] Zhao L L, Wang P C, Chen C,etal. Effects of exogenous Ca2+on the growth, physiological characters ofSophoraviciifoliaseedlings in karst mountain area under the drought stress. Journal of Nuclear Agricultural Sciences, 2017, 31(10): 2036-2046.
赵丽丽, 王普昶, 陈超, 等. 外源钙对干旱胁迫下岩溶山区白刺花幼苗生长、生理特性的影响. 核农学报, 2017, 31(10): 2036-2046.
[5] Xiong L M, Schumaker K S, Zhu J K. Cell signaling during cold drought and salt stress. Plant Cell, 2002, 14(8): 165-183.
[6] Zhou Y C, Wang S J, Lu H M. Spatial distribution of soil in the process of rocky desertification in karst. Earth and Environment, 2010, 38(1): 1-7.
周运超, 王世杰, 卢红梅. 喀斯特石漠化过程中土壤的空间分布. 地球与环境, 2010, 38(1): 1-7.
[7] Huang F, Yang C, Cao J. Response of germination physiology ofCajanuscajanseeds to drought stress: comparison between karst water and allogenic water treatments. Journal of Resources & Ecology, 2015, 6(4): 263-268.
[8] Hao F, Liu X, Zhang X,etal. Effect of complex saline alkali stress on nitrogen and phosphorus absorption and physiological traits of alfalfa seedlings. Journal of the American College of Cardiology, 2015, 64(16): C237.
[9] Bai X L, Zhou Q P, Chen S Y,etal. Effects of PEG simulated drought stress on seed germination of six oat cultivars. Journal of South-west Agricultural University (Natural Science Edition), 2015, 41(2): 133-137.
柏晓玲, 周青平, 陈仕勇, 等. PEG模拟干旱胁迫对6种燕麦品种种子萌发的影响. 西南民族大学学报(自然科学版), 2015, 41(2): 133-137.
[10] Liu F Q, Liu J L, Zhu R F,etal. Physiological responses and tolerance of four oat varieties to salt stress. Acta Prataculturae Sinica, 2015, 24(1): 183-189.
刘凤歧, 刘杰淋, 朱瑞芬, 等. 4种燕麦对NaCl胁迫的生理响应及耐盐性评价. 草业学报, 2015, 24(1): 183-189.
[11] Zhang N, Zhao B P, Guo R L,etal. Effect of water stress on physiological characteristics of different oat cultivars. Journal of Triticeae Crops, 2012, 32(1): 150-156.
张娜, 赵宝平, 郭若龙, 等. 水分胁迫对不同抗旱性燕麦品种生理特性的影响. 麦类作物学报, 2012, 32(1): 150-156.
[12] Gao Z W, Nin J X, Shao S,etal. Effects of complex salt-alkali stresses on seed germination of oat. Pratacultural Science, 2014, 31(3): 451-456.
高战武, 蔺吉祥, 邵帅, 等. 复合盐碱胁迫对燕麦种子发芽的影响. 草业科学, 2014, 31(3): 451-456.
[13] Liu J X, Wang J C, Liu X L. Effects of salt-alkaline mixed stress on proline and polyamine metabolism in leaves of naked oat seedlings. Chinese Journal of Ecology, 2016, 35(11): 2974-2982.
刘建新, 王金成, 刘秀丽. 盐碱混合胁迫对裸燕麦幼苗叶片脯氨酸和多胺代谢的影响. 生态学杂志, 2016, 35(11): 2974-2982.
[14] Liu J X, Wang J C, Wang R J,etal. Effects of drought stress on the growth and osmotic adjustment of oat seedlings. Journal of Soil and Water Conservation, 2012, 26(3): 244-248.
刘建新, 王金成, 王瑞娟, 等. 旱盐交叉胁迫对燕麦幼苗生长和渗透调节物质的影响. 水土保持学报, 2012, 26(3): 244-248.
[15] Yan Q Z, Bi X H. International rules for seed testing. Beijing: China Agriculture Press, 1999.
颜启传,毕新华. 国际种子检验规程. 北京: 中国农业出版社, 1999.
[16] Gao J F. Plant physiology experiment guidance. Beijing: China Higher Education Press, 2006.
高俊凤. 植物生理学实验指导. 北京: 高等教育出版社, 2006.
[17] Li H S. Principle and technology of plant physiology and biochemistry experiment. Beijing: Higher Education Press, 2000.
李合生. 植物生理生化试验原理和技术. 北京: 高等教育出版社, 2000.
[18] Guo Y D, Ren Y X, Liu G H,etal. Effects of exogenous calcium and gibberellin on physiological characteristics of alfalfa seedlings under drought stress. Journal of Grassland Science, 2015, 24(7): 89-96.
郭郁频, 任永霞, 刘贵河, 等. 外源钙和赤霉素对干旱胁迫下苜蓿幼苗生理特性的影响. 草业学报, 2015, 24(7): 89-96.
[19] Yan Q Z. International rules for seed testing. Beijing: China Agriculture Press, 2001.
颜启传. 种子学. 北京: 中国农业出版社, 2001.
[20] Chen Y, Zhong L, Zhao L L,etal. Physiological and biochemical responses of the seed germination period to the high calcium drought in the karst habitat. Jiangsu Agricultural Sciences, 2014, 42(9): 335-339.
陈莹, 钟理, 赵丽丽, 等. 截叶铁扫帚种子萌发期对岩溶生境高钙干旱的生理生化反应. 江苏农业科学, 2014, 42(9): 335-339.
[21] Zhao L L, Wang P C, Chen C,etal. Effects of exogenous calcium on the seed germination ofLespedezabicolorunder drought stress. Acta Agrestia Sinica, 2015, 23(1): 120-124.
赵丽丽, 王普昶, 陈超, 等. 干旱胁迫下外源钙对二色胡枝子种子萌发的影响. 草地学报, 2015, 23(1): 120-124.
[22] Cai X Y, Chen X D, Li C Z,etal. Effects of exogenous Ca2+on the seed germination ofKoelreuteriapaniculatain limestone area of Southwest China under drought stress. Chinese Journal of Applied Ecology, 2013, 24(5): 1341-1346.
蔡喜悦, 陈晓德, 李朝政, 等. 干旱胁迫下外源钙对石灰岩地区复羽叶栾树种子萌发的影响. 应用生态学报, 2013, 24(5): 1341-1346.
[23] Zhou M. Effects of exogenous calcium on seed germination ofEurycorymbuscavalerieiandRhododendrondelavayi. Chinese Agricultural Information, 2014, 17: 27-28.
周敏. 干旱胁迫下外源钙对伞花木和马缨杜鹃种子萌发的影响. 中国农业信息, 2014, 17: 27-28.
[24] Chen C S, Xie Z X, Liu X J. Interactive effects of drought and salt stresses on winter wheat seedlings growth and physiological characteristics of stress-resistance. Chinese Journal of Applied Ecology, 2009, 20(4): 811-816.
陈成升, 谢志霞, 刘小京. 旱盐互作对冬小麦幼苗生长及其抗逆生理特性的影响. 应用生态学报, 2009, 20(4): 811-816.
[25] Fu S L, Zhou Y B, He X Y,etal. Effects of drought stress on photosynthetic physiological indexes of poplar. Chinese Journal of Applied Ecology, 2006, 17(11): 2016-2019.
付士磊, 周永斌, 何兴元, 等. 干旱胁迫对杨树光合生理指标的影响. 应用生态学报, 2006, 17(11): 2016-2019.
[26] Zhu L J, Yan Q J. Effect of exogenous calcium chloride on salt stress injury inGarlicseedlings. Jiangsu Agricultural Sciences, 2016, 44(8): 242-244.
朱利君, 闫秋洁. 外源氯化钙对大蒜幼苗盐胁迫伤害的缓解作用. 江苏农业科学, 2016, 44(8): 242-244.
[27] Sun D Y, Guo Y L, Ma L G. Cell signal transduction. Beijing: Science Press, 1998.
孙大业, 郭艳林, 马力耕. 细胞信号转导. 北京: 科学出版社, 1998.
[28] Gu X H, Sun L Q, Gao B,etal. Effects of calcium on physiological characteristics, yield and quality ofPeanutunder drought stress. Chinese Journal of Applied Ecology, 2015, 26(5): 1433-1439.
顾学花, 孙莲强, 高波, 等. 施钙对干旱胁迫下花生生理特性、产量和品质的影响. 应用生态学报, 2015, 26(5): 1433-1439.
[29] Jiang Y B, Cui G W, Li H. Effect of coatingMedicagosativaseeds in calcium on drought resistance and interrelated physiological indexes under drought conditions. Acta Prataculturae Sinica, 2005, 14(5): 32-36.
姜义宝, 崔国文, 李红. 干旱胁迫下外源钙对苜蓿抗旱相关生理指标的影响. 草业学报, 2005, 14(5): 32-36.
[30] Li Q, Cao J H, Yu L J,etal. Effects of exogenous calcium on the photosynthetic characteristics ofLonicerajaponicaThunb under drought stress. Ecology and Environmental Sciences, 2010, 19(10): 2291-2296.
李强, 曹建华, 余龙江, 等. 干旱胁迫过程中外源钙对忍冬光合生理的影响. 生态环境学报, 2010, 19(10): 2291-2296.
[31] Fan S L, Yuan Z H, Feng L J,etal. Effects of drought stress on physiological and biochemical parameters ofDahliapinnata. Chinese Journal of Applied Ecology, 2011, 22(3): 651-657.
范苏鲁, 苑兆和, 冯立娟, 等. 干旱胁迫对大丽花生理生化指标的影响. 应用生态学报, 2011, 22(3): 651-657.
[32] He M, Yang X R, Liu X D. Effect of Ca2+on the physiological response and mesophyll ultrastructure in leaves ofHemerocallismiddendorfiiunder osmotic stress. Chinese Journal of Grassland, 2012, 34(1): 84-88.
何淼, 杨絮茹, 刘晓东. 钙对渗透胁迫下大花萱草幼苗生理响应及叶肉组织超微结构的影响. 中国草地学报, 2012, 34(1): 84-88.
[33] Jiang M M, Xu S, Xia B,etal. Effects of exogenous calcium chloride, salicylic acid and nitric oxide on drought resistance ofLycorisradiataunder drought stress. Journal of Plant Physiology, 2012, 48(9): 909-916.
蒋明敏, 徐晟, 夏冰, 等. 干旱胁迫下外源氯化钙、水杨酸和一氧化氮对石蒜抗旱性的影响. 植物生理学报, 2012, 48(9): 909-916.
尹国丽, 吴芳, 陶茸, 等. 苜蓿轮作玉米小麦土壤浸提液对苜蓿种子萌发和幼苗生理及生长的影响. 草业学报, 2018, 27(5): 153-161.
Yin G L, Wu F, Tao R,etal. Effects of rhizosphere soil extraction from alfalfa-corn and alfalfa-wheat fields on alfalfa seed germination and seedling physiology and growth. Acta Prataculturae Sinica, 2018, 27(5): 153-161.
猜你喜欢
中老年保健(2022年5期)2022-11-25
现代畜牧科技(2021年9期)2021-10-13
今日农业(2021年7期)2021-07-28
现代园艺(2020年19期)2020-10-02
种子(2019年6期)2019-07-13
种子(2019年4期)2019-05-28
植物研究(2019年2期)2019-03-19
现代农村科技(2019年4期)2019-01-06
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
