
多元判别法鉴别中国魔芋种
2018-06-29 04:58吴学尉刘丹丹杨雨嫣王丽花吴治成
江苏农业科学 2018年11期
吴学尉, 叶 辉, 乔 琴, 李 琳, 刘丹丹, 杨雨嫣,王丽花, 吴治成
(1.云南大学农学院,云南昆明 650504; 2.云南省农业科学院花卉研究所,云南昆明 650205;3.云南临沧滇晟农林科技有限公司,云南临沧 677500)
魔芋是天南星科(Araceae)魔芋属(AmorphophallusBlume)多年生草本植物[1-5]。目前全球已经记载的魔芋种已经达到200个[6]。Hetterscheid等指出魔芋主要分布在亚洲,即中南半岛和云南为主的我国南部和东南亚[7]。我国是魔芋的起源中心,目前分布和引种栽培的有17个种,其中10个种是我国分布的特有种[5]。云南是魔芋野生种质资源的主要分布中心,有近2 000年的魔芋栽培历史[8],经过长期的自然变异和栽培驯化,种类繁多,栽培品种混乱,出现很多同名异物和异名同种的现象[9-10]。魔芋喜好温暖湿润、适度遮阴的栽培环境,被引种到我国很多地区进行栽培,栽培面积较大的为云南、湖南、四川、广西、贵州、山西和江西等省(区)的阴凉山地[11-12],主要栽培花魔芋(Amorphophalluskonjac)、白魔芋(Amorphophallusalbus)、珠芽魔芋类型和一些地方品种[13-15]。在不同地区,很多栽培品种被给予了地方名,如西双版纳红魔芋、德宏花魔芋、富源花魔芋、井冈山花魔芋、金佛山魔芋、西盟种和红河种等[16],造成地方品种混杂。不同地方魔芋栽培品种间的特异性差异较大。植株大小、叶柄颜色、花器官形态以及球茎葡甘聚糖含量、魔芋精粉黏度等差异显著。魔芋球茎的葡甘聚糖含量和精粉黏度是区别魔芋品质的重要指标[17],不同魔芋种类的葡甘聚糖含量和精粉黏度差异巨大,如花魔芋葡甘聚糖含量为55%左右,白魔芋葡甘聚糖含量为60%左右,田阳魔芋葡甘聚糖含量为40%左右,而一些珠芽类魔芋种的葡甘聚糖含量高达76.6%[18],但有的魔芋种类葡甘聚糖含量很低,如滇魔芋的葡甘聚糖含量不到5%[19]。近年来,魔芋产品需求量持续增长,云南的很多地区掀起了魔芋种植热潮,各种不同类型的魔芋种源被用于生产栽培,但种苗生产者、种植者缺乏对魔芋种的识别能力,导致魔芋栽培品质和魔芋精粉质量极不稳定。因此,魔芋产业的发展亟需准确而简便的品种鉴别技术。
魔芋种类繁多且地方品种极其多样,为魔芋种的亲缘关系研究带来了挑战。张玉进等利用RAPD分子标记将22份魔芋资源分为5类[20]。腾彩珠利用AFLP技术结合ISSR多态性标记分析,发现白魔芋、珠芽魔芋分别与花魔芋、西盟魔芋亲缘关系较近[21]。蒋学宽利用ISSR分子标记技术将96份魔芋种在相似系数D=0.753时分为9大类,验证了数量分类学方法和性状选取的合理性。但这些分子标记技术还很难用于田间品种鉴别。蒋学宽研制的DUS(新颖性、特异性和稳定性)测试的55个性状主要用于新品种的申报[22],在品种鉴别时必须要有多年多点数据,鉴定周期长,对于田间栽培期间品种鉴别实用性不强。不同类型的魔芋种地下茎特征、叶柄、叶形、花器官和珠芽类型等具有丰富的多样性,且在无性繁殖后代可以稳定保持,而这些形态学特征仅见于中国植物志和Flora of China等分类研究,各种的特异性区别方法缺乏图文并茂的鉴别资料。
本研究以李恒等确认的在中国分布和栽培的17个魔芋种[25]为基础,利用模式标本,结合不同生长时期栽培照片及品种特性记录,利用魔芋地下茎、叶柄、叶形、花柄、花器官和珠芽类型等结构参数对其进行多元判别分析,用图文资料展示其区别关键点,为魔芋品种田间栽培鉴定提供借鉴。
1 材料与方法
1.1 材料
供试材料为白魔芋(A.albusLiu & Wei)、珠芽魔芋[A.bulbifer(Roxb.) Bl.]、桂平魔芋(A.coaetaneusLiu & Wei)、田阳魔芋(A.corrugatusN.E. Br.)、南蛇棒(A.dunniiTutch.)、红河魔芋(A.hayiHett.)、台湾魔芋(A.henryiN.E. Br.)、密毛魔芋(A.hirtusN.E. Br.)、勐海魔芋(A.kachinensisEngl. & Gehrm.)、东亚魔芋(A.kiusianusMakino)、花魔芋(A.konjacK. Koch)、西盟魔芋(A.krauseiEngl.)、米勒魔芋(A.muelleriBl. )、疣柄魔芋[A.paeoniifolius(Dennst.) Nicols.]、东京魔芋(A.tonkinensisEngl. & Gehrm.)、攸乐魔芋(A.yuloensisH. Li)和滇魔芋(A.yunnanensisEngl.),共17个种。
1.2 方法
分别采集各魔芋种地下茎、花器官形态、叶片形态、结实情况和实生籽图片以及花芽不同生长时期图片。全套样品分别于2012年、2013年和2014年生长季3次取样观察。形态指标观察记录及赋值如下。
1.2.1 地下茎 A1:魔芋地下茎形状。伸长的赋值为0;近球形赋值为1;碟形赋值为2。A2:地下茎芽点分布。梅花形分布赋值为0;螺旋形分布赋值为1。A3:地下茎。无匍匐茎赋值为0;具匍匐茎赋值为1。A4:地下茎根系。仅从主芽处生长赋值为0;从各个部位生长赋值为1。
1.2.2 叶 B1:发芽时地上部分叶的发生。具单叶赋值为0;具复合叶赋值为1。B2:花与叶出现时间。先花后叶赋值为0;同时发生赋值为1。B3:花柄与叶柄相比。花柄短于叶柄赋值为0;花柄与叶柄等长赋值为1。B4:鳞片叶与叶柄相比。鳞片叶与叶柄等长赋值为0;鳞片叶短于叶柄赋值为1。B5:叶柄或花柄。光滑赋值为0;表面有疣粒赋值为1。B6:叶的对称性。两侧对称赋值为0;辐射对称赋值为1。B7:第一叶。具1~3小叶赋值为0;具5小叶赋值为1;具7小叶赋值为2;具9小叶赋值为3。B8:小叶。长3倍于宽赋值为0;长2倍于宽赋值为1;长1~1.5倍于宽赋值为2。B9:中心小叶。具叶柄赋值为0;不具叶柄赋值为1。B10:侧边小叶。所有小叶具叶柄赋值为0;有的小叶无叶柄赋值为1。B11:叶腋珠芽球。无珠芽球赋值为0;有珠芽球赋值为1。
1.2.3 花器官 C1:花管。接近嘴部收缩赋值为0;接近嘴部不收缩赋值为1。C2:花管内表面。光滑赋值为0;有棱纹赋值为1;有薄层赋值为2。C3:花嘴。无卷曲或轻微卷曲赋值为0;具宽的卷曲赋值为1。C4:佛焰苞片与肉穗花序(含附属器)相比。佛焰苞短于肉穗花序赋值为0;佛焰苞长于肉穗花序赋值为1。C5:佛焰苞片。戴头巾状的赋值为0;不戴头巾状的赋值为1。C6:佛焰苞片。部分或完全地呈网纹赋值为0;无网纹赋值为1。C7:佛焰苞片。具平条纹赋值为0;具斑点赋值为1。C8:佛焰苞片。基部具白色斑点(块)赋值为0;基部无斑点(块)赋值为1。C9:佛焰苞片。有深色的过渡带赋值为0;无深色的过渡带赋值为1。C10:佛焰苞片。尖状赋值为0;钝形赋值为1。C11:肉穗花序。附属物顶部鞭状/锥形赋值为0;附属物顶部钝形赋值为1。C12:肉穗花序。附属物光滑赋值为0;附属物具皱纹赋值为1;附属物多刺的赋值为2。C13:肉穗花序。具柄的赋值为0;无柄赋值为1。C14:花管内表面。光滑赋值为0;有皱纹赋值为1。C15:胎座上部。胎座与附属器连接处有缩短的末端赋值为0;胎座与附属器连接处没有缩短的末端赋值为1。C16:胎座雄蕊上部。无中性物质赋值为0;具中性物质赋值为1。
1.2.4 果实 D1:顶部浆果扁平赋值为0;圆形赋值为1。D2:浆果颜色红色赋值为0;蓝色赋值为1。
1.3 数据分析
分别以地下茎的4个指标(A1、A2、A3、A4)、叶片的11个指标(B1、B2、…、B10、B11)、花器官的16个指标(C1、C2、…、C15、C16)和果实的2个指标(D1、D2)为变量,对17个种进行逐步聚类分析,根据聚类结果对种间距离相近的种进行判别分析。以参试17个魔芋种4类植物器官33个性状参数构成16×4×33的数据矩阵进行判别分析,判别方法参照高惠璇的实用统计方法[23]。利用SAS 9.0统计软件进行判别分析。
2 结果与分析
2.1 地下茎结构参数聚类
图1聚类结果显示,台湾魔芋与其他种分开,其地下茎芽点呈螺旋形分布(图2-A);桂平魔芋和红河魔芋聚为一组,这2种魔芋地下茎伸长,但桂平魔芋地下茎呈链子状(图2-B),红河魔芋地下茎呈圆筒形匍匐状(图2-C);白魔芋、密毛魔芋、花魔芋、东亚魔芋、珠芽魔芋、南蛇棒、勐海魔芋、米勒魔芋、疣柄魔芋、东京魔芋、攸乐魔芋、田阳魔芋、西盟魔芋和滇魔芋聚为一组,这14种魔芋的地下茎都是近球形。白魔芋、田阳魔芋、红河魔芋、密毛魔芋、花魔芋和西盟魔芋具匍匐茎(表1)。白魔芋、桂平魔芋、台湾魔芋、密毛魔芋、东亚魔芋和花魔芋的根系仅从主芽处长出。


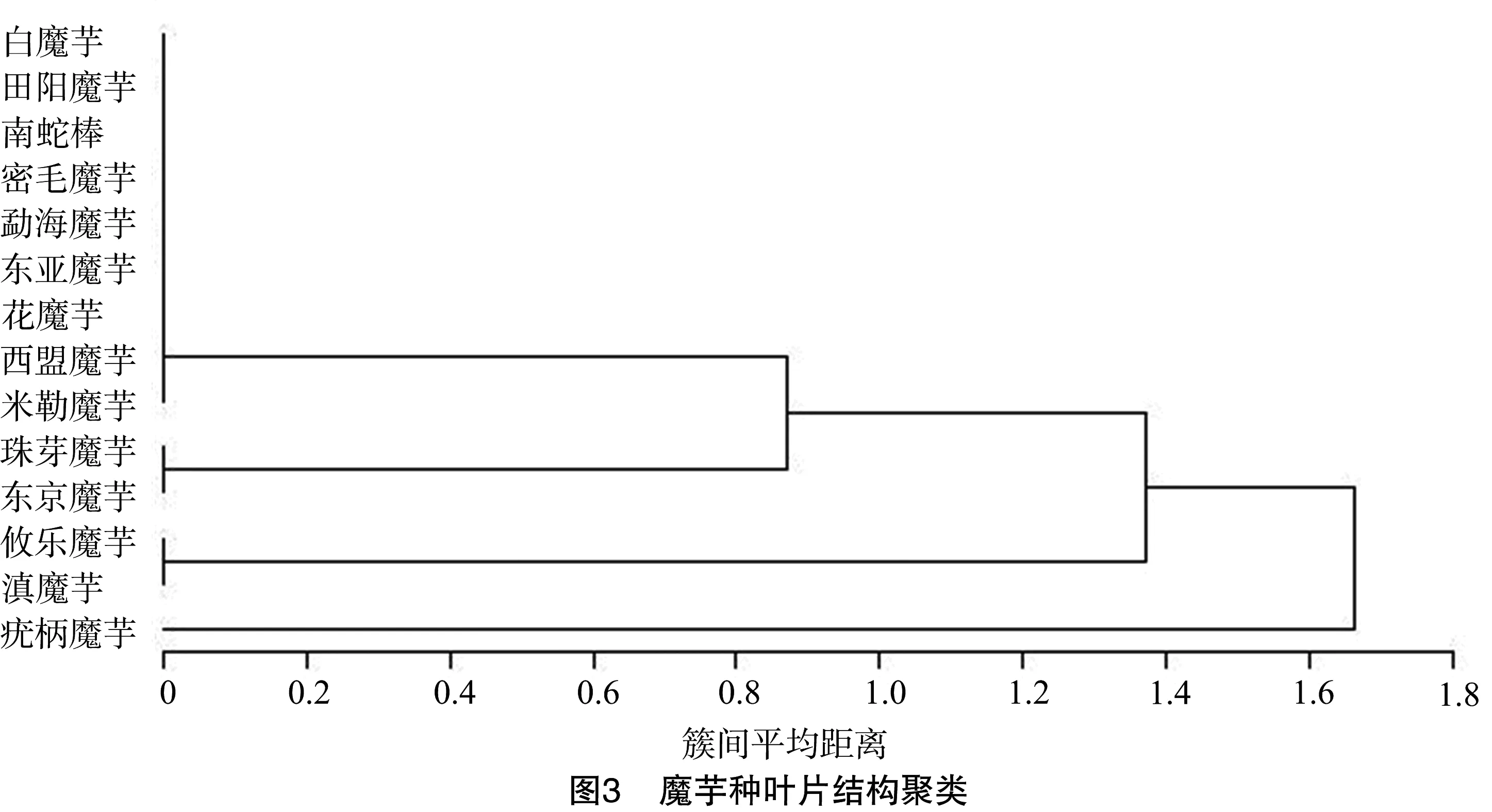
2.2 叶片结构参数聚类
图3聚类结果显示,疣柄魔芋单独分为一组,其叶柄表面具有疣粒(图4-A);东京魔芋和珠芽魔芋聚为一组,其叶腋长有珠芽(图4-B);攸乐魔芋和滇魔芋因其鳞片叶与叶柄等长聚为一组。发芽时桂平魔芋具复合叶,密毛魔芋花柄长度与叶柄长度近似,攸乐魔芋鳞片叶与叶柄等长,珠芽魔芋的叶辐射对称(表1)。
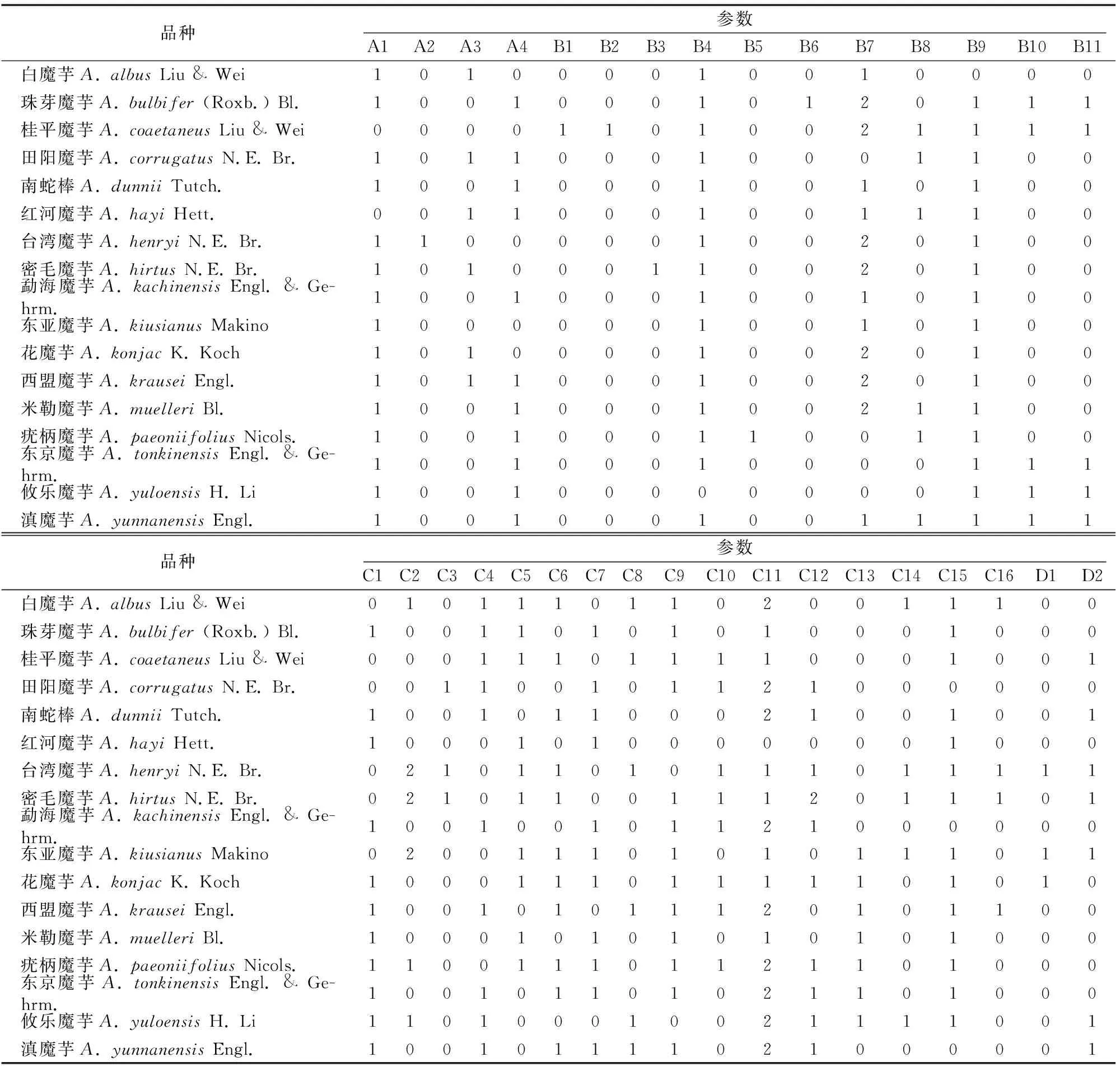
表1 17个魔芋种的33个结构特征

2.3 花器官形态
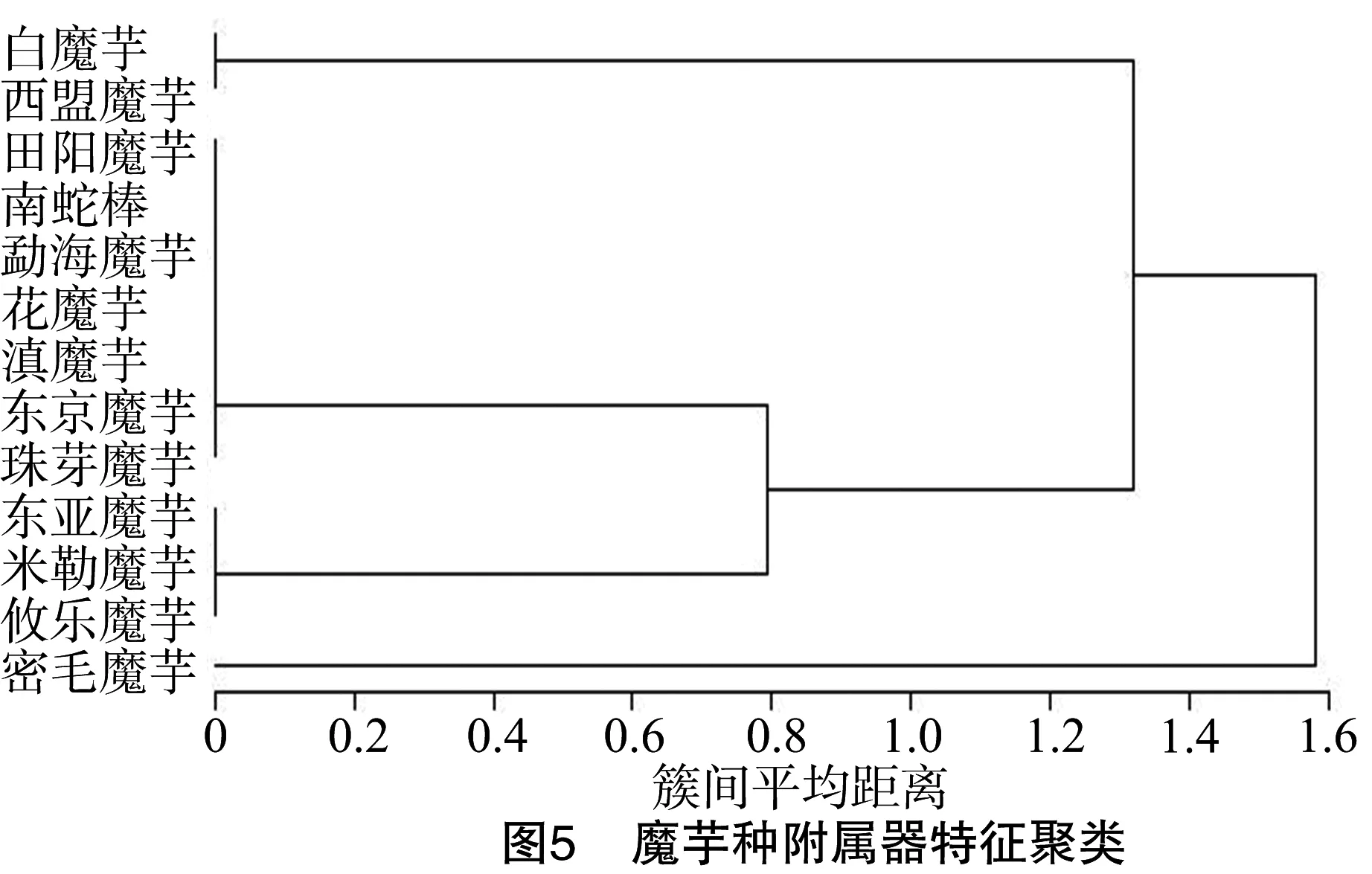
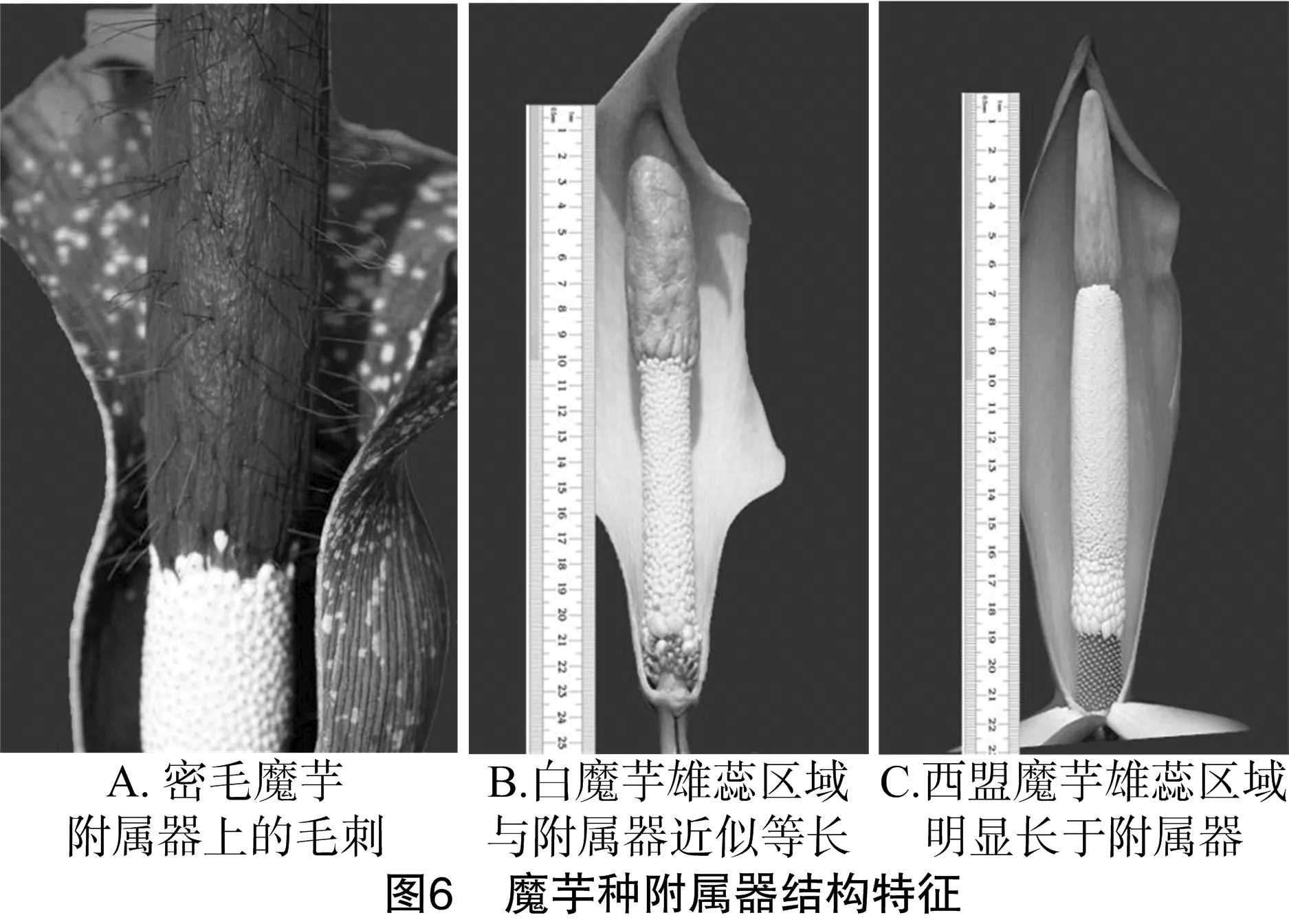
2.3.1 附属器结构特性聚类 图5将密毛魔芋单独聚为一类,因其附属器上长满密毛(图6-A);白魔芋和西盟魔芋聚为一组,这2种魔芋雄蕊和雌蕊之间有中性物质,但白魔芋雄蕊区长度近似于附属器长度(图6-B),而西盟魔芋雄蕊区长度明显比附属器的长度长(图6-C);其他的田阳魔芋、南蛇棒、勐海魔芋、花魔芋、滇魔芋、东京魔芋和珠芽魔芋聚为一组,因其附属器具皱纹;而附属器光滑的东亚魔芋、米勒魔芋和攸乐魔芋聚为一组(表1)。
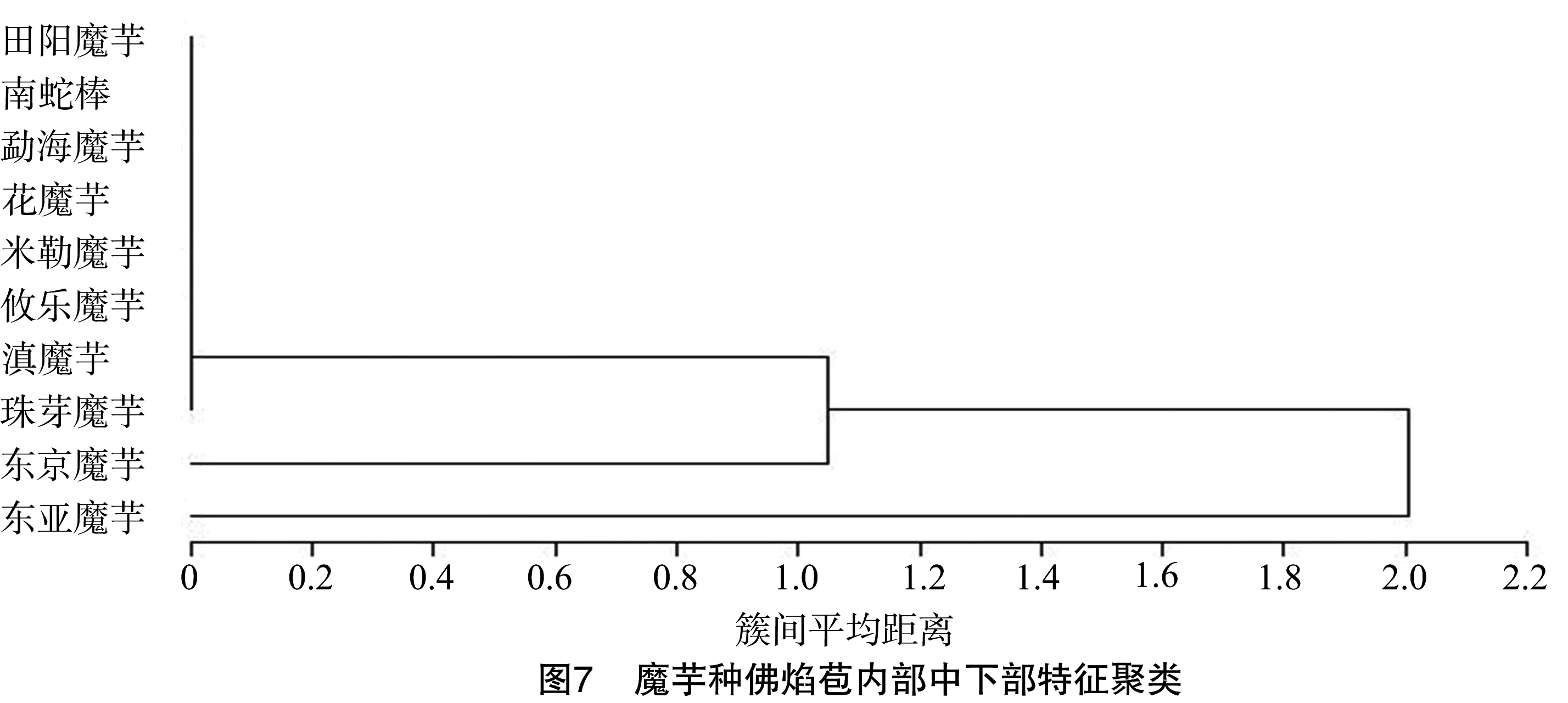
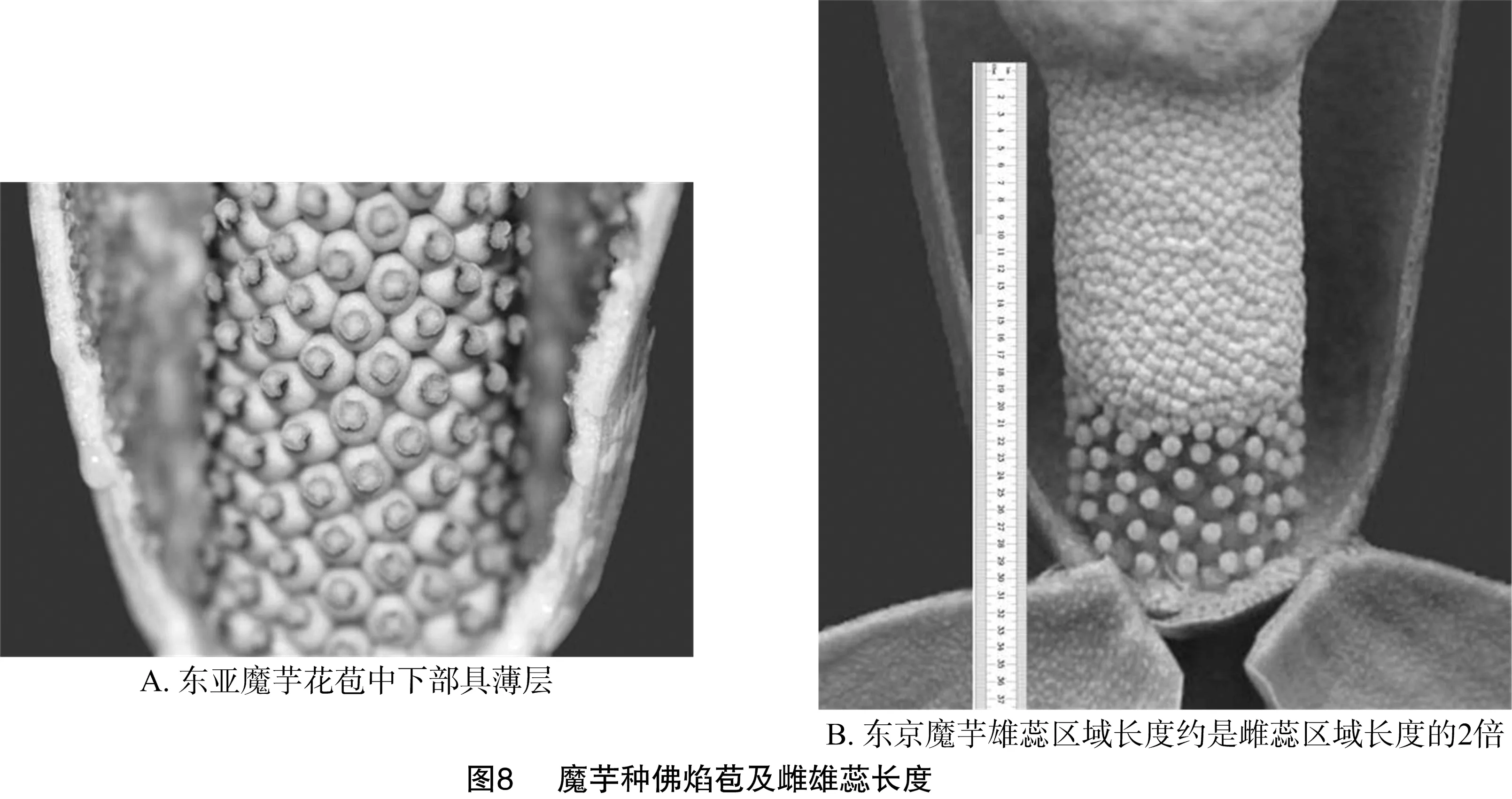
2.3.2 佛焰苞内部中下部特性聚类 从图7可以看出,东亚魔芋单独聚为一组,因其佛焰苞内部中下部有薄层(图8-A),观察到台湾魔芋、密毛魔芋也有类似的薄层(表1);东京魔芋聚为一组,其雄蕊区域长度约是雌蕊区域长度的2倍(图8-B)。田阳魔芋、台湾魔芋和密毛魔芋花嘴处有宽的卷曲(表1)。
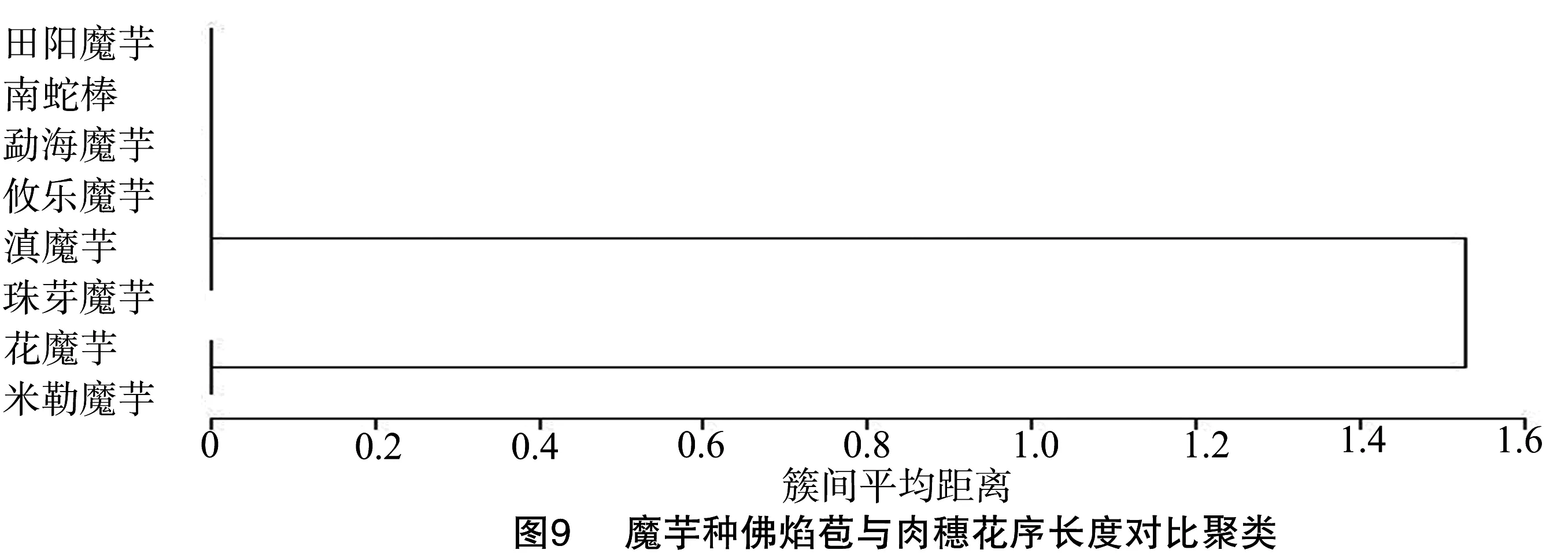
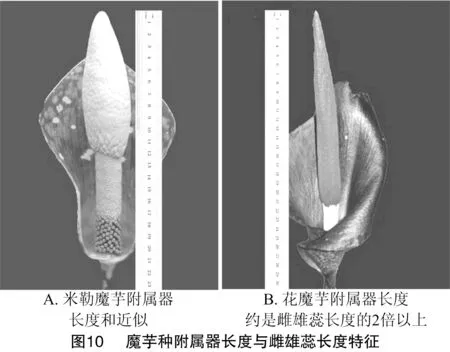
2.3.3 佛焰苞与肉穗花序(含附属器)对比聚类 图9显示花魔芋和米勒魔芋聚为一类,因其花序(含附属器)长于佛焰苞。台湾魔芋、密毛魔芋、东亚魔芋、疣柄魔芋和红河魔芋的花序(含附属器)也长于佛焰苞(表1)。米勒魔芋附属器长度与雌雄蕊长度和近似(图10-A),花魔芋附属器长度是雌雄蕊长度的2倍以上(图10-B)。
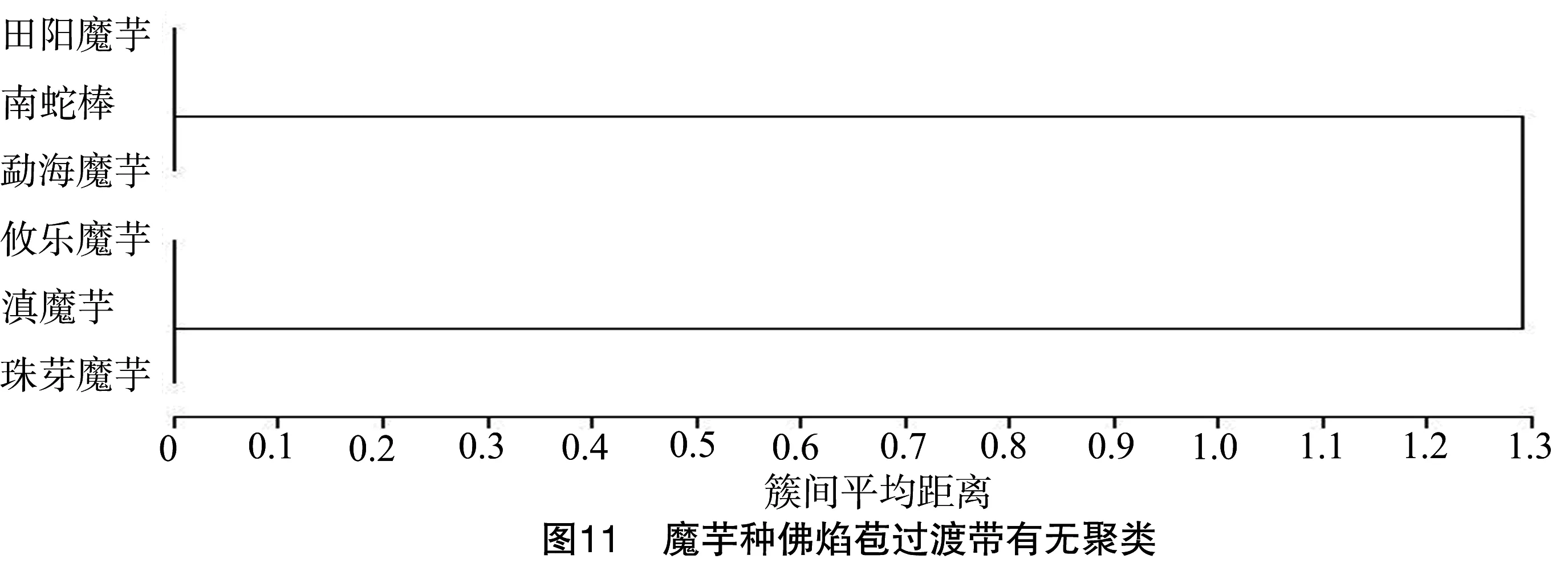
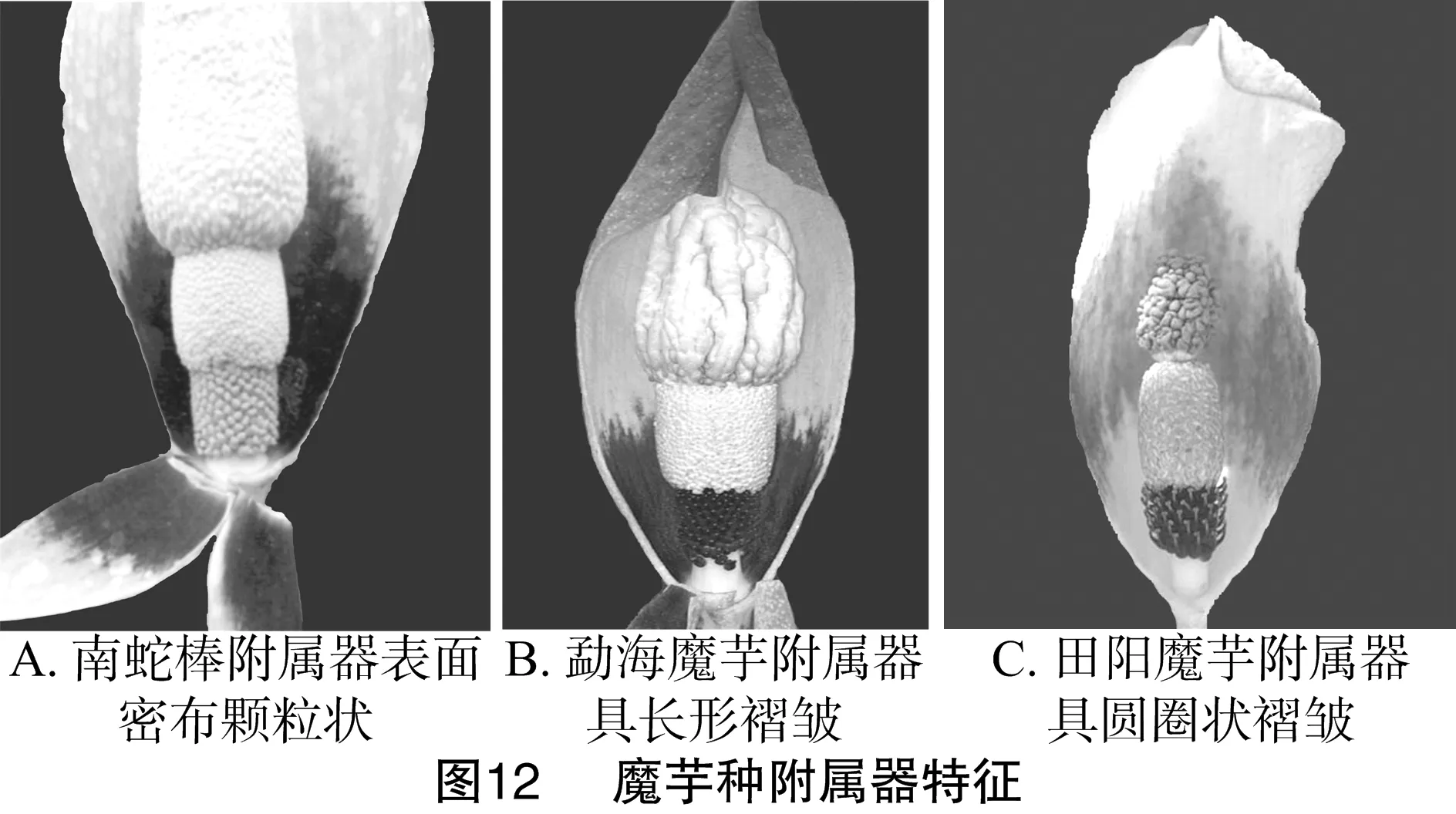
2.3.4 佛焰苞过渡带有无聚类 从图11可以看出,南蛇棒、勐海魔芋和田阳魔芋聚为一组,因其佛焰苞有过渡带(表1)。南蛇棒附属器表面密布颗粒状物质(图12-A),勐海魔芋附属器表面具明显条形皱褶(图12-B),田阳魔芋附属器密布圆圈状褶皱(图12-C);南蛇棒和勐海魔芋过渡带延伸至佛焰苞底部,而田阳魔芋过渡条带仅位于佛焰苞中部(图12-C)。
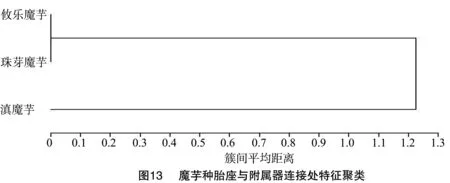
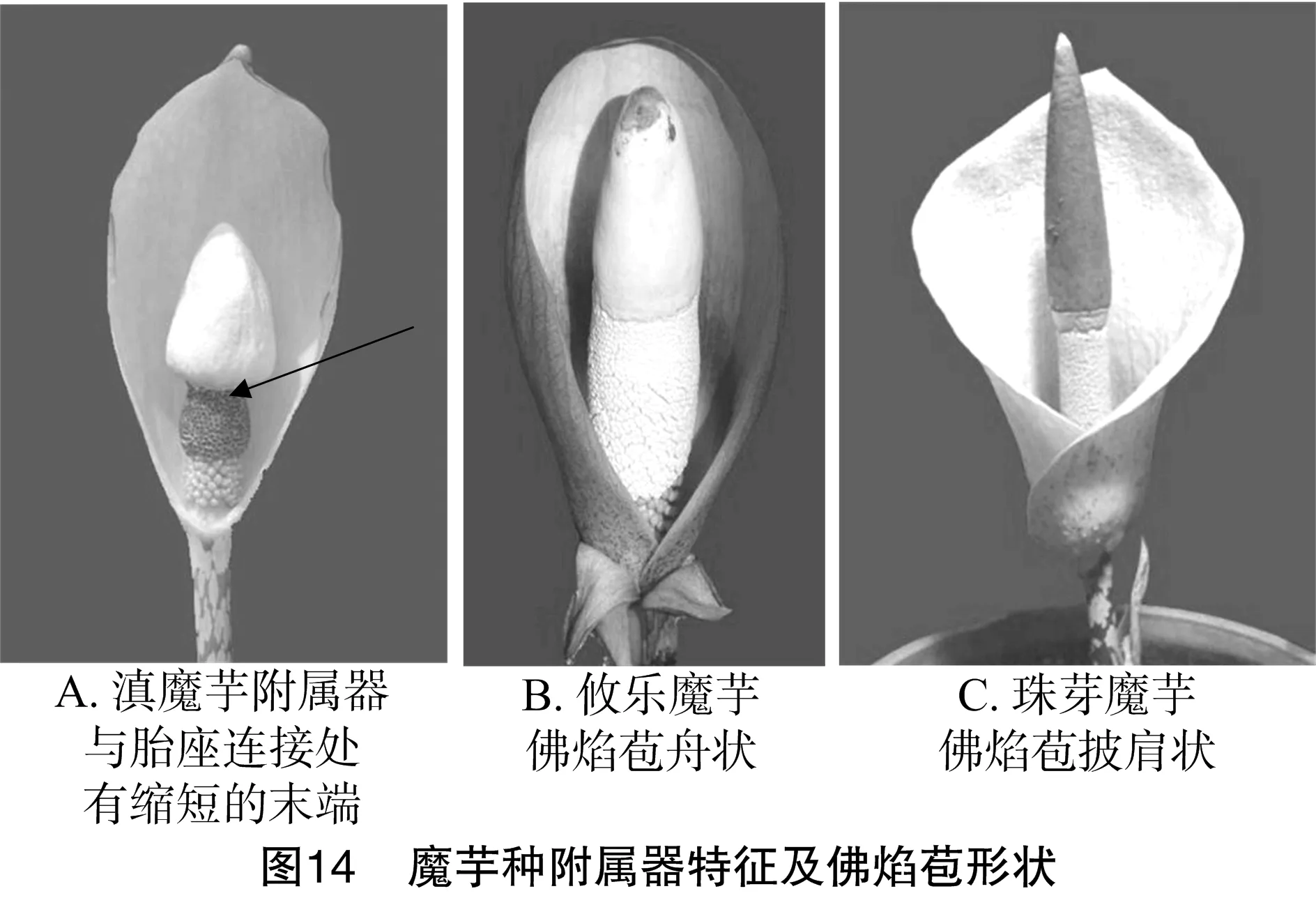
2.3.5 胎座与附属器连接处缩短的末端有无聚类 从图13可以看出,滇魔芋单独聚为一组,因其附属器与胎座连接处有缩短的末端(图14-A箭头所示区域)。珠芽魔芋和攸乐魔芋聚为一组,但两者的佛焰苞形状不同,攸乐魔芋佛焰苞呈舟状(图14-B),而珠芽魔芋佛焰苞呈披肩形(图14-C)。
3 讨论
许多学者从形态学[23-26]、细胞学[27]、分子生物学[28]等多个角度对魔芋种间亲缘关系进行了研究,但田间鉴定难度还是很大。魔芋形态指标图文展示结合聚类方法,可以增加魔芋田间鉴别速度和鉴别准确度。植物的表型是基因型和环境因素共同作用的结果,涉及大量质量性状和数量性状。本研究结合魔芋的关键性状,将质量性状转化为二元形态性状,结合照片取证识别特征,可以快速识别魔芋种类。这种方法已经被成功应用于葡萄[29]和菊花[30]。传统分类学根据形态学特征对魔芋进行区分,无疑是最准确的,但文字表述不同人的理解之间是有差距的,由传统分类学与统计学及计算机技术结合产生的数量分类学方法,使基于形态学特征的研究更加高效。
在辨别魔芋地下球茎时,根据形状、芽点分布、匍匐茎有无及根系着生情况可以将魔芋分为几个类别,这与Lamprecht等用系统分类法得到的结果[3-6,31-32]是一致的。分类后,球形地下茎螺旋状芽点分布的台湾魔芋很容易就能跟其他种区别开来。链子状地下茎的桂平魔芋和筒形匍匐状的红河魔芋地下茎也很容易鉴别出来。
从叶片特征看,疣柄魔芋特殊的疣粒状叶柄,是其不同于其他种的显著特征。叶片的对称性在小苗期间可以辨别珠芽魔芋,随着叶片复叶的增多,其对称性的辨别难度加大。叶柄颜色和叶柄斑点在不同时期会有明显变化[33-35],不同的栽培条件下叶柄颜色和斑点也会发生变化。因此,据叶片特征来辨别魔芋种难度很大。
花器官是鉴别魔芋种的重要特征。魔芋的花器官包括佛焰苞和肉穗花序,肉穗花序包括附属器、雄蕊和雌蕊,部分魔芋种雄蕊和雌蕊区域间有中性物质[36-37],如白魔芋、台湾魔芋、密毛魔芋、西盟魔芋和东京魔芋。附属器长度与佛焰苞长度对比,附属器长度与雌雄蕊区域长度对比,都是识别魔芋种的重要特征,可以快速区别出白魔芋、西盟魔芋、米勒魔芋和花魔芋。密毛魔芋附属器上密布的毛刺则成了识别该种的一个显著特征[38]。佛焰苞内部条纹和薄层、附属器皱纹样式,也是区分魔芋种的重要特征,可以快速区分东亚魔芋、东京魔芋、南蛇棒、勐海魔芋、滇魔芋和田阳魔芋。佛焰苞的形状是区别滇魔芋与攸乐魔芋的重要特征,我国分布的滇魔芋和攸乐魔芋2个种的佛焰苞都是舟形的,但是可以通过附属器与胎座连接处是否有缩短的末端来区别这2个魔芋种,滇魔芋附属器与胎座连接处有缩短的末端,而攸乐魔芋没有。
致谢:感谢荷兰Von Gimborn Arboretum的Hetterscheid教授提供部分图片,感谢中国科学院昆明植物研究所李恒研究员对魔芋研究工作的指导,感谢云南临沧滇晟农林科技有限公司吴昊林和李永强提供研究材料、种植场地并对魔芋种的栽培管护。
参考文献:
[1]李 恒,龙春林. 中国魔芋属的分类问题[J]. 云南植物研究,1998,20(2):167-170.
[2]Hetterscheid W L A,Ittenbach S,Bogner J,et al. Notes on the genusAmorphophallus(Araceae)10.Revision of the endemicAmorphophallusspecies of Madagascar[J]. Bot Jahrb Syst,1999,121(1):1-17.
[3]Lamprecht I,Seymour R S. Thermologic investigations of three species ofAmorphophallus[J]. Journal of Thermal Analysis and Calorimetry,2010,102(1):127-136.
[4]李 川,王 军,崔 鸣. 魔芋属植物开花的研究进展[J]. 中国农学通报,2011,27(10):73-75.
[5]白立伟,牛 义,刘海利,等. 魔芋种质资源及育种研究进展[J]. 南方农业,2016,10(4):48-52.
[6]Li H,Hetterscheid W L A.Amorphophallus[M]// Flora of China.Science Press,Missouri Botanical Garden Press,2010:23-33.
[7]Hetterscheid W L A,Van der Ham R. Notes on the genusAmorphophallus(Araceae)-11. New and obsolete species from East Malaysia and continental Southeast Asia[J]. Blumea,2001,46(2):253-282.
[8]龙德清,刘传银,朱圣平. 魔芋的开发利用与研究进展[J]. 食品科技,2003(11):18-20.
[9]蒋学宽,刘海利,牛 义,等. 4种魔芋资源的裂片及其表皮细胞的形态学比较研究[J]. 西南大学学报(自然科学版),2012,34(4):53-57.
[10]张风洁,刘海利,张 洁,等. 珠芽魔芋资源研究进展[J]. 南方农业,2013,7(8):64-67.
[11]牛 义,张盛林,刘好霞,等. 不同倍性白魔芋植株的细胞学形态观察与核型分析[J]. 西南大学学报(自然科学版),2008,30(12):79-84.
[12]郭 晟. 三种魔芋葡甘聚糖不同特性比较研究[D]. 长沙:湖南农业大学,2012:36-39.
[13]刘佩瑛. 魔芋学[M]. 北京:农业出版社,2004:75-81.
[14]张盛林,牛 义,刘佩瑛. 中国魔芋粉市场回顾与展望[J]. 中国食品添加剂,2006(3):78-81.
[15]张东华,汪庆平. 缅甸魔芋(AmorphophallusMuelleri)资源分布及产业基本概况[J]. 热带农业科学,2013,33(4):46-51.
[16]张东华,汪庆平. 中国魔芋种植业如何应对日本重返缅甸[J]. 热带农业科学,2015,35(7):76-80.
[17]刘 楠,杨 芳. 魔芋葡甘聚糖的研究进展及应用现状综述[J]. 安康学院学报,2011,23(4):95-98.
[18]吴金平,宋志红,刁 英,等. 珠芽魔芋的组织培养与快速繁殖[J]. 植物生理学通讯,2007,43(5):887-888.
[19]李松林,崔 熙,谢学键,等. 魔芋属7种植物中魔芋葡甘露聚糖的检测[J]. 中国中药杂志,1996,21(8):456-458,509.
[20]张玉进,张兴国,刘佩瑛,等. 魔芋种质资源的RAPD分析[J]. 西南农业大学学报,2001,23(5):418-421.
[21]滕彩珠. 魔芋DNA分子标记技术及其初步应用[D]. 桂林:广西师范大学,2008:41-48.
[22]蒋学宽. 魔芋新品种DUS测试指南制定研究[D]. 重庆:西南大学,2012:30-38.
[23]刘佩瑛,陈劲枫. 魔芋属一新种[J]. 西南农学院学报,1984(1):67-69.
[24]刘寿养,韦松基. 广西魔芋属一新种[J]. 广西植物,1986,6(3):183-186.
[25]李 恒,龙春林. 药用植物魔芋的考证[J]. 天然产物研究与开发,1989,1(2):87-92.
[26]Hetterscheid W L A. Notes on the genusAmorphophallus(Araceae)-2. New species from Tropical Asia[J]. Blumea,1994,39:237-281.
[27]宋学华,戴志凌,崔 熙. 六种魔芋属植物块茎的显微鉴定研究[J]. 中草药,1997,28(9):554-558.
[28]滕彩珠,刁 英,常福浩森,等. 云南魔芋种质资源亲缘关系的ISSR分析[J]. 安徽农学通报,2006,12(11):54-56.
[29]祁栋灵,周庆阳,刘三军,等. 利用叶形结构数值分析葡萄种质亲缘关系的研究[J]. 中国南方果树,2005,34(3):64-66.
[30]王江民,陈发棣,房伟民,等. 基于叶形特征的切花菊品种鉴别[J]. 植物学报,2013,48(6):608-615.
[31]Hetterscheid W L A,Peng C I. Notes on the genusAmorphophallus(Araceae). IV. Revision of the species in Taiwan[J]. Botanical Bulletin-Academia Sinica Taipei,1995,36:101-112.
[32]Hetterscheid W L A,Ltterlands S. Everything you always wanted to know aboutAmorphophallus,but were afraid to stick your nose into!!![J]. Aroideana,1996,19:7-13.
[33]Barthlott W,Bogner J. Rediscovery ofAmorphophallusstaudtii(Engl.)N.E.Br. in the Tai National Park,Ivory Coast[J]. Aroideana,1981,4(4):109-113.
[34]Li H,Long C L. New taxa ofAmorphophallusfrom China[J]. Aroideana,1989,11(1):4-9.
[35]Ittenbach S. Revision der afrikanischen Arten der GattungAmorphophallus(Araceae)[J]. Englera,2003(25):134-139.
[36]van der Ham R,Grob G,Hetterscheid W,et al. Notes on the genusAmorphophallus(Araceae)-13. Evolution of pollen ornamentation and ultrastructure inAmorphophallusandPseudodracontium[J]. Grana,2005,44(4):252-265.
[37]Punekar S A,Kumaran K P N. Pollen morphology and pollination ecology ofAmorphophallusspecies from North Western Ghats and Konkan region of India[J]. Flora - Morphology,Distribution,Functional Ecology of Plants,2010,205(5):326-336.
[38]Hetterscheid W L A,Yadav S R,Patil K S. Notes on the genusAmorphophallus(Araceae)-5.Amorphophalluskonkanensis,a new species from India,and taxonomic reflections onAmorphophallussectionRhaphiophallus[J]. Blumea,1994,39:289-294.
猜你喜欢
贵州农业科学(2023年10期)2023-10-27
红蜻蜓·高年级(2022年10期)2022-12-31
小天使·四年级语数英综合(2022年6期)2022-06-21
农产品加工(2022年4期)2022-03-11
草地学报(2021年6期)2021-07-05
作文小学中年级(2021年4期)2021-05-07
特种经济动植物(2021年2期)2021-02-25
材料科学与工程学报(2016年1期)2017-01-15
食品科学(2013年17期)2013-03-11
食品科学(2013年14期)2013-03-11
