
泥东风螺线粒体基因组及其遗传多样性分析
2019-01-17 02:05周先文王晓清巫旗生曾志南
水产科学 2019年1期
熊 钢,周先文,王晓清,巫旗生,康 骊,秦 溱,曾志南
( 1.湖南生物机电职业技术学院,湖南 长沙 410127; 2.水产高效健康生产湖南省协同创新中心,湖南 常德 415000; 3.湖南农业大学,湖南 长沙 410128; 4.福建省水产研究所,福建 厦门 361013 )
东风螺属(Babylonia)属软体动物门、腹足纲、新腹足目、蛾螺科[1],俗称花螺、南风螺和泥螺等,是我国沿海重要的经济软体动物。我国现有方斑东风螺(B.areolata)、泥东风螺(B.lutosa)和台湾东风螺(B.formosae)3种[2-3]。泥东风螺肉质鲜美,是畅销的优质海产贝类,在近十年开发为海水养殖品种[4-5]。然而泥东风螺野生资源锐减,为此泥东风螺人工繁育[6-7]、人工增殖[8]和增殖效果评估[9]相关研究工作相继开展。
泥东风螺作为后生动物,其线粒体DNA为典型的环状双链分子结构, 长度大多为14~18 kb,编码37个基因,包括13个蛋白质基因、22个tRNA以及12S RNA和16S RNA[10]。线粒体基因组是核外遗传物质,具有分子量小、结构简单、母系遗传等特点,成为生物多样性、群体遗传学与系统进化研究中的重要分子标记[11-12]。目前,线粒体基因已作为种类鉴定、系统发育分析和群体遗传多样性的重要分子标记,并且线粒体ND2、16S RNA、Cyt b基因、D-loop序列已被应用于贝类[13-14]、甲壳类[15]、鱼类[16-17]等水产动物物种遗传多样性及系统发育关系分析中。
为调查泥东风螺野生群体资源遗传多样性,笔者采用PCR扩增和直接测序技术对采自北海、连江和长乐的野生泥东风螺群体样本进行群体遗传多样性和遗传距离分析,从分子水平阐明我国沿海泥东风螺的遗传性状,研究结果将为泥东风螺资源恢复、保护和利用提供科学依据。
1 材料与方法
1.1 样品采集
泥东风螺为自福建连江(30个样本)、福建长乐(30个样本)和广西北海(30个样本)捕捞的野生泥东风螺群体。
1.2 总DNA的提取
取泥东风螺腹足,采用天泽基因柱式动物DNA提取试剂盒提取泥东风螺总DNA。总DNA用1%琼脂糖凝胶电泳检测,-20 ℃保存备用。
1.3 基因扩增和测序
以获得的泥东风螺转录组中线粒体基因信息设计扩增引物[18],扩增福建连江野生泥东风螺样本总DNA。PCR扩增条件为:95 ℃预变性3 min;94 ℃ 30 s,50 ℃ 30 s,72 ℃ 60 s,共33个循环;最后72 ℃延伸5 min。PCR产物经1.0%琼脂糖凝胶电泳、切胶和回收纯化后送铂尚生物进行双向测序。

表1 泥东风螺线粒体基因组扩增引物
1.4 线粒体基因组环状结构和序列分析
将送检测序结果用DNAMAN软件进行校对和人工去除错误拼接(GenBank登陆号:KF897830)。利用OGDraw软件[19]对泥东风螺线粒基因组进行制图,并在GenBank中进行Blast分析;采用Mega 5.1[20]软件构建最大似然法和邻接法物种线粒体基因组分子系统树,1000次重复抽样自展法评估各分支的置信度。用DnaSp 5.1[21]软件分析3个野生群体基因的遗传多样性参数,使用Mega 5.1[20]软件计算群体内和群体间的遗传距离。
2 结果与分析
2.1 线粒体基因组环状结构
OGDraw构建的泥东风螺线粒体基因组结构(图1)表明,泥东风螺线粒体基因组含有13个蛋白质编码基因、22个tRNA和2个rRNA;其中ATPase6、ND1、ND4L基因的终子密码子为TAG,ND5基因则为不完全终子密码子;其中ND2与COXⅠ,tRNA-Trp与tRNA-Gln,tRNA-Gly与tRNA-Glu,tRNA-Glu与12S RNA、tRNA-Glu,ND6与Cytb、ND4L与ND4基因存在重叠区;在环状线粒体基因结构中,tRNA-Met、tRNA-Tyr、tRNA-Cys、tRNA-Trp、tRNA-Gln、tRNA-Gly、tRNA-Glu和tRNA-Thr位于环形线粒体L链上。
2.2 基因相似性和保守性分析
在GenBank基因库中进行相似性分析得到17种腹足纲贝类动物线粒体基因组(表2)。使用DNAMAN软件分析泥东风螺与17种腹足纲动物线粒体基因组全长及线粒体上12S RNA、16S RNA、COXⅠ、COXⅡ、COXⅢ、和Cytb基因相似性(表3)。研究结果表明,18种腹足纲动物线粒体基因组间相似性为70.1%~92.9%;泥东风螺与其他贝类间的相似性为74.1%~92.9%,泥东风螺与方斑东风螺相似性最高,达92.9%;除与似鲍罗螺(Concholepasconcholepas)的COXⅢ基因相似性为50.4%外,泥东风螺线粒体上的COXⅠ、COXⅡ、COXⅢ、12S RNA、16S RNA、Cytb基因与其他17个物种对应基因的相似性为70.07%~99.00%;将泥东风螺与其他贝类线粒体基因组全长及各基因相似性作折线图(图2)可知,12S RNA和16S RNA相似性折线与线粒体基因组全长相似性折线变化相似。经12S RNA和16S RNA基因保守性分析发现,物种之间12S RNA的391~491 bp区域为高度变异区,物种之间16S RNA的1~106 bp和361~481 bp区域为高度变异区,而物种之间16S RNA的1191~1317 bp区域为高度保守区;物种之间COXⅢ相似性与线粒体基因全长相似性表现相当,而COXⅠ、COXⅡ基因相似性更高,这两个基因的物种间相似性均值分别为82.96%和83.00%。

图1 泥东风螺线粒体基因组环状结构
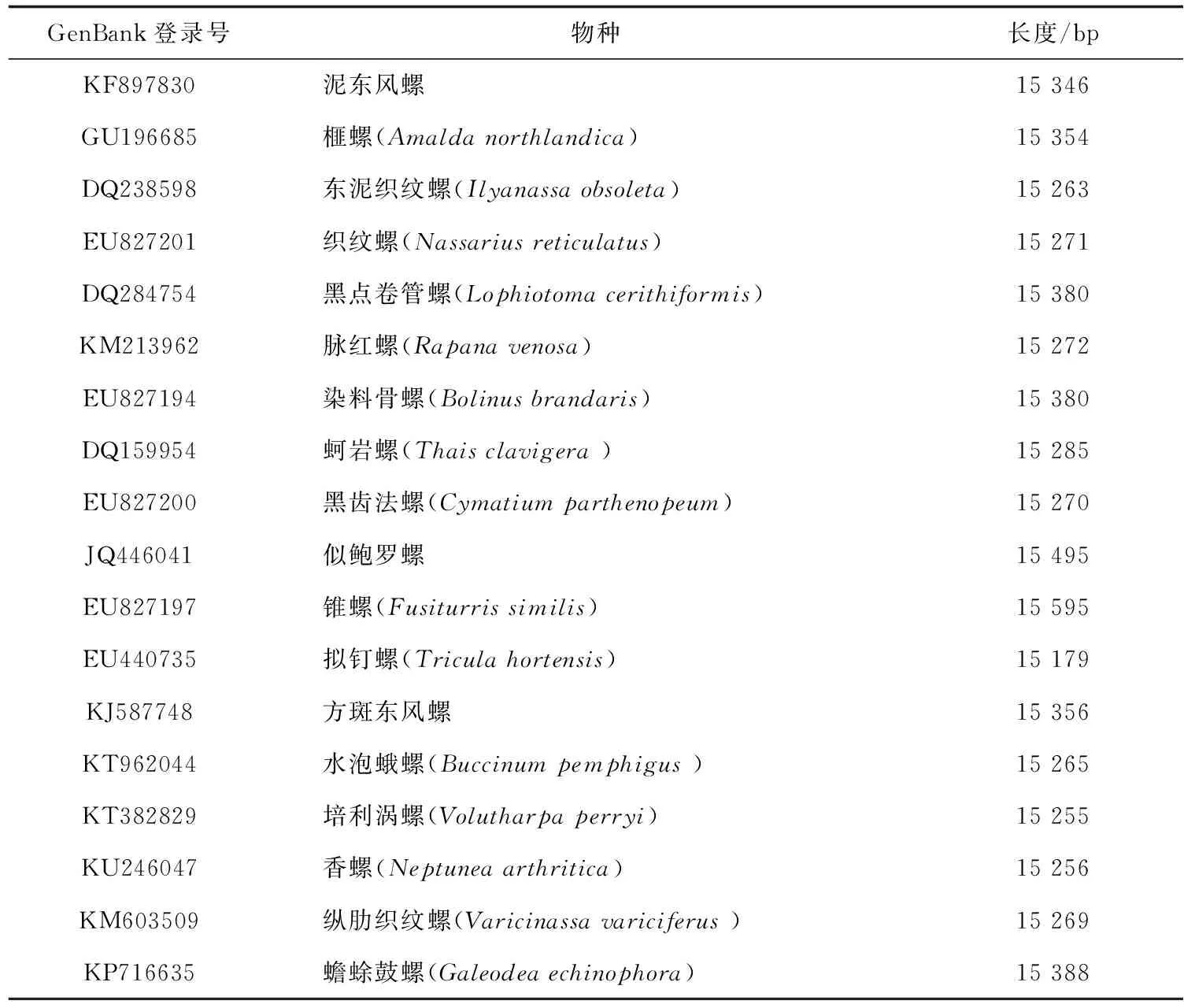
GenBank登录号物种长度/bpKF897830泥东风螺15 346GU196685榧螺(Amalda northlandica)15 354DQ238598东泥织纹螺(Ilyanassa obsoleta)15 263EU827201织纹螺(Nassarius reticulatus)15 271DQ284754黑点卷管螺(Lophiotoma cerithiformis)15 380KM213962脉红螺(Rapana venosa)15 272EU827194染料骨螺(Bolinus brandaris)15 380DQ159954蚵岩螺(Thais clavigera )15 285EU827200黑齿法螺(Cymatium parthenopeum)15 270JQ446041似鲍罗螺15 495EU827197锥螺(Fusiturris similis)15 595EU440735拟钉螺(Tricula hortensis)15 179KJ587748方斑东风螺15 356KT962044水泡蛾螺(Buccinum pemphigus )15 265KT382829培利涡螺(Volutharpa perryi)15 255KU246047香螺(Neptunea arthritica)15 256KM603509纵肋织纹螺(Varicinassa variciferus )15 269KP716635蟾蜍鼓螺(Galeodea echinophora)15 388

表3 腹足纲贝类线粒体基因组相似性 %
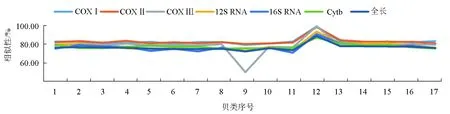
图2 泥东风螺与17种腹足纲贝类线粒体基因相似性1.榧螺;2.东泥织纹螺;3.织纹螺;4.黑点卷管螺;5.脉红螺;6.染料骨螺;7.蚵岩螺;8.黑齿法螺;9.似鲍罗螺;10.锥螺;11.拟钉螺;12.方斑东风螺;13.水泡蛾螺;14.培利涡螺;15.香螺;16.纵肋织纹螺;17.蟾蜍鼓螺.
2.3 系统进化分析
由Mega 5.1[20]软件最大似然法和邻接法构建的线粒体基因组系统发生树(图4)可知,蛾螺科中泥东风螺和方斑东风螺聚成一支,织纹螺科的织纹螺和纵肋织纹螺于蛾螺科内后聚成一支;圆口螺科的拟钉螺独立成支;榧螺和东泥织纹螺聚成一支,似鲍罗螺与骨螺科相似性较高;由这18种软体动物线粒体基因组构建的系统发育树看,线粒体基因组系统发育树与形态学分类有异。
2.4 序列变异特征分析
3个野生泥东风螺群体每个群体取3个个体扩增线粒体基因组全长序列,比对试验试验结果发现,在COXⅠ与COXⅡ基因相连区域有较多的变异位点。进一步对3个群体内各30个野生泥东风螺DNA样本以引物F5扩增测序,结果表明,变异位点分布在4067~4784 bp区域,共40个变异位点(图5),变异均为转换或颠换,没有检测到插入和缺失现象。
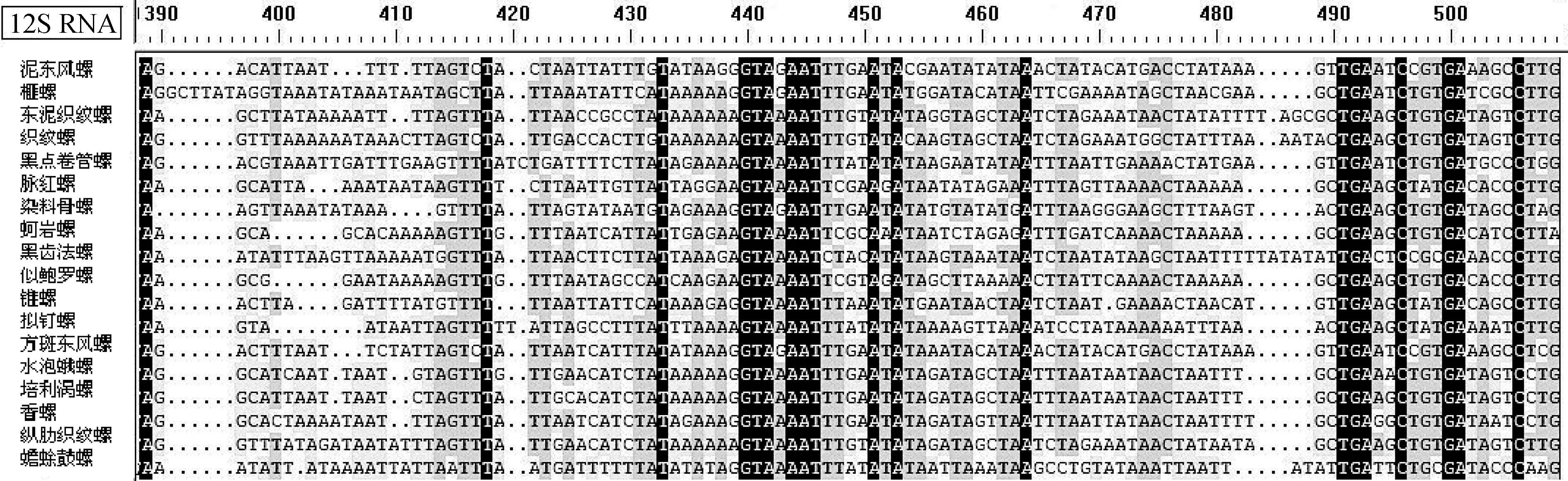



图3 腹足纲贝类12S RNA和16S RNA基因保守区和变异区
2.5 群体遗传多样性和遗传结构
泥东方螺各野生群体的单倍型分布(表4)和群体遗传距离(表5)分析表明,连江野生群体和北海野生群体中均检测到10个单倍型,而北海野生群体中检测到16个多态位点,3个野生群体的单倍型多样性为(0.406±0.112)~(0.532±0.113),3个野生群体的核苷酸多样性为(0.00094±0.00124)~(0.00111±0.00123);北海野生群体与连江野生群体和长乐野生群体之间的遗距离分别仅为0.00094和0.00099。

图5 F5引物扩泥东风螺线粒体基因组片段序列变异位点
种类年龄体质量/g体长/cm全长/cm鲢鱼2龄706.2±110.335.1±1.9542.7±2.12(n=9)510~86530.6~37.537.6~45.03龄1148.1±244.240.5±4.0549.0±4.64(n=8)925~168534.6~48.042.2~56.84龄2058.3±166.649.3±2.4759.5±3.10(n=6)1880~221047.2~52.056.8~62.9鳙鱼2龄858±70.235.64±0.9343.6±1.03(n=7)780~98534.2~36.941.8~45.23龄1641.7±143.444.82±0.8653.5±0.61(n=6)1415~181543.3~45.752.6~54.04龄3391.7±385.857.0±2.3868.6±2.92(n=5)2890~402553~59.464.3~72.05龄7495.7±510.374.1±4.4892.3±3.51(n=5)6921~789669.9~78.889~96
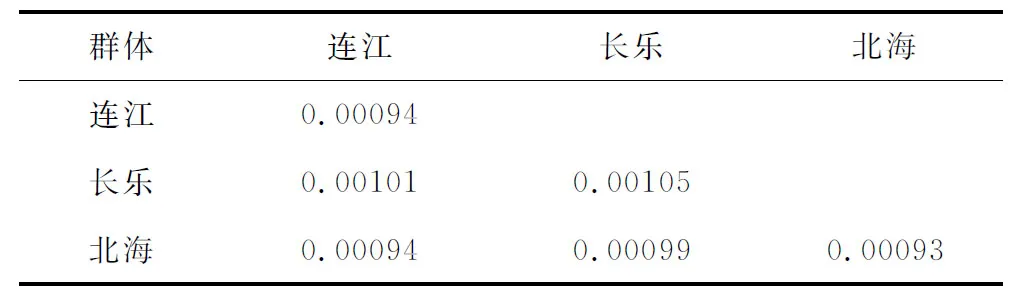
表5 泥东风螺群体遗传距离
3 讨 论
3.1 线粒体基因组遗传进化
对18种腹足纲动物线粒体基因组保守性分析发现,12S RNA的391~491 bp区域和16S RNA的1~106 bp和361~481bp区域为高度变异区,16S RNA的1191~1317 bp区域为高度保守区。泥东风螺作为后生动物,线粒体基因组中常含有多个长度不等的非编码区[22],通常认为在最大的非编码区中包含有线粒体基因复制和转录信号,这些非编码区被称为控制区[10],在其他水产动物中非编码控制区存在D-loop结构[17],但在本研究的泥东风螺线粒体基因组非编码区未发现D-loop结构。另外,泥东风螺线粒体基因组中也未发现在后生动物线粒体控制区中常见的较高AT含量的简单串连重复序列[23],这种结构在线粒体基因的复制和转录的早期阶段发挥着重要作用[23-24]。
3.2 泥东风螺野生群体遗传分化
遗传多样性是生物多样性的基础和物种进化潜能的保证,基因核苷酸多样性是衡量线粒体基因遗传多样性的重要指标。本研究采用16S RNA[25]和COXⅠ[26]基因通用引物对3个泥东风螺野生群体扩增片段测序,结果比对并未发现变异位点。但在比对3个群体中扩增的部分泥东风螺线粒体基因组全长序列后发现,COXⅠ和COXⅡ基因相邻区域附近存在较多变异位点,进一步用F5引物扩增测序发现,野生泥东风螺线粒体基因组变异位点主要分布在4067~4784 bp区域,3个野生群体在此片段共检测到40个变异位点,26个单倍型,整体遗传多样性较低;泥东风螺种内变异区间自COXⅠ基因的3′编码区段至COXⅡ基因的5′编码区,40个变异位点在COXⅠ基因3′编码区、COXⅡ基因5′编码区和COXⅠ与COXⅡ基因相邻的非编码区无明显的分布特征。通常物种间基因的非编码区为变异区,但在线粒体上各相邻基因的非编码区段非常短,有的相邻基因甚至共用编码区。如泥东风螺线粒体上基因存在12个基因共用编码片段,12个相邻基因间无非编码的碱基相隔,且相邻基因间的非编码区长度也为0~58 bp,可能是这种现象使得种内基因变异位点在非编码区分布不集中。考虑泥东风螺野生资源减少,因此推测野生群体遗传进化潜力水平受到了人工捕捞的影响,核苷酸变异和分化程度较低。连江和长乐野生群体之间的遗传距离相对北海群体较高的原因除地理因素之外还有人工捕捞因素,因此促进了两群体之间的基因交流。
由于所涉及野生样品数量限制,本研究只使用线粒体基因突变位点较多的片段序列进行分析,因此,下一步还需结合微卫星[27]和扩增片段长度多态性[28]指纹相关的分子标记技术,更全面地评估野生泥东风螺资源状况和泥东风螺人工增殖效果。本研究为利用线粒体进行泥东风螺群体的分子遗传学、遗传结构和种质鉴定提供参考依据。
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
数学物理学报(2022年5期)2022-10-09
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国卒中杂志(2021年7期)2021-11-29
河北画报(2020年8期)2020-10-27
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
医药前沿(2019年35期)2019-01-05
环球市场信息导报(2017年1期)2017-04-08
百科知识(2015年18期)2015-09-10
