
杉木人工林下原生与引入树种叶性状变异特征
2019-05-13 01:49邢文黎虞木奎成向荣
生态学报 2019年6期
熊 静,邢文黎,虞木奎,成向荣,*
1 中国林业科学研究院亚热带林业研究所,杭州 311400 2 南京林业大学,南京 210037
人工林在全球森林资源中扮演着重要角色,是缓解采伐天然林资源提供木材供给的有效补充,但普遍存在结构不尽合理、生产力不高、生态功能较弱和生态稳定性下降的问题[1]。目前,人工林的可持续经营理论与多目标经营的优化模式仍在不断实践探索中。杉木(Cunninghamialanceolata(Lamb.) Hook.) 是亚热带地区主要造林树种之一,其造林面积占世界人工林面积的6.5%,占我国人工林面积的19%,人工林蓄积量的25%,在我国林业生产中具有重要的地位[2]。随着杉木人工林集约经营发展,杉木连栽导致的土壤质量退化、林分生产力降低等问题日益突出[3- 4]。改变人工纯林单一林分结构,构建复层异龄林对改善林地土壤质量,提高林地生产力发挥了重要作用[5- 6]。
光是影响森林中林下植物生长和分布最主要的环境因素[7],是构建异龄复层林最主要的限制因子之一。植物对林下弱光环境的适应能力大小对其生长发育和生产力高低至关重要。叶片是植物物质生产、能量转换及交换的主要器官,其性状特征直接影响群落的结构和功能[8]。叶性状是植物的重要特性之一,属于植物功能性状的二级性状,具有相对的稳定性[9]。叶性状分为功能型性状和结构型性状,包括叶的形态、养分和渗透调节物质等[10],易测定且可同时对大量植物种类进行比较[11]。叶性状可直接影响植物的基本行为与功能[12- 13],能反映植物在表征生态系统功能方面的生态指示作用,强调其与生态系统过程及功能的关系[14]。在众多植物性状中,植物的一些叶片性状与植物的生存对策及植物利用资源的能力紧密联系,通常为适应环境变化而形成一系列生存对策[11],同时植物叶功能性状之间亦存在相关性,这反映出植物对环境的趋同适应特征[8]。随着全球环境及气候的变化,应用叶性状因子(如形态、解剖结构、化学计量等)研究植物在不同生境中的适应机制成为生理生态学领域近年来研究中的热点[15- 17]。目前对叶性状的研究主要集中在干扰和胁迫[12,18]、不同地区或流域特殊环境下植物叶性状间关系[19- 22]或集中在某一物种或生活型水平上叶性状的研究[12,22- 24],但叶性状在人工林林分结构改造上的探究较少。研究小区域小尺度不同生活型植物在形态、生理及光合作用方面的叶功能性状的差异性表达,有利于探究植物的适应机制,同时也有利于筛选与该地区植物生长及适应机制紧密联系的关键性状。本研究通过同质园试验在浙江省开化县林场杉木纯林下套种耐阴性伴生乔木和灌木树种,探讨其与林下生境过滤后自然更新的原生乔木与灌木树种之间叶性状差异性及相关性,以期为亚热带地区杉木复层异龄林伴生树种的筛选及其对林下环境适应机制解析提供一定理论参考与科学依据。
1 材料与方法
1.1 试验区概况
研究区位于浙江省开化县林场城关分场小桥头林区(118°25′E,29°09′N)(图1)。该区年日照总时数1334.1 h,年平均气温16.4℃,年平均降雨量1814 mm,无霜期252 d,气候属亚热带季风气候。土壤类型为红黄壤,pH为4.0—4.9。供试杉木纯林为1996年春季造林,初植密度为2500株/hm2,2006年进行1次抚育间伐,目前保留密度为1358株/hm2,平均树高和胸径分别为16.7 m和21.6 cm,林分郁闭度约0.85。
1.2 试验方法
2011年3月,在小桥头林区杉木人工林下引入13种耐阴树种(无外来种)构建杉木复层异龄林,树种详细信息见表1。灌木树种沿等高线栽植,密度3000株/hm2;乔木树种沿杉木行间栽植,密度2500株/hm2,具体树种配置方式见图1。造林苗木均采用1年生健壮容器苗,每个树种块状随机栽植于杉木林下,面积0.2—0.5 hm2。为比较引入树种与原生树种之间叶性状的关系,同时选择杉木林下自然生长的优势乔木与灌木(表1)。根据研究需要,将所有树种分为原生树种与引入树种;原生树种又分为原生乔木与原生灌木;引入树种分为引入乔木与引入灌木。
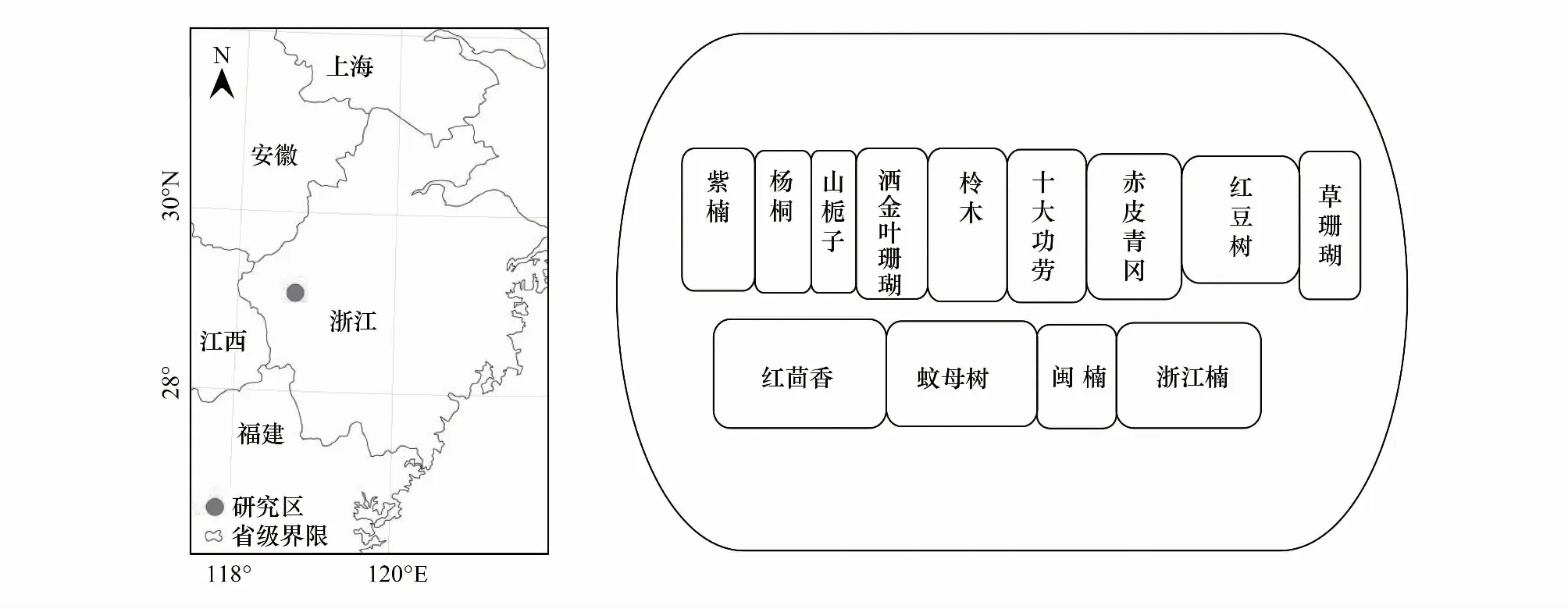
图1 试验区地理位置及引入树种栽植配置图Fig.1 Geographical location of the test area and planting configuration map of introduced tree species
1.3 叶性状测定
2017年8月在杉木人工林下分别选取引入和原生树种的当年生中上部小枝上的健康成熟叶片150—200片,避开林窗与林缘,采集的叶片编号后及时放入装有冰袋的保温箱内。将采集的叶片带回实验室后,一部分用于叶片形态指标分析,如叶长(Leaf length,LL) 、叶宽(Leaf width,LW)、叶长/叶宽(LL /LW)、叶面积(Leaf area,LA)等指标用LI-8400plus叶面积仪测定后采用WinFOLIA分析系统确定,扫描后的叶片烘干至恒重测叶干重(Leaf dry weight,LDW)并计算比叶面积(Specific leaf area,SLA)=扫描叶面积/扫描叶干重。剩余叶片用于生理和养分含量测定,叶绿素a(Chlorophyll a,Chla)、叶绿素b(Chlorophyll b,Chlb)含量采用乙醇-丙酮比色法测定;叶片淀粉(Starch,St)含量采用高氯酸水解-蒽酮比色法测量;可溶性糖(Soluble sugar,SS)采用蒽酮比色法量;非结构性碳水化合物(Non-structural carbohydrate,NSC)为淀粉与可溶性糖含量总和;叶碳(C)含量采用TOC分析仪测定;叶氮(N)含量用浓硫酸-过氧化氢消煮-半微量凯氏定氮法测定;叶片磷(P)含量采用HClO4-H2SO4消煮-钼锑抗比色法测定[25]。
1.4 数据处理
采用Excel 2013和SPSS 22进行数据分析和处理。以SPSS 22统计分析软件对实验数据进行方差分析、并对不同功能群进行Duncan多重比较检验,对叶性状的相关性进行Pearson相关分析,不同功能群叶性状的线性相关图采用对数函数转换后的数据进行绘制。以Canoco 4.5对叶性状数据进行主成分分析;表中数据为平均值(Mean)±标准差(Standard of deviation,SD);变异系数(Coefficient of variation,CV)=(标准差/平均值)×100%。研究区位置图采用ArcGIS 10.2绘制。

表1 引入树种与原生树种植物名录及功能群分类
2 结果与分析
2.1 原生树种与引入树种叶性状特征
根据原生树种与引入树种叶性状分析(表2)可知:总体上引入树种各叶形态参数数值略大于原生树种,其中引入与原生树种的叶宽、叶长宽比、叶面积与叶干重无显著差异(表2)。引入树种的叶长比原生乔木大32%(P<0.05),而与原生灌木无显著差异。引入树种比叶面积显著高于原生乔木(P<0.05),而与原生灌木无显著差异。无论是引入树种或原生树种,其叶面积、叶干重及比叶面积均表现出较高的变异性,其中原生乔木总体叶片形态指标变异系数普遍较低。
引入树种的总叶绿素含量显著高于原生树种(P<0.05)。引入乔木的总叶绿素含量显著高于引入灌木(P<0.05),原生灌木的总叶绿素含量显著高于原生乔木(P<0.05),引入乔木与原生灌木的总叶绿素含量无显著差异。引入乔木的叶绿素a含量显著高于其他树种(P<0.05),原生灌木的叶绿素b含量显著高于其他树种(P<0.05);引入树种与原生灌木的叶绿素a及叶绿素b含量的变异系数高于原生乔木。引入树种的叶绿素a/b显著大于原生植物(P<0.05),引入乔木和引入灌木的叶绿素a/b比值接近,分别比原生乔木和原生灌木高60%和45%。
引入树种叶碳含量显著高于原生树种(P<0.05),尤其是引入乔木叶碳含量最高。原生灌木叶氮含量最高,其次为引入乔木,原生乔木和引入灌木的叶氮含量没有显著差异。引入乔木和灌木树种与原生灌木的叶磷含量没有显著差异,但引入乔木和原生灌木叶磷含量显著高于原生乔木(P<0.05)。引入树种与原生树种的叶氮、磷含量的变异系数高于叶碳含量。引入树种与原生乔木的叶碳/氮均显著大于原生灌木(P<0.05),引入树种与原生灌木的叶碳/磷显著小于原生乔木(P<0.05),引入乔木与原生树种的叶氮/磷显著大于引入灌木(P<0.05)。
灌木树种的非结构性碳水化合物总量显著大于乔木树种(P<0.05),引入灌木显著小于原生灌木(P<0.05)。引入树种与原生乔木之间的淀粉含量差异不显著,但均显著小于原生灌木(P<0.05)。引入乔木与原生树种间的可溶性糖含量无显著差异,但均显著小于引入灌木(P<0.05)。引入树种的可溶性糖与淀粉含量的比值显著高于原生树种(P<0.05),其中引入灌木的可溶性糖与淀粉含量的比值取得最大值。引入树种与原生树种的可溶性糖及淀粉均表现出较高的变异性。
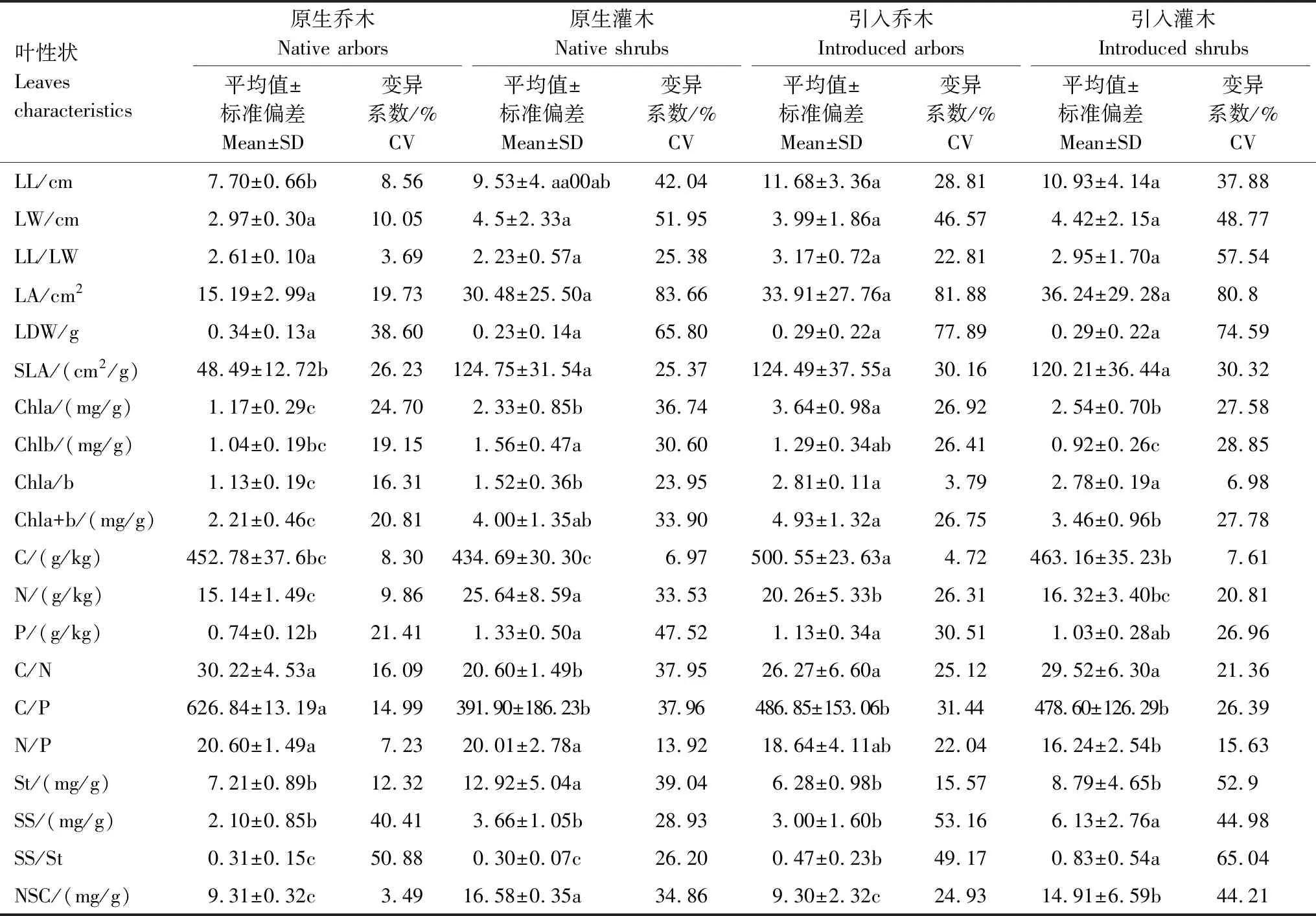
表2 原生树种与引入树种叶性状特征
不同小写字母表示同一叶性状在不同功能群之间具有显著差异(P<0.05);SD:标准偏差,Standard of deviation;CV:变异系数,Coefficient of variation;LL:叶长,Leaf length;LW:叶宽,Leaf width;LL/LW:叶长/叶宽,Leaf length/Leaf width;LA:叶面积,Leaf area;LDW:叶干重,Leaf dry weight;SLA:比叶面积,Specific leaf area;Chla:叶绿素a,Chlorophyll a;Chlb:叶绿素b,Chlorophyll b;Chla/b:叶绿素a/b,Chlorophyll a/b;Chla+b:叶绿素a+b,Chlorophyll a+b;C:碳,Carbon;N:氮,Nitrogen;P:磷,Phosphorus;C/N:碳/氮,Carbon/Nitrogen;C/P:碳/磷,Carbon/Phosphorus;N/P:氮/磷,Nitrogen/Phosphorus;St:淀粉,Starch;SS:可溶性糖,Soluble sugar;SS/St:可溶性糖/淀粉,Soluble sugar/Starch;NSC:非结构性碳水化合物,Non-structural carbohydrate
2.2 原生树种与引入树种的叶性状相关性
引入树种与原生树种的叶性状具有显著的正相关关系(R2=0.2354,P=0.032,图2),其中原生乔木与原生灌木(R2=0.96,P<0.001,图2)、引入乔木与引入灌木(R2=0.9789,P<0.001,图2)的叶性状之间呈高度线性正相关,表明原生树种之间、或引入树种之间叶性状的相关度均较强。引入乔木与原生乔木(图2)、引入乔木与原生灌木(图2)、引入灌木与原生乔木(图2)、引入灌木与原生灌木(图2)的叶性状之间也呈现良好的正相关关系(P<0.05),表明在杉木人工林下乔木与灌木的叶性状特征变化趋势类似。
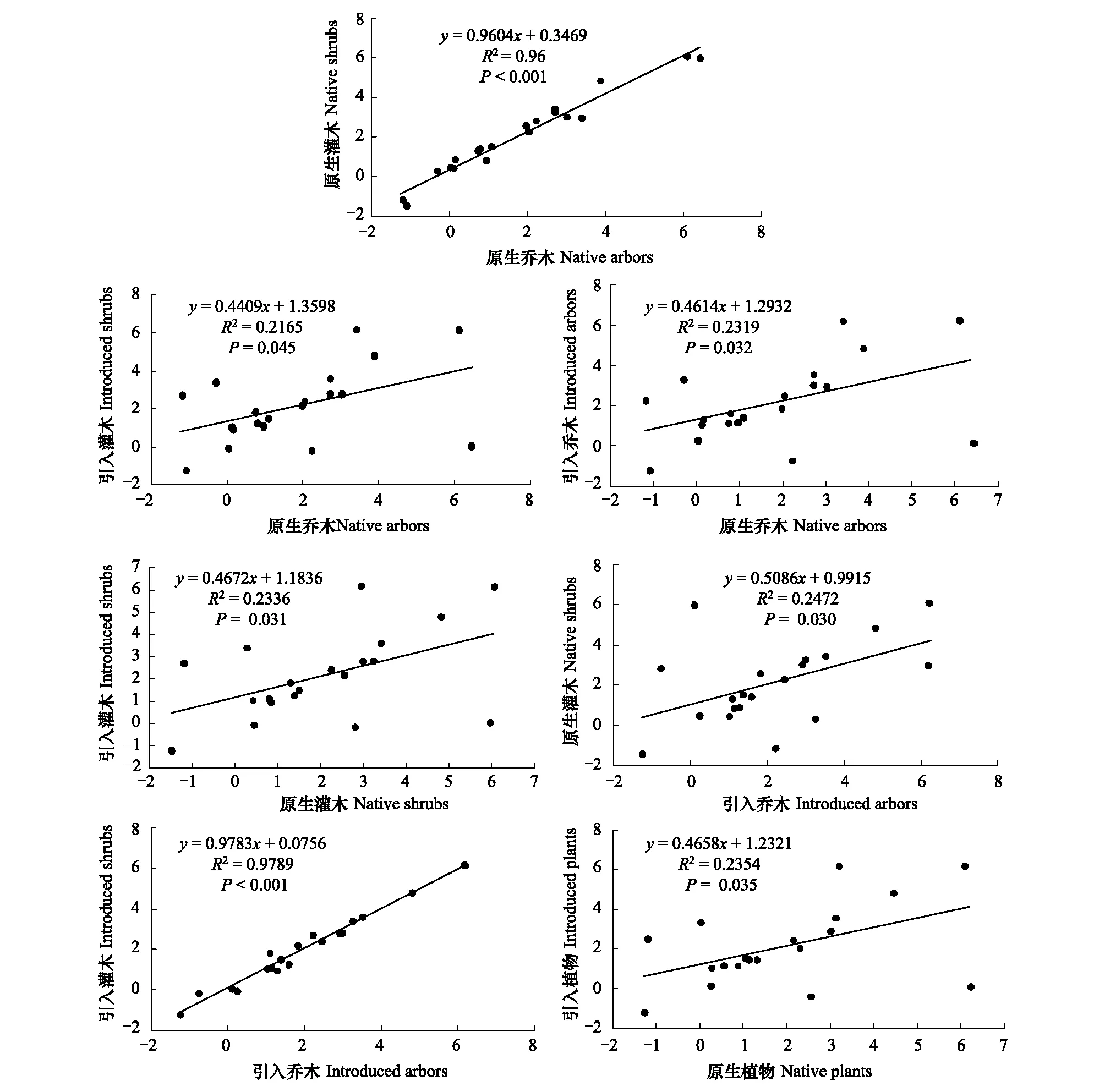
图2 不同功能群间叶性状的线性关系Fig.2 Linear relationship of leaf traits among different functional groups
从表3可以看出,总体上引入乔木与原生乔木的叶绿素含量与叶长宽比及叶干重无显著相关性,而与叶长、叶宽、叶面积及比叶面积呈显著正相关(P<0.05)。叶片养分含量中,叶碳、氮、磷含量与比叶面积呈极显著正相关(P<0.01),与色素含量呈显著正相关(P<0.05),叶碳/氮、叶碳/磷、叶氮/磷与比叶面积、叶绿素含量呈极显著负相关(P<0.01)。非结构性碳水化合物中可溶性糖含量与其他叶性状无显著相关性,叶片淀粉含量、总非结构性碳水化合物含量与色素含量、叶氮含量和叶磷含量呈极显著负相关(P<0.01),与比叶面积呈显著负相关(P<0.05)。
引入灌木与原生灌木间Pearson相关性分析显示:总体上叶绿素含量与叶形态性状呈显著正相关(P<0.05),而与叶干重无显著相关性。叶片养分含量中,叶碳含量与叶长宽比呈极显著负相关(P<0.01),而与其他形态性状及色素含量无显著相关性;叶氮含量和叶磷含量与叶宽、比叶面积及色素含量呈极显著正相关(P<0.01);总体上叶碳/氮、叶碳/磷、叶氮/磷与叶形态性状及叶绿素含量呈显著负相关(P<0.05)。非结构性碳水化合物与比叶面积、叶绿素a+b含量无显著相关性;淀粉与叶形态性状及叶绿素a含量呈负相关(P<0.05);可溶性糖含量与叶绿素b含量呈显著负相关(P<0.05);非结构性碳水化合物总量与叶绿素b含量、叶氮含量、叶氮/磷呈显著正相关,与叶绿素a/b、叶碳含量、叶碳/氮呈显著负相关(P<0.05)。
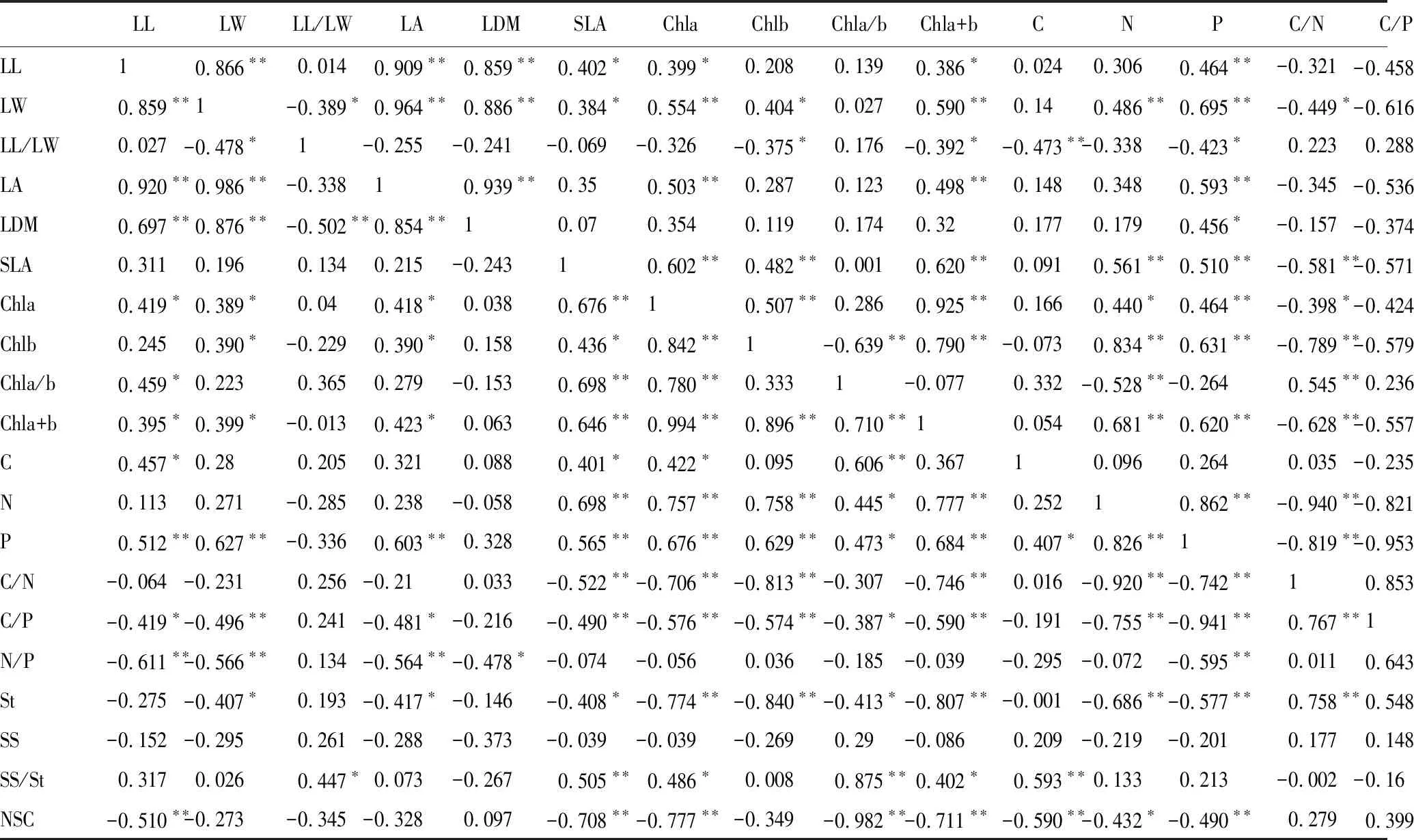
表3 乔木之间及灌木之间叶性状Pearson相关性
表左下角为原生乔木与引入乔木间Pearson相关性,右上角为原生灌木与引入灌木间Pearson相关性。表中*表示在0.05水平上达到显著水平(P<0.05),**表示在0.01水平上达到极显著水平(P<0.01)
基于叶性状和功能群的主成分分析(PCA)表明,前2个主成分可以解释叶性状变异总方差的87.9%,其中PC1可以解释总方差的48.8%,PC2可以解释总方差的39.1%(图3)。叶长、叶宽、叶面积、比叶面积、叶绿素a、叶绿素a+b、可溶性糖等对PC1轴的贡献率较大;叶碳/氮、叶长宽比、叶碳含量、叶绿素b、淀粉、总非结构性碳水化合物总量等对PC2轴的贡献率较大。引入乔木(IA)与引入灌木(IS)树种在叶碳含量、叶绿素a/b、可溶性糖、叶绿素a、叶面积等与其他功能群植物差异较大。原生乔木(NA)在叶碳/磷、叶碳/氮、叶干重等与其他功能群植物差异较大。原生灌木(NS)在淀粉、叶绿素b、叶氮、叶磷含量等与其他功能群植物差异较大。引入乔木与引入灌木的叶性状较为接近;引入树种与原生灌木的叶性状差异相对较小,而与原生乔木差异较大;原生乔木与原生灌木树种的叶性状差异相对较大。
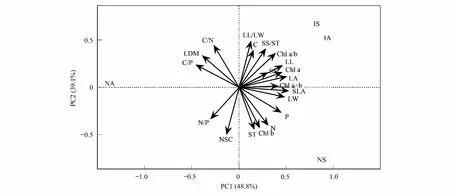
图3 原生植物与引入植物叶性状的主成分分析Fig.3 Principal component analysis (PCA) of traits of native and introduced plantsNA:原生乔木,Native arbors;NS:原生灌木,Native shrubs;IA:引入乔木,Introduced arbors;IS:引入灌木,Introduced shrubs
3 讨论
植物叶性状受环境的影响很大[26],而植物自身作为自组织有机体,对外界环境适应并非单一性状的适应,通常是和其他性状协同适应[27]。形态性状是植物适应环境变异最直接的外在表现,比叶面积则是植物功能生态学研究中最常用性状,是植物比较生态学研究中的首选指标[11- 12],可以反映植物获取资源能力,亦可作为叶遮阴度的指数使用[28]。通过对山栀子(Gardeniajasminoides)和浙江楠(Phoebechekiangensis)在不同光照下叶片特征研究发现,比叶面积随有效光照强度降低而显著增加[29- 30]。此外,一般在低光环境下生长的植物,会出现叶片更加宽大,叶长宽比也往往较小的现象[31]。随着杉木林分郁闭度增加,林下植物自然更新受到弱光限制,从而发生生境过滤作用,通过长期适应与环境选择出叶性状与林下弱光环境相适应的原生树种。本研究中引入乔木的叶长及比叶面积显著大于原生乔木,表明其受林下弱光环境影响,引入乔木树种通过增加比叶面积协同叶长调整,来加强捕获光能,以促进光合作用和维持自身的正常生长发育。本研究表明引入乔木与引入灌木的比叶面积没有显著差异,Chen等[32]、刘贵峰等[14]研究也发现乔灌木间的比叶面积没有明显差异;而原生灌木的比叶面积大于原生乔木,这与吕金枝等[33]对山西霍山30种不同功能型植物间的叶面积关系一致。本研究中引入树种与原生树种比叶面积变化的差异可能与引入灌木在低光环境的自我调节能力的大小和适应策略有关。
变化环境中,植物主要通过两条适应策略,其一为改变叶性状,其二则调整植物性状间的关系[34]。叶绿素与植物光合作用密切相关,本研究中引入乔木的叶绿素a及总叶绿素含量显著高于原生乔木,说明引入乔木通过增加叶绿素a含量加快光合产物的合成提高自身生产力,以及协同较高的叶面积与比叶面积来适应林下弱光环境,这与梯度弱光胁迫下杨桐(Adinandramillettii)及紫楠(Phoebesheareri)的生理响应一致[35- 36]。然而有研究指出随着光照减弱,光合色素调节主要以叶绿素b的提高为主[37],低光环境中耐阴植物叶绿素b含量增加的幅度往往高于叶绿素a,从而形成较低的叶绿素a/b,以提高植物对蓝光的利用效率[38]。本研究中灌木树种之间叶形态性状及叶绿素a含量无显著变化,这可能反映了引入灌木树种受林下弱光环境的影响较小,但引入灌木的叶绿素b含量显著低于原生灌木,说明引入灌木与原生灌木的比叶面积—叶绿素关系格局发生了位移现象,引入灌木对林下弱光的捕获和利用能力小于原生灌木,原生灌木的适应性可能更强。
碳是植物体内干物质最主要的构成元素,叶碳含量高则表明其比叶重大,光合速率低,生长速率慢,具有较强的防御能力[39]。本研究中引入树种的叶碳含量显著大于原生树种,根据叶碳含量适应策略可初步将耐阴性排序为:引入乔木<原生乔木、引入灌木<原生灌木。叶片氮、磷含量是植物与环境长期适应结果,研究表明在弱光环境中植物可通过增加氮在捕光组分中的含量和分配来提高光能捕获能力,或通过增加磷的投资比例来增大细胞的分裂能力,进而获得较大的比叶面积[37]。本研究中比叶面积、叶绿素、叶氮含量、叶磷含量之间的极显著正相关关系在不同功能群中变化趋势一致,表明林下植物主要通过形态适应或生理调节促进比叶面积及色素含量的增加以适应林下弱光环境。氮和磷是陆地生态系统植物生长的主要限制因子,可以直接反映植物受养分胁迫的状态[40]。一些研究者采用氮/磷来诊断植物的元素受限状况[41],在对欧洲湿地植物的研究发现,当氮/磷<14,植物受氮制约,当氮/磷>16,植物受磷制约[42]。本研究中,大多数植物的氮/磷>16,说明该地区植物主要受磷制约;引入树种的氮/磷显著小于原生树种,表明引入树种的氮、磷内稳性可能较原生树种强。有研究表明,内稳性较弱的树种的适应性更强[43],进一步说明原生树种的适应性高于引入树种;而内稳性较高的树种的养分利用方式可能更加保守[44],因而内稳性较高的引入树种在弱光环境中更倾向于生存而非生长,这种保守性策略可能是导致其光合产物低于原生树种的主要原因。
非结构性碳水化合物是植物光合作用的主要产物,包括淀粉和可溶性糖,不仅可以反映植物碳收支状况[45],亦可表征植物生长和存活的能力以及应对外界胁迫的缓冲能力[46]。淀粉是植物主要的长期能量储存物质之一,是植物储存器官中最重要的碳水化合物[47]。可溶性糖是光合作用的直接产物,是植物通过生理调节适应环境变化的重要机制[48]。引入乔木与原生乔木的淀粉、可溶性糖含量及非结构性碳水化合物总量无显著差异,但引入乔木的可溶性糖含量与淀粉的比值显著大于原生乔木,说明引入乔木主要以可溶性糖形式参与机体代谢及抵御外界胁迫为主,而原生乔木的光合产物则以淀粉的形式用于能量储存及自身生长,这一结论辅证了引入乔木的耐阴性小于原生乔木。引入灌木与原生灌木的非结构性碳水化合物总量接近,但引入灌木的可溶性糖含量与淀粉的比值显著大于原生灌木,表明引入灌木的耐荫性较原生灌木弱,其亦发动了可溶性糖调节机制,通过增加可溶性糖含量来调节渗透压、缓冲弱光胁迫压力。乔木一般处在森林群落中的优势层,接收到的光照最强,光合作用也最为充分,植物叶片的碳同化能力最大,而灌木处于次优势层,由于乔木层的遮挡其碳同化能力较弱,因此灌木的非结构性碳水化合物较乔木低[49- 50]。本研究中不论是引入树种或原生树种,其中乔木非结构性碳水化合物均显著低于灌木,这可能是引入或原生的乔木与灌木树种均处在林下层,灌木能够进行正常生长,而乔木并未处在适宜的生态位上,林下光照不足制约了光合作用。
在植物生长发育过程中,由于受到物理、生理等因素的综合作用,不同叶性状间存在相互促进或制约关系[51]。本研究也发现,引入树种与原生树种的叶性状呈显著线性相关,乔木或灌木的叶绿素含量与叶形态性状、叶氮、叶磷含量与叶绿素含量均呈显著正相关,其淀粉含量均与比叶面积呈显著负相关,说明相同生活型植物叶性状对弱光环境的响应具有高度相似性[52],表现出较高的趋同适应特征。另外植物类群和生活型对性状分异产生一定影响[53],PCA排序结果显示叶性状在引入乔木与引入灌木间的差异较小,但原生乔木与原生灌木间叶性状的差异较大,这反映了不同生活型植物的叶性状仍具有一定差异。
4 结论
本研究探讨了杉木人工纯林改造为杉木复层异龄林7年后,林下引入乔灌木树种与原生乔灌木树种的叶性状变异特征,研究发现引入树种与原生树种的叶形态性状差异较小,引入树种及原生灌木的比叶面积及色素含量显著高于原生乔木,引入树种主要通过高比叶面积—叶绿素a来适应林下弱光环境,原生树种主要采用增加比叶面积—叶绿素b适应策略。引入乔木的叶碳氮磷含量均大于引入灌木,而原生乔木均小于灌木,引入与原生树种均受土壤磷元素制约。引入灌木的可溶性糖含量最高,原生乔木的淀粉含量最高,灌木的非结构性碳水化合物显著大于乔木,灌木之间光合产物较为接近,乔木之间无显著差异。引入树种与原生树种的叶性状高度相关,表现出趋同适应特征。引入乔木与灌木的叶性状较为接近,引入树种与原生乔木和灌木叶性状存在一定差异,这不仅与植物遗传特性密切相关,也与植物耐阴性和所处生态位有关。综合叶性状分析,本研究中引入灌木树种的适应性优于乔木树种。因此,在研究区构建杉木异龄复层林需根据林木生态适应特征,进行适当间伐,降低林分郁闭度,将有利于林下植被层的快速恢复,尤其是伴生乔木树种需要更多的光照资源和生长空间。
猜你喜欢
建筑与预算(2022年6期)2022-07-08
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
农村实用技术(2020年1期)2020-12-16
新疆农业科学(2020年1期)2020-02-14
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
乡村科技(2018年8期)2018-02-13
现代园艺(2018年14期)2018-01-18
诗潮(2017年2期)2017-03-16
中国麻业科学(2015年5期)2015-12-28
学与玩(2009年2期)2009-03-09
