
桑树潜在致敏原鉴定与生物信息学分析
2018-06-29 04:58韩利红宋培兵张亚萍唐利洲
江苏农业科学 2018年11期
刘 潮, 韩利红, 宋培兵, 张亚萍, 唐利洲
(曲靖师范学院云南高原生物资源保护与利用研究中心/云南省高校云贵高原动植物多样性及生态适应性进化重点实验室,云南曲靖 655011)
过敏属于世界性的健康问题,植物来源的过敏原能引起人类的Ⅰ型过敏反应,人们既可以从上呼吸道吸入,也可以通过存在于种类繁多的植物食品中的过敏原引起食物过敏反应。近年来,食物过敏的患病率有所上升,在西方国家中,多达8%的幼儿和2%的成人受到影响[1]。桑树是一种常见的落叶树种,其叶片多汁,是桑蚕的主要饲料,曾有报道指出,桑树花粉能引起呼吸过敏反应和接触性荨麻疹[2]。桑树的果实桑葚能直接食用或用作酿酒原料,也是常用的中草药成分,过敏者食用桑葚能引发全身性荨麻疹,导致呼吸困难、胸闷和腹绞痛等[3]。
致敏原属于多个蛋白家族,结构复杂、功能多样。编码序列和致敏原的三维(three dimensional,简称3D)结构对引起致敏性至关重要。大多数过敏原都比较小、稳定、结构良好,这些蛋白质家族多为种子贮藏蛋白、结构蛋白和防御相关的蛋白质——病程相关蛋白。序列或结构相似的蛋白容易引发免疫球蛋白E(IgE)交叉反应,可以通过电脑模拟的方法预测其致敏性。计算工具能相当精确和快速地预测变应原分子的表位位置,并将其用于确定分子结构,从而确定B细胞表位[4]。在本研究中,筛选了63个桑树潜在致敏原,分为13个家族。这些潜在的抗原表位的IgE结合残基的功能研究将为后续研究提供借鉴。
1 材料与方法
1.1 致敏原蛋白序列的获取
2016年10月,笔者通过食物过敏研究和资源计划(FARRP)的致敏原蛋白质数据库(www.allergenonline.org)和致敏原命名委员会官方网站(http://www.allergen.org/index.php),下载了截至2016年8月4日公布的705条植物和真菌致敏原蛋白序列数据。
1.2 桑树潜在致敏原的筛选
以致敏蛋白全序列为探针,通过Blast桑树(Morusnotabilis)蛋白数据库,根据序列间的相似性筛选潜在的桑树致敏蛋白。桑树潜在致敏蛋白选择阈值为序列一致性(identity)≥60%[5]和E值≤10-20(www.allergenonline.org)。
1.3 桑树潜在致敏原的结构和功能分析
利用UniProt数据库(http://www.uniprot.org/)对筛选的潜在致敏原蛋白进行结构域和基因本体(gene ontology,简称GO)功能分类分析。利用WEGO在线软件(Web Gene Ontology Annotation Plotting,http://wego.genomics.org.cn/)对蛋白GO富集度进行计算。
1.4 桑树潜在致敏原的三级结构预测
蛋白同源三级结构模型用Swiss-Model[6]构建完成。3D结构的可视化使用VMD 1.8.5 (http://www.ks.uiuc.edu/Research/vmd/vmd-1.8.5/)完成。
2 结果与分析
2.1 桑树潜在致敏原的筛选
以从FARRP和Allergome数据库上下载的致敏原蛋白为探针,BLAST桑树蛋白数据库,以序列一致性(identity)≥60%和E值≤10-20为筛选条件,共获得143条桑树潜在致敏原(其中4条注释为Allergen的序列低于筛选条件),详见表1。
2.2 桑树潜在致敏原的功能分类
GO注释将基因产物分为细胞组分、分子功能和生物过程3类,可以对基因及其蛋白的功能进行分类以了解其功能。在143条桑树潜在致敏原中,有113条具有GO功能注释,分析发现71条不同的GO注释子条目,7类GO子分类基因作为细胞结构组分起作用,其中参与细胞和细胞部分组成的均占46.4%,参与细胞器结构成分的占32.1%。在分子功能分类中,具有结合功能的蛋白占67.9%,具有催化活性的蛋白占25.0%,具有结构分子活性的蛋白占14.3%。13类GO子分类基因参与了生物学过程,其中参与细胞过程的占39.3%,参与代谢过程的占28.6%,参与应激反应的占23.2%(图1)。
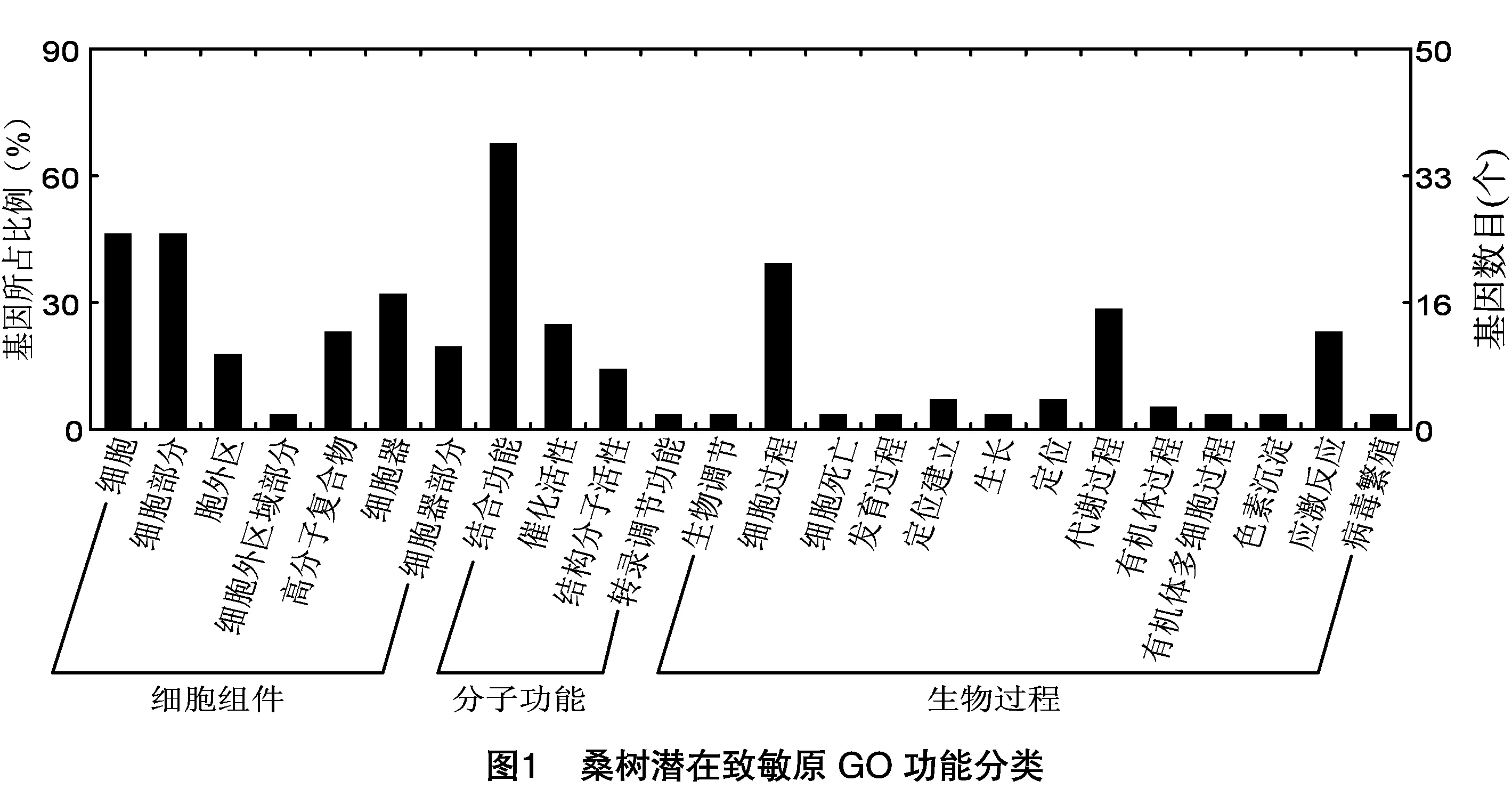
2.3 桑树潜在致敏原的家族分布
在143条桑树潜在致敏原中共查询到172个pfam保守功能域,分属于49个pfam家族(图2),根据其所属pfam家族共分为22类(表1)。其中热休克蛋白家族含有HSP70(PF00012)、HSP20(PF00011)和HSP90(PF00183)3类结构域,共有25条序列,该类蛋白是蛋白质折叠复合体的重要成分,可保护植物免受胁迫的损伤[7],关于该家族成员致敏原活性的报道较少。含有Profilin(PF00235)功能域的有3条序列,且与已知致敏原序列一致性较高,该类蛋白在调节肌动蛋白聚合中起作用,在多种植物的花粉、果实或种子中具有致敏原活性。肽基脯氨酰顺反异构酶(Pro_isomerase,编号PF00160),也称为亲环素(cyclophilin),在哺乳动物中是免疫抑制剂环孢素A(CsA)的主要靶细胞组分,也具有分子伴侣和细胞信号转导的功能,桑树潜在致敏原中有12个成员含有Pro_isomerase功能域。类甜蛋白(thaumatin-like protein,简称TLP)是一类能响应胁迫的蛋白家族,多种植物水果或花粉的TLPs具有致敏原活性[8],桑树有4个潜在的TLP致敏原成员。捕光色素A-B结合蛋白在植物光合作用中发挥重要作用,在芹菜中的Api g 3具有致敏原活性[9],桑树潜在致敏原中有5个成员具有Chloroa_b-bind(PF00504)功能域。
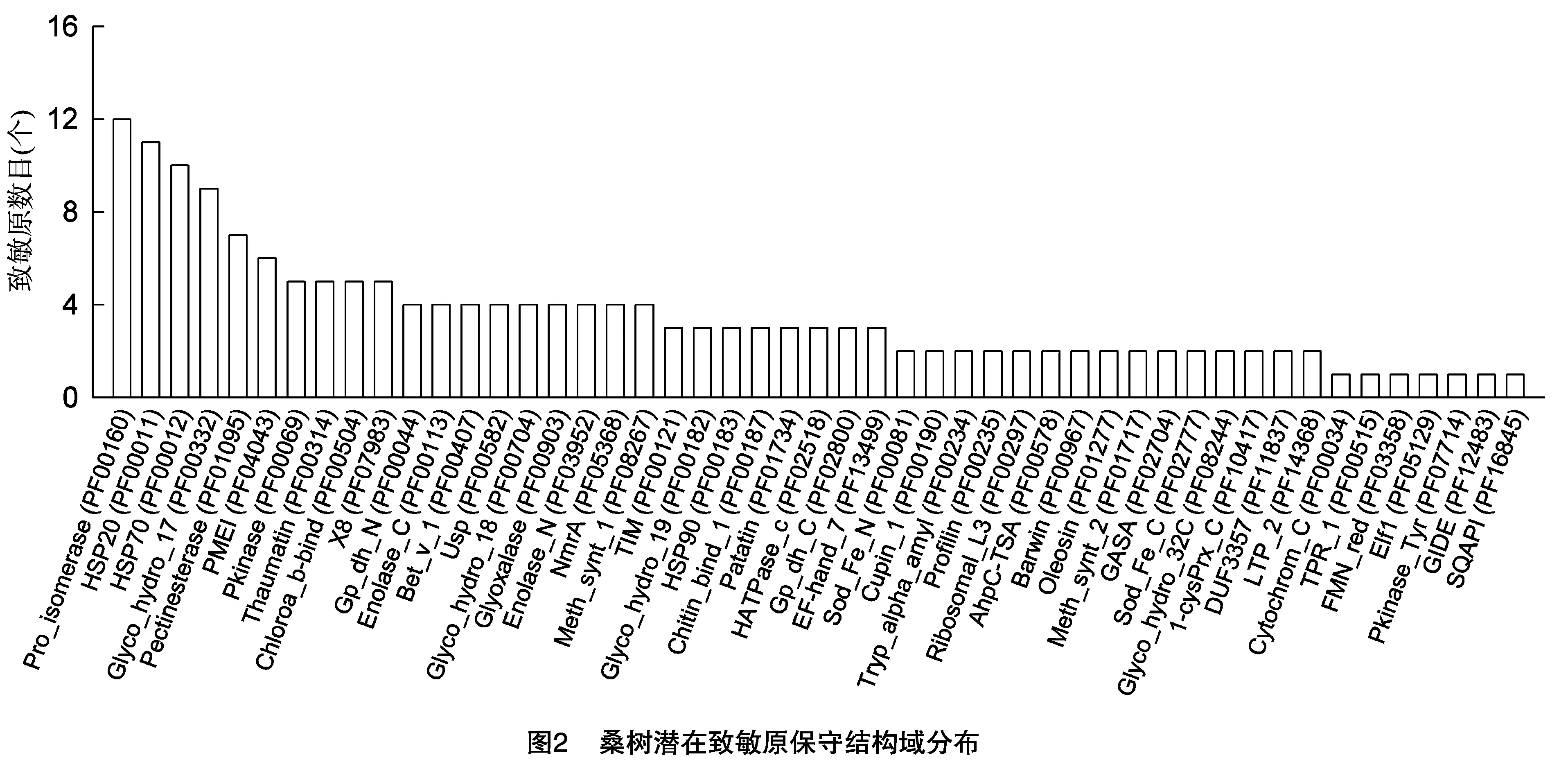
有部分序列同时含有多个功能域,如L484_024225同时含有HSP90(PF00183)和HATPase_c(PF02518)2个功能域,L484_020224同时含有Glyco_hydro_18(PF00704)和Thaumatin(PF00314)2个功能域,L484_006986则同时含有Glyco_hydro_17(PF00332)、Pkinase(PF00069)和Profilin(PF00235)3个功能域。
2.4 桑树潜在致敏原的结构和功能分析
蛋白初级结构是高级结构的基础,具有序列相似性的蛋白,其三维结构可能有明显不同,蛋白序列或结构,尤其是高级结构在其发挥功能过程中起决定作用,蛋白三维结构表层的氨基酸残基具有参与致敏活性的可能。Ⅰ型致敏原具有多样的生理生化特征、多样的初级结构和三维结构。蛋白质只有正确折叠,才具有生物学活性,为充分了解桑树潜在致敏原的致敏潜力,笔者以部分蛋白质为例,构建了蛋白质三维模型,以便更准确地了解蛋白质的致敏性。
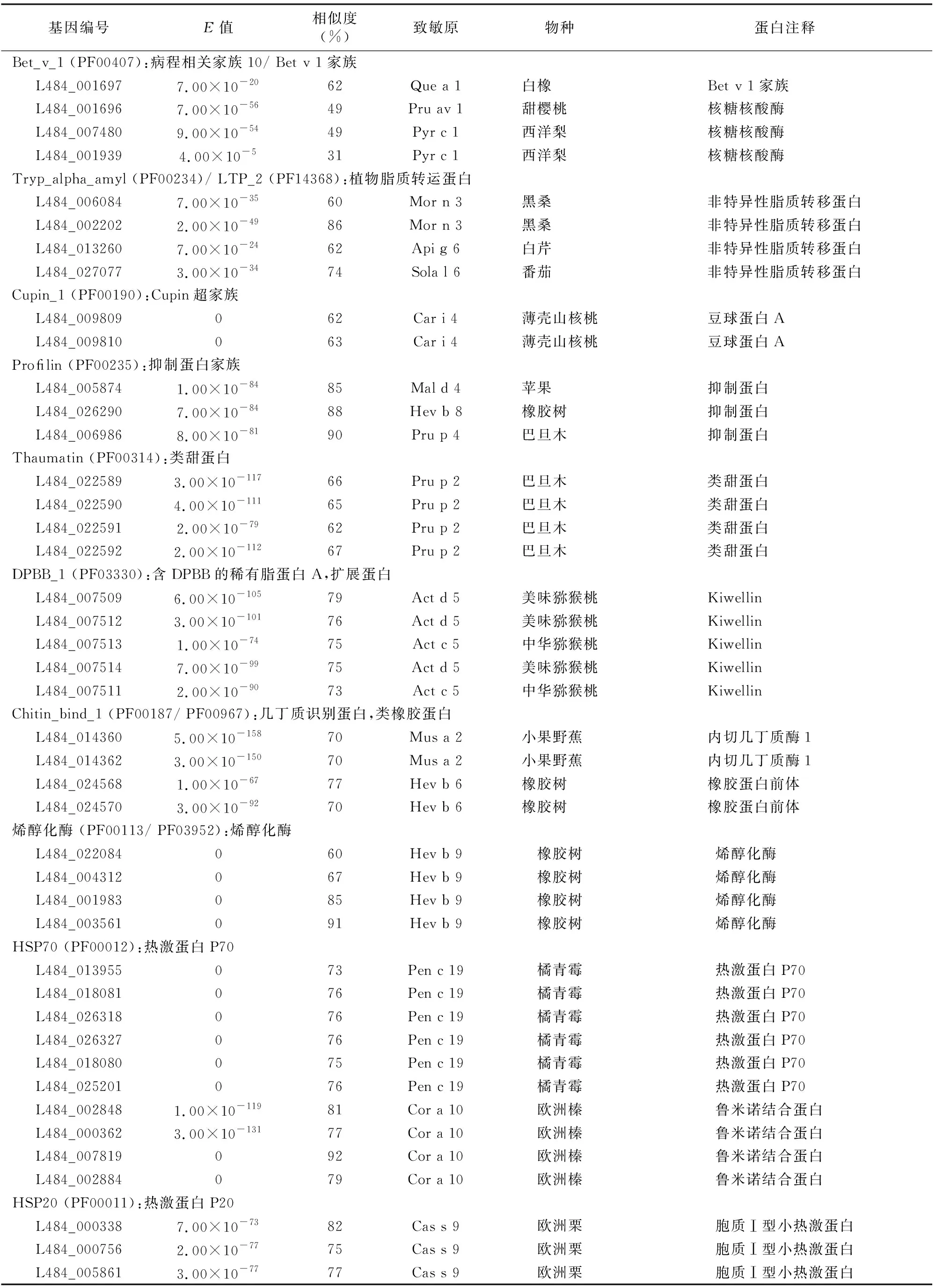
表1 桑树潜在致敏原蛋白信息
续表1
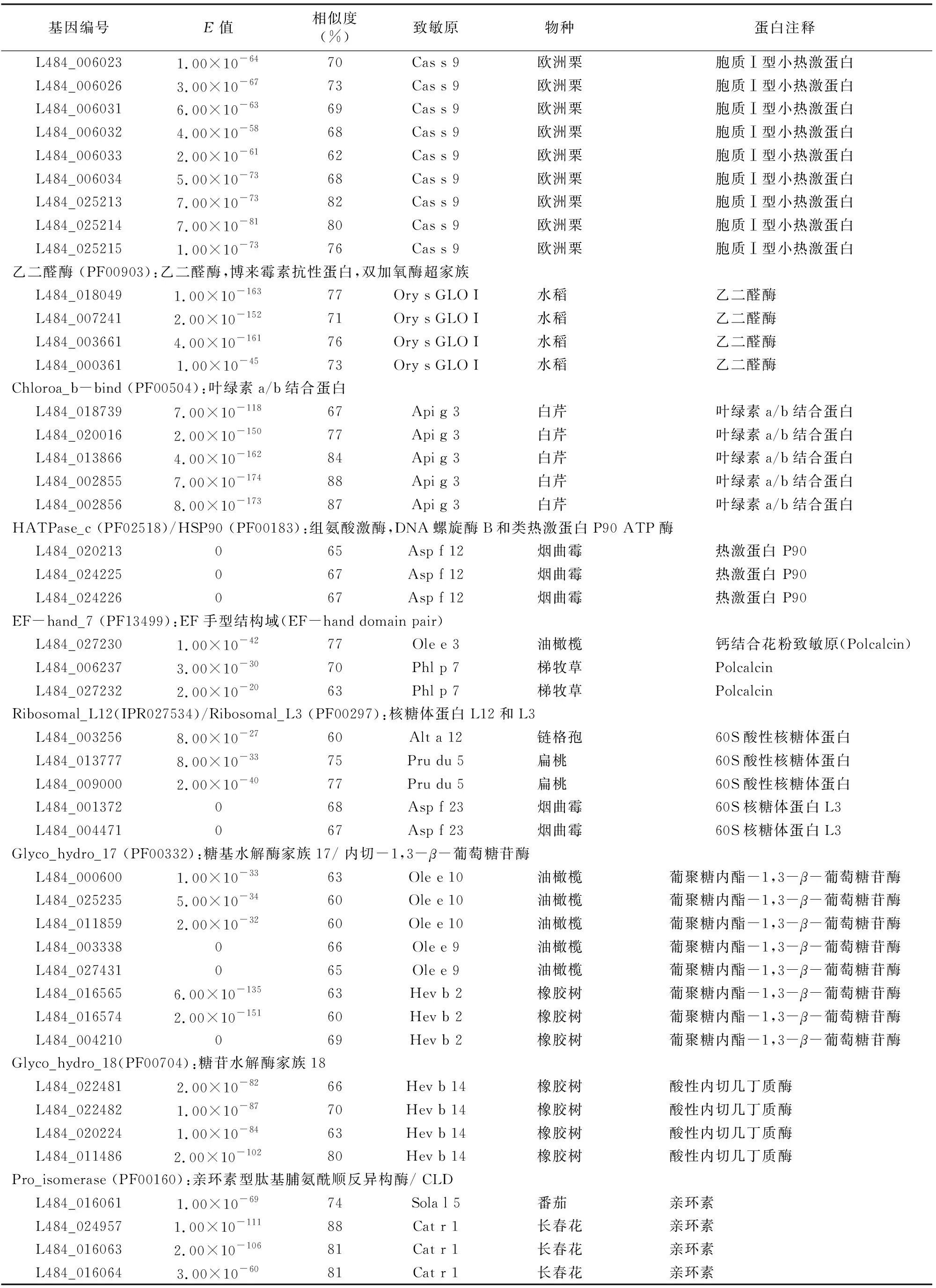
基因编号E值相似度(%)致敏原物种蛋白注释L484_0060231.00×10-6470Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0060263.00×10-6773Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0060316.00×10-6369Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0060324.00×10-5868Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0060332.00×10-6162Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0060345.00×10-7368Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0252137.00×10-7382Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0252147.00×10-8180Cas s 9欧洲栗胞质Ⅰ型小热激蛋白L484_0252151.00×10-7376Cas s 9欧洲栗胞质Ⅰ型小热激蛋白乙二醛酶 (PF00903):乙二醛酶,博来霉素抗性蛋白,双加氧酶超家族L484_0180491.00×10-16377Ory s GLO I水稻乙二醛酶L484_0072412.00×10-15271Ory s GLO I水稻乙二醛酶L484_0036614.00×10-16176Ory s GLO I水稻乙二醛酶L484_0003611.00×10-4573Ory s GLO I水稻乙二醛酶Chloroa_b-bind (PF00504):叶绿素a/b结合蛋白L484_0187397.00×10-11867Api g 3白芹叶绿素a/b结合蛋白L484_0200162.00×10-15077Api g 3白芹叶绿素a/b结合蛋白L484_0138664.00×10-16284Api g 3白芹叶绿素a/b结合蛋白L484_0028557.00×10-17488Api g 3白芹叶绿素a/b结合蛋白L484_0028568.00×10-17387Api g 3白芹叶绿素a/b结合蛋白HATPase_c (PF02518)/HSP90 (PF00183):组氨酸激酶,DNA螺旋酶B和类热激蛋白P90 ATP酶L484_020213065Asp f 12烟曲霉热激蛋白 P90L484_024225067Asp f 12烟曲霉热激蛋白 P90L484_024226067Asp f 12烟曲霉热激蛋白 P90EF-hand_7 (PF13499):EF手型结构域(EF-hand domain pair)L484_0272301.00×10-4277Ole e 3油橄榄钙结合花粉致敏原(Polcalcin)L484_0062373.00×10-3070Phl p 7梯牧草PolcalcinL484_0272322.00×10-2063Phl p 7梯牧草PolcalcinRibosomal_L12(IPR027534)/Ribosomal_L3 (PF00297):核糖体蛋白L12和L3L484_0032568.00×10-2760Alt a 12链格孢60S酸性核糖体蛋白L484_0137778.00×10-3375Pru du 5扁桃60S酸性核糖体蛋白L484_0090002.00×10-4077Pru du 5扁桃60S酸性核糖体蛋白L484_001372068Asp f 23烟曲霉60S核糖体蛋白L3L484_004471067Asp f 23烟曲霉60S核糖体蛋白L3Glyco_hydro_17 (PF00332):糖基水解酶家族17/ 内切-1,3-β-葡萄糖苷酶L484_0006001.00×10-3363Ole e 10油橄榄葡聚糖内酯-1,3-β-葡萄糖苷酶L484_0252355.00×10-3460Ole e 10油橄榄葡聚糖内酯-1,3-β-葡萄糖苷酶L484_0118592.00×10-3260Ole e 10油橄榄葡聚糖内酯-1,3-β-葡萄糖苷酶L484_003338066Ole e 9油橄榄葡聚糖内酯-1,3-β-葡萄糖苷酶L484_027431065Ole e 9油橄榄葡聚糖内酯-1,3-β-葡萄糖苷酶L484_0165656.00×10-13563Hev b 2橡胶树葡聚糖内酯-1,3-β-葡萄糖苷酶L484_0165742.00×10-15160Hev b 2橡胶树葡聚糖内酯-1,3-β-葡萄糖苷酶L484_004210069Hev b 2橡胶树葡聚糖内酯-1,3-β-葡萄糖苷酶Glyco_hydro_18(PF00704):糖苷水解酶家族18L484_0224812.00×10-8266Hev b 14橡胶树酸性内切几丁质酶L484_0224821.00×10-8770Hev b 14橡胶树酸性内切几丁质酶L484_0202241.00×10-8463Hev b 14橡胶树酸性内切几丁质酶L484_0114862.00×10-10280Hev b 14橡胶树酸性内切几丁质酶Pro_isomerase (PF00160):亲环素型肽基脯氨酰顺反异构酶/ CLDL484_0160611.00×10-6974Sola l 5番茄亲环素L484_0249571.00×10-11188Cat r 1长春花亲环素L484_0160632.00×10-10681Cat r 1长春花亲环素L484_0160643.00×10-6081Cat r 1长春花亲环素
续表1
基因编号E值相似度(%)致敏原物种蛋白注释L484_0138973.00×10-3662Asp f 27烟曲霉亲环素L484_0245325.00×10-9776Sola l 5番茄亲环素L484_0087156.00×10-2768Cat r 1长春花亲环素L484_0220051.00×10-8571Sola l 5番茄亲环素L484_0023812.00×10-2461Asp f 27烟曲霉亲环素L484_0174751.00×10-7266Asp f 27烟曲霉亲环素L484_0239734.00×10-6660Sola l 5番茄亲环素L484_0275742.00×10-7162Cat r 1长春花亲环素Pkinase(PF00069)/ Usp(PF00582)/ Pkinase_Tyr (PF07714):蛋白激酶结构域,络氨酸激酶L484_0082132.00×10-15064Sal k 2猪毛菜蛋白激酶L484_0198637.00×10-14561Sal k 2猪毛菜蛋白激酶L484_0138435.00×10-13960Sal k 2猪毛菜蛋白激酶L484_0134664.00×10-15765Sal k 2猪毛菜蛋白激酶L484_0030017.00×10-15664Sal k 2猪毛菜蛋白激酶TIM(PF00121):磷酸丙糖异构酶L484_0249516.00×10-11161Tri a 31普通小麦磷酸丙糖异构酶L484_0252393.00×10-12474Tri a 31普通小麦磷酸丙糖异构酶L484_0107433.00×10-14978Tri a 31普通小麦磷酸丙糖异构酶Pectinesterase(PF01095)/ PMEI(PF04043):植物转化酶/ 果胶甲基酯酶抑制剂L484_0255947.00×10-13666Act d 7美味猕猴桃果胶酯酶L484_0104671.00×10-14960Sal k 1猪毛菜果胶酯酶L484_0007993.00×10-14265Act d 7美味猕猴桃果胶酯酶L484_0111205.00×10-13175Act d 7美味猕猴桃果胶酯酶L484_0111155.00×10-13762Act d 7美味猕猴桃果胶酯酶L484_0111182.00×10-13160Act d 7美味猕猴桃果胶酯酶L484_0111191.00×10-13563Act d 7美味猕猴桃果胶酯酶其他Pfam家族L484_0090416.00×10-6151Alt a 7链格孢YCP4 蛋白L484_0251346.00×10-12274Tri a 32普通小麦1-Cys过氧化还原酶L484_0139581.00×10-10667Cand a 3白念珠菌1-Cys过氧化还原酶L484_0000528.00×10-5095Sal k 3猪毛菜不依赖于维生素B12的甲硫氨酸合成酶L484_0000341.00×10-3691Sal k 3猪毛菜不依赖于维生素B12的甲硫氨酸合成酶L484_009365089Sal k 3猪毛菜不依赖于维生素B12的甲硫氨酸合成酶L484_003436076Sal k 3猪毛菜不依赖于维生素B12的甲硫氨酸合成酶L484_019026065Sola l 2番茄酸性β-呋喃果糖苷酶L484_0138872.00×10-10960Mus a 2小果野蕉Ⅰ类几丁质酶L484_0042735.00×10-3661Act d 4美味猕猴桃巯基蛋白酶抑制剂L484_023933073Tri a 34普通小麦甘油醛-3-磷酸脱氢酶L484_006727087Tri a 34普通小麦甘油醛-3-磷酸脱氢酶L484_006912087Tri a 34普通小麦甘油醛-3-磷酸脱氢酶L484_0176563.00×10-15767Tri a 34普通小麦甘油醛-3-磷酸脱氢酶L484_0095762.00×10-2363Tri a 41普通小麦NFKB 1线粒体泛素连接酶活化子L484_0170681.00×10-7680Cor a 13欧洲榛油质蛋白L484_0205536.00×10-6563Cor a 12欧洲榛油质蛋白L484_0039562.00×10-14461Hev b 7橡胶树马铃薯块茎特异蛋白L484_0039582.00×10-15461Hev b 7橡胶树马铃薯块茎特异蛋白L484_0132392.00×10-15261Hev b 7橡胶树马铃薯块茎特异蛋白L484_0255024.00×10-4460Cur l 3新月弯孢菌细胞色素CL484_0106767.00×10-12578Pis v 4阿月浑子超氧化物歧化酶L484_0112252.00×10-13186Hev b 10橡胶树超氧化物歧化酶L484_003380064Sola l 2番茄β-呋喃果糖苷酶L484_0081962.00×10-15570Pyr c 5西洋梨异黄酮还原酶相关蛋白L484_009016086Bet v 6疣皮桦苯基香豆素苄基醚还原酶
续表1
基因编号E值相似度(%)致敏原物种蛋白注释L484_0102551.00×10-3281Pru p 7巴旦木赤霉素调节蛋白L484_0114262.00×10-5283Tri a 45普通小麦延伸因子ⅠL484_0115004.00×10-2873Pru p 7巴旦木赤霉素调节蛋白L484_0131725.00×10-5673Hum j 1葎草未知蛋白L484_0185642.00×10-18080Pyr c 5西洋梨异黄酮还原酶相关蛋白L484_0185654.00×10-18079Pyr c 5西洋梨异黄酮还原酶相关蛋白
Bet v 1成员属于PR10家族,在植物的花粉、种子和果实中高表达,在多种植物的花粉和果实中具有过敏原活性[10]。有研究发现,桑葚中的1个17 ku蛋白疑似PR10家族的Bet v 1,是主要的过敏原[3]。Bet v 1的二级结构元件构成1个大的疏水口袋,具有储存或载体蛋白的功能[11],能够结合黄酮类化合物[12]。在本研究中,4个已被初步鉴定为致敏原的成员属于该家族,具有引起患者致敏反应的潜力。L484_001696的3-D结构模型具有典型的Bet v 1三级结构,进一步说明该家族桑树潜致敏原具有致敏潜力,值得深入研究。
TLP属于植物病程相关蛋白第5家族,能被多种胁迫诱导表达,在多种水果和作物中具有过敏原活性[8,13-14]。TLP蛋白3D结构中的功能域Ⅱ表现出非常保守的构象,预测这些氨基酸序列为主要的IgE结合表位[15]。L484_022590蛋白具有保守结构域THN,其三级结构由3个功能域组成:Domain Ⅰ为N端核心功能域,由2个反向平行的β片层组成;Domain Ⅱ由半胱氨酸二硫键形成的3个较短的α螺旋构成;Domain Ⅲ由2个β折叠和1个大环构成[15]。桑树中有4个蛋白与致敏蛋白Pru p 2具有60%以上的序列一致性。预测的L484_022590三维结构(图3)与典型TLP非常相似,其具体致敏活性值得研究。
RlpA-like_DPBB家族包含保守的DPBB(double-psi beta-barrel)折叠,DPBB功能域的N端常常连接花粉致敏原[16]。Kiwellin是猕猴桃中重要的富含半胱氨酸的RlpA-like_DPBB家族致敏原蛋白,来自绿色猕猴桃的Act d 5蛋白大约为22 ku,由N端kissper功能域和C端KiTH功能域组成,其中后者又由具有2个扩展环的6个β片层构成的筒体构成[17]。本研究中搜索到5个桑树潜在致敏原与猕猴桃Act d 5具有高度一致性。L484_007509的三维结构具有典型的kissper和KiTH功能域,具有致敏潜力。
几丁质结合蛋白家族属于碳水化合物结合结构域家族18(carbohydrate-binding module family 18,简称CBM18),具有几丁质识别或结合活性,存在于多种植物与真菌中,能结合N-乙酰葡萄糖胺[18]。该家族成熟蛋白N端的Chitin bind 1(PF00187)功能域能识别或结合几丁质亚基。本研究中发现4个该家族桑树潜在致敏原蛋白,与小果野蕉Mus a 2或巴西橡胶Hev b 6序列一致性均达70%以上,并构建了L484_024568的三维结构(图3)。
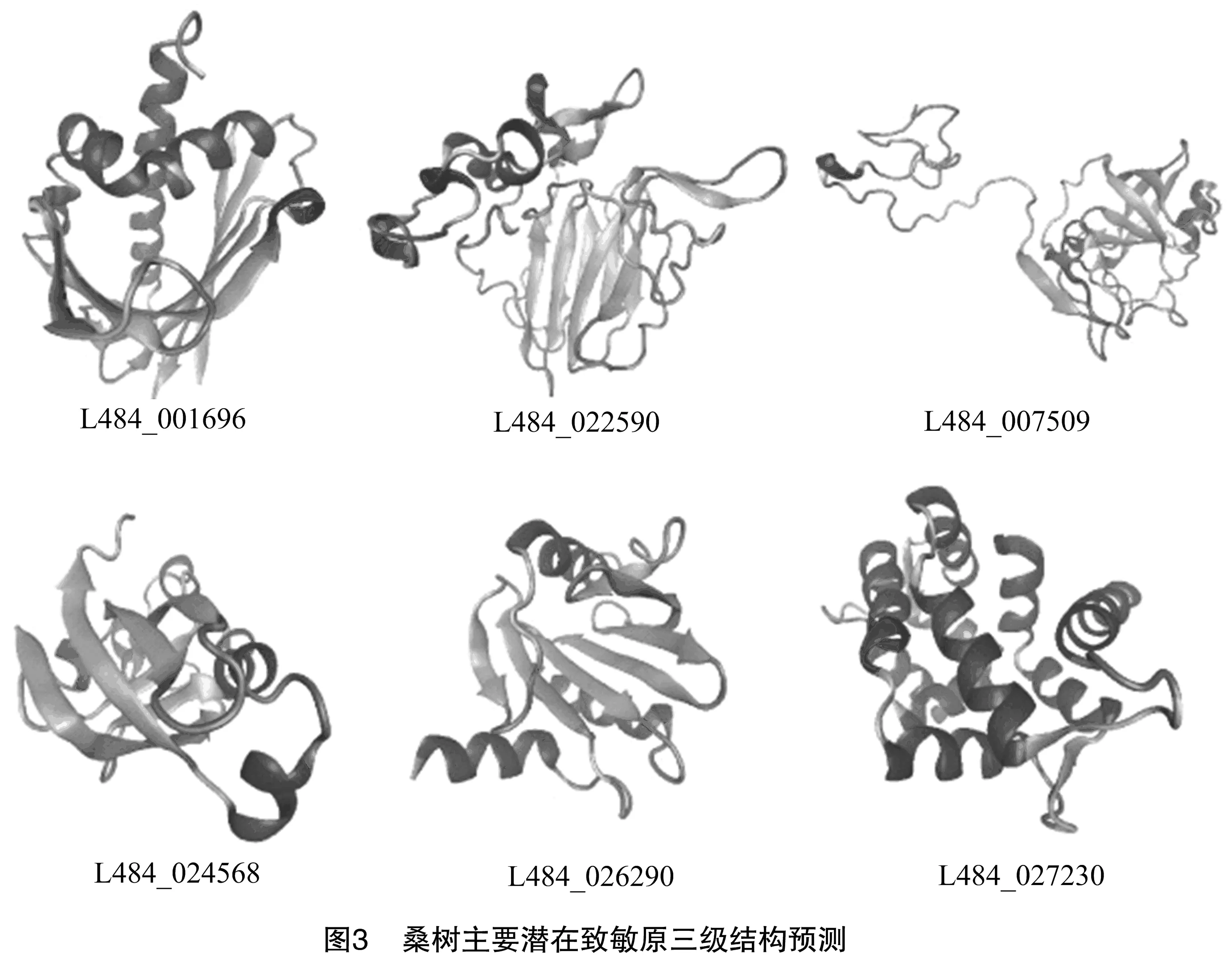
EF-hand家族属于钙结合蛋白,EF-hand基序包含螺旋-环-螺旋拓扑结构,能在环上结合钙离子。源自藜、狗牙根、油橄榄和猫尾草等植物花粉中的EF-hand家族成员具有致敏原活性。本研究发现,2个桑树潜在过敏原属于该家族,构建了L484_027230蛋白三维结构(图3),这些成员具有典型EF-hand家族蛋白的类似结构,很可能在桑树的花粉中具有致敏原活性。
3 讨论与结论
过敏性疾病是临床上的常见病症,因此,过敏反应性疾病的诊断和治疗的关键是分离和鉴定出引发过敏反应的致敏原。传统鉴定和获得致敏蛋白的方法需要经过表达、纯化、结晶、磁共振或X射线衍射等复杂的试验步骤,在实际操作中会遇到诸多问题。目前,随着基因组和蛋白质组数据的增加,以及生物信息学的发展,出现了多种比对和结构建模的软件,这些预测结果大大缩减了科研工作量,使研究工作更有针对性。
植物花粉和果实过敏的发病率都很高,目前在很多植物中已经鉴定出多种植物源致敏原。桑树花粉和果实均能引起人的过敏反应[2-3],而对桑树致敏原的研究较少。全面了解桑树内源性致敏原,包括致敏原的种类和数量、致敏原的时空表达特征等,对于桑树过敏症和交叉反应的诊断具有重要的参考价值。本研究以已知致敏原序列为探针,搜索桑树蛋白质数据库,共获得143条潜在致敏原序列。GO注释显示,它们属于细胞部分,参与细胞过程,说明它们主要为细胞建成成分,参与了重要的细胞生物学功能,具有结合活性,参与代谢过程和应激反应,说明这些蛋白能与其他组分结合,可能具有抗原抗体结合活性,是这些蛋白具有致敏性的重要前提。
桑树致敏原蛋白结构多样,本研究共查询到77个pfam保守结构域,主要包括HSP70、Profilin、Tubulin、Tubulin_C、Bet_v_1、Chloroa_b-bind和RlpA-like_DPBB结构域等。其中Bet v 1家族、Profilin家族、Thaumatin家族、kiwellin家族是较常见的致敏原类型,在多种植物的花粉和果实中具有过敏原活性[10,13-14,17],有些蛋白具有抗真菌活性。本研究中发现了多个桑树潜在致敏原成员属于这些家族,其蛋白与已鉴定的致敏原蛋白具有较高的序列一致性,其三级结构与已鉴定的致敏原蛋白结构类似,其致敏潜力值得深入研究。本研究也发现,部分桑树潜在致敏原蛋白属于糖基水解酶、糖苷水解酶和果胶酯酶家族,这些酶类参与了重要的细胞进程,关于其致敏原活性的报道较少,主要在植物的生长发育过程中发挥作用。
参考文献:
[1]Cianferoni A,Spergel J M.Food allergy:review,classification and diagnosis[J]. Allergology International,2009,58(4):457-466.
[2]Navarro A M,Orta J C,Sanchez M C,et al.Primary sensitization toMorusalba[J]. Allergy,1997,52(11):1144-1145.
[3]Choi J H,Sim J K,Oh J Y,et al.An IgE-mediated allergic reaction caused by mulberry fruit[J]. Allergy Asthma & Immunology Research,2015,7(2):195-198.
[4]Nair S,Kukreja N,Singh B P,et al.Identification of B cell epitopes of alcohol dehydrogenase allergen ofCurvularialunata[J]. PLoS One,2011,6(5):e20020.
[5]Hileman R E,Silvanovich A,Goodman R E,et al.Bioinformatic methods for allergenicity assessment using a comprehensive allergen database[J]. International Archives of Allergy and Immunology,2002,128(4):280-291.
[6]Bordoli L,Kiefer F,Arnold K,et al.Protein structure homology modeling using SWISS-MODEL workspace[J]. Nature Protocols,2008,4(1):1-13.
[7]Morano K A.New tricks for an old dog[J]. Annals of the New York Academy of Sciences,2007,1113(1):1-14.
[8]Breiteneder H.Thaumatin-like proteins-a new family of pollen and fruit allergens[J]. Allergy,2004,59(5):479-481.
[9]Faeste C K,Jonscher K R,Sit L,et al.Differentiating cross-reacting allergens in the immunological analysis of celery (Apiumgraveolens) by mass spectrometry[J]. Journal of AOAC International,2010,93(2):451-461.
[10]Soeria-Atmadja D,Önell A,Kober A,et al.Multivariate statistical analysis of large-scale IgE antibody measurements reveals allergen extract relationships in sensitized individuals[J]. Journal of Allergy and Clinical Immunology,2007,120(6):1433-1440.
[11]Fernandes H,Michalska K,Sikorski M,et al.Structural and functional aspects of PR-10 proteins[J]. The FEBS Journal,2013,280(5):1169-1199.
[12]von Loetzen C S,Jacob T,Hartl-Spiegelhauer O,et al.Ligand recognition of the major birch pollen allergen Bet v 1 is isoform dependent[J]. PLoS One,2015,10(6):e0128677.
[13]Esteve C,D’Amato A,Marina M L,et al.In‐depth proteomic analysis of banana (Musaspp.) fruit with combinatorial peptide ligand libraries[J]. Electrophoresis,2013,34(2):207-214.
[14]Hegde V L,Ashok K H G,Sreenath K,et al.Identification and characterization of a basic thaumatin-like protein (TLP 2)as an allergen in sapodilla plum(Manilkarazapota)[J]. Molecular Nutrition & Food Research,2014,58(4):894-902.
[15]Leone P,Menu-Bouaouiche L,Peumans W J,et al.Resolution of the structure of the allergenic and antifungal banana fruit thaumatin-like protein at 1.7-A[J]. Biochimie,2006,88(1):45-52.
[16]Jorgenson M A,Chen Y,Yahashiri A,et al.The bacterial septal ring protein RlpA is a lytic transglycosylase that contributes to rod shape and daughter cell separation inPseudomonasaeruginosa[J]. Molecular Microbiology,2014,93(1):113-128.
[17]Offermann L R,Giangrieco I,Perdue M L,et al.Elusive structural,functional,and immunological features of Act d 5,the green kiwifruit kiwellin[J]. Journal of Agricultural and Food Chemistry,2015,63(29):6567-6576.
[18]Wright H T,Sandrasegaram G,Wright C S.Evolution of a family ofN-acetylglucosamine binding proteins containing the disulfide-rich domain of wheat germ agglutinin[J]. Journal of Molecular Evolution,1991,33(3):283-294.
[19]Valenta R,Duchene M,Ebner C,et al.Profilins constitute a novel family of functional plant pan-allergens[J]. The Journal of Experimental Medicine,1992,175(2):377-385.
猜你喜欢
音乐教育与创作(2022年1期)2022-04-26
当代水产(2021年3期)2021-07-20
今日农业(2020年16期)2020-12-14
中成药(2018年5期)2018-06-06
小学生作文(中高年级适用)(2018年3期)2018-04-18
中成药(2017年8期)2017-11-22
学生天地(2016年16期)2016-05-17
淮海医药(2015年2期)2016-01-12
现代检验医学杂志(2015年1期)2015-02-06
中国药业(2014年17期)2014-05-26
