
猪伪狂犬病病毒的检测及主要毒力基因的分子特征分析
2018-10-15 10:21何小莉梁海英曾智勇汤德元张爱琼
中国兽医杂志 2018年6期
何小莉,梁海英,曾智勇,汤德元,王 彬,黄 涛,张爱琼,咸 文
(贵州大学动物科学学院,贵州贵阳550025)
猪伪狂犬病是由猪伪狂犬病病毒引起的一种高度接触性传染病,主要以新生仔猪的急性死亡以及4周龄以上仔猪出现神经症状,妊娠母猪繁殖障碍、流产、死胎及呼吸道症状为特征[1]。自1902年匈牙利首次发现该病以来已广泛分布于世界各地,1974年传入我国,至今全国已有 30多个省份发生流行,是严重危害我国养猪业的重要疫病之一[2]。伪狂犬病病毒(Pseudorabiesviurs,PRV)属疱疹病毒科α-疱疹病毒亚科,水痘病毒属,双链DNA病毒,全长约150 kb,基因组GC含量最高达73%,基因组编码约100种蛋白质,成熟的病毒粒子约含有50种蛋白质[3]。gE和TK基因是PRV主要毒力基因,但不是病毒增殖所必需的。gE基因在促进感染细胞向邻近非感染细胞融合,介导病毒在细胞间扩散起作用,TK基因所编码的胸苷激酶与病毒在神经组织中增殖有关[4]。自2011年以来,由于变异毒株的出现导致猪伪狂犬病发生率逐渐上升,也给本省养猪业造成了一定的经济损失。本试验通过对贵州省某规模化猪场疑似猪伪狂犬病发病猪进行病理剖检病变观察及PCR检测,并对其主要毒力基因(gE、TK基因)进行克隆测序及遗传进化分析,旨在为猪伪狂犬病的检测及相关基因遗传变异状况提供参考。
1 材料与方法
1.1 病料来源 贵州省丹寨地区某规模化养殖场送检的疑似PRV感染后发病死亡猪1头。
1.2 试验主要材料 大肠杆菌TOP10感受态细胞,购自上海生工生物工程技术服务有限公司; pMD19-T载体、DL-2 000 Marker、DNA抽提试剂盒及LATaqDNA聚合酶,购自TaKaRa公司;胶回收试剂盒,购自OMEGA公司。
1.3 引物 根据在GenBank中已发表的猪伪狂犬病毒PRV Fa株(KM189913.1)gE和TK基因序列,利用Oligo 5.0设计2对引物并委托TaKaRa公司合成,引物序列分别为gE-F1:5′-GGGGTACCATGCGGCCCTTTCTGCT-3′;gE-R1:5′-CCGGAATTCTTAAGCGGGGCGGGACAT-3′;TK-F2:5′-TTTGAATTCGCGGCACGTCTTGAGCTCGA-3′;TK-R2:5′-TTTAAGCTTCGCCGACCAGGACGAACAGG-3′。
1.4 病猪剖检及病原检测 无菌剖检发病死猪,观察其组织病理病变,适量取其心、肝、脾、肺、肾、脑等组织混合研磨,反复冻融3次后,取上清,用DNA抽提试剂盒提取核酸,用PRVgE特异性引物进行PCR检测,反应结束后,分别取7 μL扩增产物于1%琼脂糖凝胶中电泳检测,并记录结果。
1.5 PRV GZDZ2016株gE和TK基因的克隆及序列分析 以组织核酸为模板进行PRVgE及TK基因的PCR扩增,分别将PCR产物纯化回收后连接于pMD19-T 载体,转入大肠杆菌TOP10感受态细胞,将重组质粒送深圳华大基因科技服务有限公司进行序列测定。用DNAStar软件对PRV GZDZ2016株的测序结果与参考毒株的gE、TK基因的序列分别进行基因分子特征分析及核苷酸、氨基酸同源性分析,并用MEGA7.0绘制遗传进化树。
2 结果
2.1 剖检病理变化及病原检测结果 剖检病死猪可见皮肤散布出血点(见中插彩版图1),脾脏表面有许多散在大小不一颗粒状的白色坏死点(见中插彩版图2),肝脏出现大面积瘀斑、且表面有白色坏死灶(见中插彩版图3),肺脏有瘀斑且间质增宽(见中插彩版图4)。将各组织器官经过处理后,提取核酸用PRVgE特异性引物进行检测,结果显示,病料中检出PRVgE特异性核酸阳性条带,且与预期大小相符。
2.2gE、TK全基因PCR扩增及鉴定gE、TK全基因PCR扩增结果显示扩增片段大小与预期条带相符,分别得到约1 800 bp和1 500 bp大小的目的条带(图5)。克隆质粒经双酶切和PCR鉴定后,并将阳性重组质粒送至上海生工生物工程技术服务有限公司进行测序。
2.3gE和TK基因序列分析
2.3.1gE基因分子特征分析gE基因的核苷酸序列分析结果显示,PRV GZDZ2016株的gE基因序列与参考毒株核苷酸序列同源性在97.6%~99.9%,其中与HN1201、HXN、JX-2012、GA株的同源性最高;PRV GZDZ2016株的gE基因序列与参考毒株氨基酸序列同源性在95.7%~99.8%,氨基酸序列分析结果表明,PRV GZDZ2016株gE基因所编码的氨基酸与PRV Fa标准株氨基酸序列相比,48位和496位均各有一个天冬氨酸(D)的插入,第236位由亮氨酸(L)变为脯氨酸(P)。
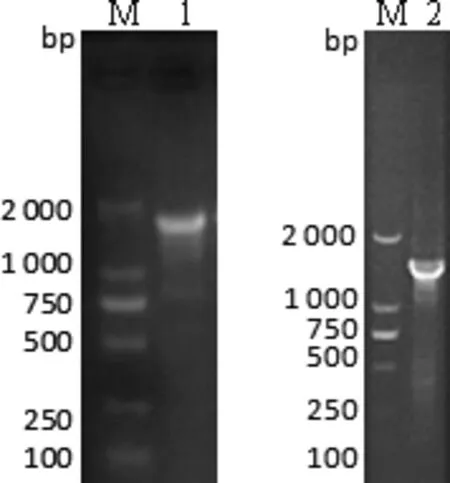
图5 PRV GZDZ 2016株 gE、TK全基因序列的PCR扩增
遗传进化树结果显示,gE基因的遗传树明显分为2个大群。其中PRV GZDZ2016株与文献报道的变异代表株处在于同一分支上,与国内早期的分离株共同构成一个亚群;国外的毒株形成另外一个大的分支,与国内的经典毒株相距较远(图6),进一步说明了该毒株具有变异毒株特征,且与国外流行毒株和国内经典毒株的亲缘关系较远。

图6 PRV GZDZ2016株gE基因遗传进化树
2.3.2TK基因分子特征分析TK基因的核苷酸序列分析结果显示,PRV GZDZ2016株TK基因与疫苗株Bartha株的核苷酸相似性和氨基酸同源性分别为99.4%、98.8%;与国外的Becker、Kaplan、NIA3等株之间的核苷酸相似性和氨基酸同源性分别为99.4%~99.6%和98.8%~99.4%;与国内近几年分离毒株JS-2012、HeN1、TJ等株之间的核苷酸相似性和氨基酸同源性分别为99.5%~99.8%和98.8%~99.4%;其中与HN1201、HeN1、JS-2012、TJ、HBCL-2012、BJ/YT株的同源性最高。氨基酸序列分析结果表明,TK基因所编码的氨基酸则第6位氨基酸由异亮氨酸(I)变为丙氨酸(A),第157位氨基酸由天冬氨酸(D)变为甘氨酸(G)。
遗传进化树结果显示,TK基因可分为4个基因群,本研究的PRV GZDZ2016株与国内分离的大多数毒株构成基因1群,国内部分毒株与国外分离株构成另外3个基因群(图7)。
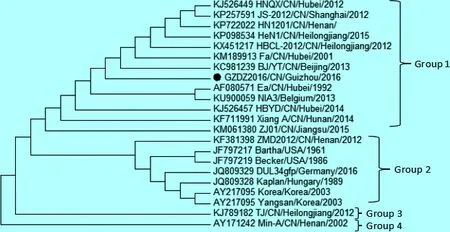
图7 PRV GZDZ2016株TK基因遗传进化树
3 讨论
目前,猪伪狂犬病疫情形势严峻,2012年以来在中国多个省份许多使用PRV基因缺失活疫苗进行免疫的规模化猪场出现了猪狂犬病的流行,并从已免疫过疫苗的猪群分离到PRV变异毒株。gE和TK基因是PRV主要毒力基因,gE基因在促进感染细胞向邻近非感染细胞融合,介导病毒在细胞间扩散起作用[5];TK基因失活,则PRV对宿主的毒力丧失或显著下降[6]。因此,对贵州省PRV临床流行毒株的gE、TK基因的克隆分析,将有利于掌握贵州省PRV流行株的遗传变异情况。
本研究通过病理剖检、PCR鉴定及序列分析等方法,证实送检发病猪由于感染PRV致死,并将该感染毒株命名为PRV GZDZ2016株。对该毒株gE全序列分析结果显示,gE基因氨基酸的第48位和第496位各存在1个天冬氨酸(D)的插入,这两个位点的变化与此前报道的HN1201[7]、TJ[8]等变异毒株具有相同的变异模式,表明 PRV GZDZ2016株为PRV变异毒株,同时第236位点出现L→P的突变,这个氨基酸位点位于gE抗原表位E区,该突变是否会对PRV的毒力或gE抗原性发生漂移尚需进一步研究。对该毒株TK基因的序列分析结果表明,最具特征性的是PRV GZDZ2016株TK基因核苷酸序列第17位和470位出现了独特性的碱基突变,第17位由T→C,第470位由 A→G,并造成第6位氨基酸由 I→A 和第157位氨基酸由D→G。Darb等研究认为5-18aa区间为ATP结合位点,该区域对TK蛋白的催化功能十分重要[9],上述PRV GZDZ2016株TK基因氨基酸序列第6 位氨基酸由I→A,因此,TK基因在该位点的突变可能会对 PRV 的毒力产生一定的影响。
基于PRV GZDZ2016株gE、TK基因系统进化树分析结果显示,PRV GZDZ2016株与2012年以来国内分离的PRV变异毒株及国内经典毒株的亲缘关系较近,而与国外毒株的亲缘关系相对较远,综上所述,本研究的PRV GZDZ2016株具有一定的当前PRV流行株的代表性,属于PRV变异毒株。PRV GZDZ2016株的分子特征将对变异株的分子流行病学调查提供重要的参考,以更好地对猪伪狂犬病的诊断和防控提供科学依据。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
农药科学与管理(2019年9期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
中国抗生素杂志(2019年6期)2019-07-06
中国饲料(2019年19期)2019-03-25
中国人兽共患病学报(2017年11期)2017-12-13
