
低温诱导下2种烤烟品种苗期叶片保护酶活性及烤后化学成分的变化
2018-12-07 06:09王欣亚吴委林王家川金大翔朴世领
延边大学农学学报 2018年3期
王欣亚, 吴委林, 万 悦, 王家川, 金大翔, 朴世领
(延边大学农学院,吉林 延吉133002)
烟草早花是指在某些特定条件下,烟株未达到正常栽培条件下应有的株高和叶片数时便提前开花的现象[1-2]。影响烟草早花的因素有品种差异,温度(低温),日照长度(短日照)等,低温是引起早花的主要原因之一[2]。早花现象的发生对烟草产业的发展影响严重[3],这是一种严重影响烟叶产量的异常现象,是生长与发育异常化的结果[4]。烟苗在移栽前受到低温寒潮侵袭时易出现早花,而且随着移栽过早或烟苗增大,出现早花少叶的程度更大[5]。延边地区气候特点为春季气温低,雨水少,无霜期短,隔年烟区便发生早花现象[3,6]。适宜烤烟生长的温度范围较大,且烟草是喜温不耐寒的作物[7],寒冷胁迫条件下的生长发育会受到抑制,从而影响其品质和产量[8-10]。在寒冷胁迫中,可以调控植物体内活性氧代谢平衡的是SOD、CAT、POD这3种最主要的酶类[11-14]。对植物进行有效的低温诱导可以提高其抗寒能力,后期作为亲本在培育抗寒品种有广泛的应用[15-17]。本文利用人为控制烟草苗期短期生长温度,通过调查农艺性状、保护酶变化以及烤后化学成分,分析了烟草苗期低温持续时间对烟草生理生化指标的影响,对烟草耐寒品种筛选、挖掘抗早花基因提供参考。
1 材料与方法
1.1 材料
试材选用6片真叶吉烟9号和云烟87,试验地为地势平坦且具有灌溉条件的延吉市依兰镇龙渊村烟叶种植基地,土壤质地为壤土,土层深厚,肥力均匀且适中。
1.2 试验设计
试验采用随机区组设计,设不同品种(A1、A2),不同低温诱导持续时间(B1、B2)2个试验因素,分别为:A1:吉烟9号,A2:云烟87,B1:在人工气候箱进行12 ℃低温处理持续10 d,B2:在人工气候箱进行12 ℃低温处理持续4 d,CK1:未经低温诱导处理的吉烟9号,CK2:未经低温处理的云烟87。
将2盘(100株/盘)正常生长的6叶龄吉烟9号及云烟87分别放置于人工气候箱中并将低温诱导温度设为(12±0.5) ℃,人工气候箱内相对湿度为75%,光暗时长分别为0.5 d,气候箱内光照度>10 000 lx,2个处理分别标记A1B1、A2B1。待A1B1和A2B1处理至第6天后,将另2盘(100株/盘)正常生长至6叶龄的吉烟9号及云烟87放置于另一个人工气候箱内,按同样设置方法对其进行处理,分别为A1B2、A2B2。CK1、CK2正常放置温室管理。当第1组处理达10 d,第2组处理达4 d后将进行低温处理的烟苗全部取出,置于温室同对照组共同进行缓苗2 d。在天气晴朗的5月25日统一移栽至大田。6个处理分别为A1B1、A1B2、CK1、A2B1、A2B2和CK2,每个处理30株,试验采取随机排列,共设18个小区,进行3次重复。
1.3 测定项目及测定方法
1.3.1 苗期保护酶活性测定
在低温诱导0、2、4、6、8、10 d和恢复后2 d时测定苗期叶片相关保护酶活性,每次在下午3点左右取样,选取大小基本一致的中部烟叶,采集样品后迅速放入密封袋存于保温冰盒取回,置于-85 ℃超低温冰箱(MDF-UT3V型)保存。采用愈创木酚法[18]测定POD的活性;采用氮蓝四唑法[19]测定SOD的活性;采用紫外分光光度法[18]测定CAT的活性。
1.3.2 移栽前总干物质积累、根冠比的调查
总干物质积累及根冠比的调查是在烟株移栽前1 d。在每个小区选取3棵长势基本一致的烟株,洗清根部并测量鲜重后,在精密鼓风干燥箱(BAO-250A)进行105 ℃杀青15 min,85 ℃恒温烘干48 h后,测量干重。用手术刀片在根茎处切开,分别计算地上部干重及地下部干重,并计算根冠比及总干物质积累。
1.3.3 田间烟株株高及有效叶数的调查
调查并记录每株的株高及有效叶数,有效叶数调查是在移栽后的79 d,通常叶片的长度大于20 cm方可记作有效叶[20]。
1.3.4 现蕾数及现蕾率的调查
在移栽后的52、53、54、56 d,分别调查每个小区现蕾数及现蕾率。以肉眼能看到花蕾为现蕾标准,每个小区现蕾的烟株数达到50%视为现蕾期[21]。
1.3.5 烤后化学成分的分析
原烟按国家分级标准(GB2635-92)进行分级,6个处理取C3F各1 kg ,化学成分的测定均委托吉林烟草工业有限责任公司进行测定。
1.4 数据统计和分析
试验采用软件SPSS 19.0进行数据统计和分析,用Duncan方法对试验数据进行单因素方差分析(One-way ANOVA)。
2 结果与分析
2.1 不同烤烟品种在苗期低温诱导下相关酶活性变化
2.1.1 不同烤烟品种在苗期低温诱导下SOD活性变化
低温持续处理10 d不同组合SOD活性变化结果(图1)说明:随着低温持续时间的增加,低温处理(A1B1、A2B1)的SOD活性均呈先升后降的趋势,在2 d时达到峰值,而未处理的对照(CK1、CK2)均呈平稳上升趋势,且低温处理的SOD活性均显著高于对照(P<0.05)。在缓苗恢复2 d后,低温处理的SOD活性趋于CK水平,且无显著性差异(P>0.05)。低温处理2~10 d期间,SOD活性云烟87显著小于吉烟9号(P<0.05)。与对照相比,低温处理(A1B1、A2B1)的增幅,在2 d时最高,分别为(115.17±7.55)%和(85.15±5.44)%,且两者增幅之间存在极显著差异(P<0.01)。
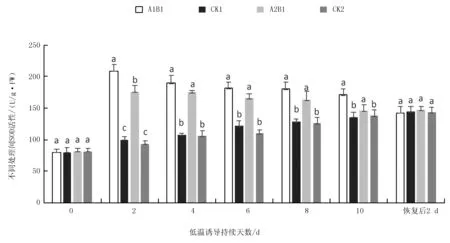
注:不同小写字母代表在处理间显著差异水平(P<0.05),下同。
低温处理结束后,不同处理间SOD活性分析结果(图2)表明:当低温处理4 d和10 d后,不同处理间均有显著差异(P<0.05),SOD活性从小到大依次为CK1,CK2,A2B1,A1B1,A2B2和A1B2。各处理增幅由大到小依次是A1B2,A2B2,A1B1和A2B1,其值分别为90.53%,58.72%,26.30%,6.22%,且增幅间均存在极显著差异(P<0.01)。

图2 低温处理4 d及10 d后不同处理的SOD活性
2.1.2 不同烤烟品种在苗期低温诱导下POD活性变化
低温持续处理10 d不同处理POD活性变化结果(图3)说明:随低温持续时间增加,在0~2 d,低温处理(A1B1、A2B1)的POD活性均上升明显,2~4 d平缓上升,在第4天时POD活性达到最高值,随后则不断下降,而对照组均呈平稳上升趋势,且低温处理的POD活性均显著大于对照(P<0.05)。在低温处理结束恢复2 d后,处理组POD活性降至CK水平,处理间均无显著差异(P>0.05)。云烟87的POD活性小于吉烟9号,但无显著性差异(P>0.05)。与对照相比,低温处理(A1B1、A2B1)的POD活性增幅吉烟9号大于云烟87,增幅在第4天时最明显,其值分别为(97.71±5.51)%和(90.51±4.55)%,其增幅间存在极显著差异(P<0.01)。

图3 低温处理持续10 d下不同烤烟品种POD活性变化
低温处理结束后,不同处理间POD活性分析结果(图4)表明:低温诱导4 d及10 d结束后,低温持续4 d的处理(A1B2、A2B2)与10 d的处理(A1B1、A2B1)及0 d的处理(CK1、CK2)有显著差异(P<0.05),而各处理间POD活性由小到大依次为CK2,CK1,A2B1,A1B1,A2B2和A1B2。当低温诱导10 d结束后,各处理增幅由大到小的顺序依次是A1B2(97.03%),A2B2(93.13%),A2B1(19.97%)和A1B1(15.21%),且低温持续10 d与4 d的处理增幅间均存在极显著差异(P<0.01)。

图4 低温处理4 d及10 d后不同处理的POD活性
2.1.3 不同烤烟品种在苗期低温诱导下CAT活性变化
低温持续处理10 d不同组合的CAT活性变化结果(图5)说明:随低温处理持续时间的增加,低温处理(A1B1、A2B1)在0~2 d均为明显上升趋势,至2 d时的CAT活性达峰值,在2~10 d趋于平缓趋势,变化不明显,而未低温处理的对照(CK1、CK2)均平缓上升,且低温处理CAT活性均显著高于对照(P<0.05)。在室温缓苗恢复2 d后,处理的A1B1、A2B1与CK1、CK2的CAT活性降至同一水平,没有显著性差异(P>0.05)。与对照相比,低温处理(A1B1、A2B1)的CAT活性增幅,吉烟9号亦大于云烟87,在2 d时增幅最大,其值分别为(115.19±4.55)%及(92.27±6.51)%,且增幅间存在极显著差异(P<0.01)。
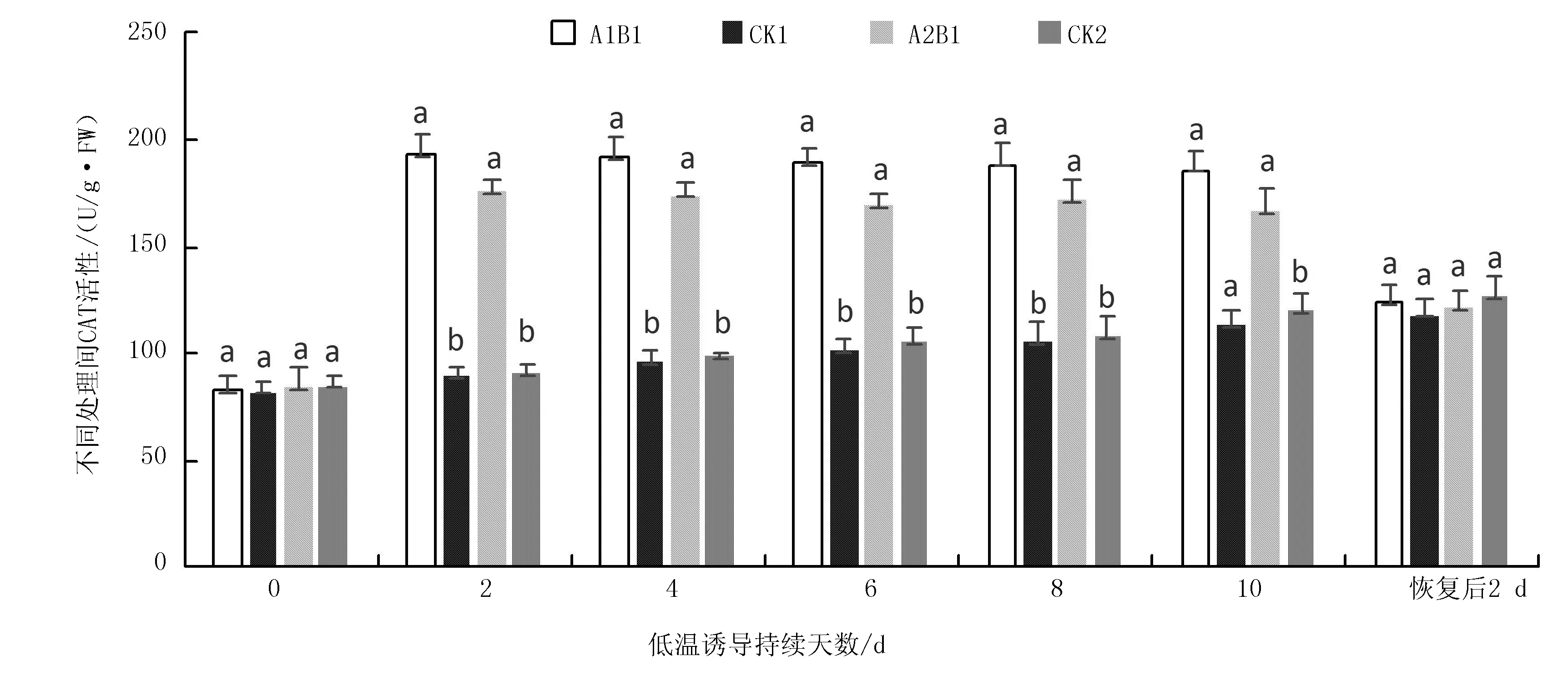
图5 低温处理持续10 d下不同烤烟品种CAT活性变化
低温处理4 d及10 d后不同处理的CAT活性(图6)均与对照(CK1、CK2)有显著差异(P<0.05),各处理间自小到大依次是CK1,CK2,A2B1,A2B2,A1B1和A1B2。各处理增幅由大到小依次是A1B2,A1B1,A2B2和A2B1,其值分别为69.54%,64.24%,57.76%,39.39%,低温持续4 d、10 d的吉烟9号(A1B2、A1B1),与低温持续4 d的云烟87(A2B2)三者增幅间无极显著差异(P>0.01),与低温持续10 d的云烟87(A2B1)增幅间均存在极显著差异(P<0.01),低温持续4 d的吉烟9号增幅最大。

图6 低温处理4 d及10 d后不同处理的CAT活性
2.2 低温诱导对移栽前总干物质的影响
移栽前1 d不同处理移栽前总干物质积累见表1。各处理间的总干物质均无显著差异(P>0.05)。根冠比上,从大到小依次为CK2,CK1,A1B2,A2B2,A1B1和A2B1。CK2与A1B1、A2B1有显著差异(P<0.05),而与其他处理(A1B2、CK1、A2B2)均无显著差异(P>0.05)。吉烟9号的根冠比上,由大到小的顺序依次为CK1,A1B2,A1B1,但其三者均无显著差异(P>0.05);云烟87的根冠比上,由大到小依次为CK2,A2B2,A2B1,且CK2与A2B1有差异性显著(P<0.05),但与A2B2差异不显著(P>0.05);不同低温持续处理时间上,CK最大,其次是4 d,最后是10 d。由此可见,随着低温持续时间的增加,总干重和根冠比均减小。
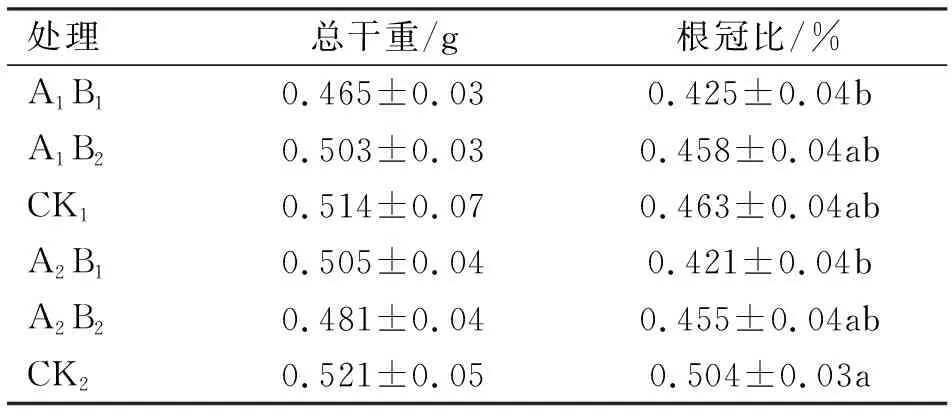
表1 移栽前不同处理的总干重及根冠比
2.3 不同处理间株高及有效叶数的调查
移栽后79 d株高(表2)从大到小依次是CK1,CK2,A1B2,A2B2,A1B1和A2B1。吉烟9号的株高由大到小依次是CK1,A1B2,A1B1,且CK1与A1B2、A1B1有显著性差异(P<0.05);云烟87的株高最高的为CK2,其次是A2B2,最后是A2B1,且CK2与A2B1、A2B2均存在显著差异(P<0.05)。移栽后79 d有效叶片数最多的为CK1,其次是A1B1与A2B2,A2B1最少。吉烟9号中,A1B2与CK1有效叶片数均大于18片,CK1与A1B2、A1B1均有显著差异(P<0.05);云烟87中,有效叶片数仅CK2大于18叶,且与A2B2、A2B1有显著性差异(P<0.05);相同低温持续时间,吉烟9号的有效叶数均大于云烟87,但差异不显著(P>0.05)。由此可见,低温持续10 d的云烟87(A2B1)和吉烟9号(A1B1)及低温持续4 d的云烟87(A2B2)极有可能已发生早花现象,低温持续4 d的吉烟9号(A1B2)和CK1、CK2均无早花迹象。

表2 移栽后79 d各处理的株高及有效叶数
2.4 烟株现蕾数、现蕾率的调查
移栽后52~56 d的现蕾数和现蕾率见表3。

表3 不同低温持续时间的现蕾数及现蕾率
由表3可知,各处理间均无显著差异(P>0.05),移栽后53 d的现蕾数和现蕾率,从小到大依次为CK1,CK2,A2B2,A1B2,A1B1,A2B1。其中,现蕾数最多的为A2B1,CK2次之,CK1最少。低温持续10 d的处理现蕾数及现蕾率最多,其次为4 d,0 d(CK)最少。大田烟株基本全部现蕾是在移栽后66 d,A2B1现蕾期是在移栽后的52 d,A1B1和A1B2的现蕾期是移栽后的54 d,而A2B2、CK1和CK23者的现蕾期均在移栽后56 d。由此可知,最早达现蕾期的为低温持续10 d的云87(A2B1),比对照(CK1、CK2)现蕾进程提前4 d左右,且比低温处理的吉烟9号(A1B1、A1B2)现蕾进程提前2 d左右。
2.5 烤后化学成分的分析
由表4可知,各处理总糖、还原糖含量总体均偏高,CK1在总糖和还原糖含量方面更接近适宜范围(总糖的适宜范围18%~22%,还原糖的适宜范围14%~18%),略低于其他处理,CK2的烟碱最高,A1B1最低,CK2的烟碱含量更接近适宜范围(烟碱的适宜范围1.5%~3.5%),烟碱含量高低很大程度上决定了烟叶的内在品质、安全性及可用性。施木克值也是衡量烟叶质量的重要指标,A1B2,CK1,CK2的施木克值与其他处理相比更低一些,但更接近适宜范围(施木克值的适宜范围在2左右)。总的来看,化学成分协调性更好的是对照(CK1,CK2)及低温持续4 d的吉烟9号(A1B2)。

表4 不同处理的化学成分含量
3 讨论与结论
3.1 讨论
植物体在经受低温胁迫时分泌大量氧自由基,超氧化物歧化酶(SOD)通过催化 O-2的歧化反应清除O-2,同时产生H2O2及·OH自由基,过氧化物酶(POD)与它们的清除密切相关,过氧化氢酶(CAT)是将H2O2还原为H2O和O-2,加强抗氧化作用,提高抗逆性从而起到保护作用[22]。
低温胁迫能显著降低保护酶活性(SOD、POD、CAT),而能保持较高的酶活性是耐寒性品种响应低温胁迫的重要生理指标。低温处理的苗期保护酶(SOD、POD、CAT)活性均显著高于CK呈先升高后降低的趋势,低温持续4 d时POD活性达到峰值,在低温持续2 d时SOD及CAT活性达到峰值,这与王家川[6]的研究结果相一致。低温持续时间上,4 d的处理(A1B2、A2B2)酶活性增幅均大于10 d的处理(A1B1、A2B1);品种上,吉烟9号苗期3种保护酶活性增幅均大于云烟87,而据前人研究表明,保护酶(SOD、POD、CAT)活性增幅越大,表明品种抗逆性强[23],因此,本研究结果说明,吉烟9号的耐寒性更强。据研究[21],成熟期的烤烟烟叶一般在18~22片,移栽后79 d有效叶数的调查表明,低温持续10 d吉烟9号、低温持续10 d云烟87以及低温持续4 d的云烟87有效叶数都未达到18片,同CK比少1~2片叶,表现为植株矮小,叶数锐减,叶片窄小,与刘建峰[24]的研究相吻合。本试验进行的大田现蕾数和现蕾率调查表明,低温持续10 d的处理(A1B1、A2B1)现蕾率较高,也存在着随低温持续时间的增加其现蕾期会逐渐提前的现象。招启柏等[25]与孙俊佳[26]的研究结果也证实了低温处理的烟株在移栽后与对照相比提前。本试验结果显示,无论是在现蕾率还是有效叶数上,低温持续时间越长,对烟草成花影响程度越大,尤其是低温持续10 d的处理(A1B1、A2B1)更为显著,与金磊等[27]研究基本一致。烟碱含量最高的是CK2,低温持续10 d的吉烟9号(A1B1)最低,结果对比后与前人研究结果基本一致[23]。烟碱上普遍低于适用范围,云烟87高于吉烟9号,且接近适宜范围;总糖及还原糖上,各处理均高于适用范围,与云烟87相比,吉烟9号更接近适宜范围;施木克值上,吉烟9号比云烟87更接近正常值;吉烟9号的钾氯比云烟87更接近适宜范围。总之,吉烟9号的化学成分协调性优于云烟87。
3.2 结论
吉烟9号苗期3种保护酶(SOD、POD、CAT)活性及增幅均大于云烟87;农艺性状(干重、根冠比、株高及有效叶数)上,吉烟9号均优于云烟87;现蕾数及现蕾率均随低温持续时间的增加而增加,且吉烟9号均低于云烟87。低温持续10 d的云烟87最早达到现蕾期,且与吉烟9号及对照相比提前2~4 d;化学成分分析上,吉烟9号比云烟87更协调。总之,两个品种在苗期12 ℃低温处理的持续时间越长,其早花迹象越明显,吉烟9号比云烟87的耐寒性更强。
猜你喜欢
今日农业(2021年9期)2021-11-26
今日农业(2021年9期)2021-11-26
今日农业(2020年23期)2020-12-15
农业技术与装备(2020年10期)2020-11-03
云南农业(2020年5期)2020-06-12
今日农业(2019年12期)2019-08-13
现代园艺(2018年3期)2018-02-10
汽车与安全(2017年2期)2017-03-14
诗歌月刊(2014年3期)2015-04-14
传奇故事(破茧成蝶)(2015年8期)2015-02-28
