
水稻锌指蛋白基因CRISPR/Cas9突变体的构建及突变分析
2020-02-22 07:37易勇郑瑞杨波栾维江郭嗣斌沙爱华
南方农业学报 2020年11期
易勇 郑瑞 杨波 栾维江 郭嗣斌 沙爱华

摘要:【目的】通过CRISPR/Cas9基因编辑技术对水稻锌指蛋白基因(OsC3H54)进行基因编辑,筛选鉴定出其突变体植株,为深入研究OsC3H54的生物学功能提供良好材料,也为水稻锌指蛋白研究提供参考依据。【方法】通过E-CRISP在OsC3H54基因的外显子上设计靶点序列,将靶点序列连接至OsU6SK载体上,再与Cas9一起连接到pCAMBIA1300双元载体上,获得CRISPR/Cas9重组双元载体,通过农杆菌介导将其转入日本晴水稻愈伤组织,利用潮霉素进行抗性筛选,获得突变体植株,并分析其靶点位置的碱基及编码氨基酸突变情况。【结果】在OsC3H54基因第2个外显子上找到2个符合靶点设计要求的靶点,分别为TG1:5'-CCGCCGCGGCTGCCTTTGGATAC-3'和TG2:5'-CCTTCCC CAATGGCGGGGGTGGC-3'。将OsU6SK载体和靶点序列正确连接的重組载体与Cas9一起连接至pCAMBIA1300双载体上,成功获得CRISPR/Cas9重组双元载体(pCAMBIA1300-Cas9-TG1和pCAMBIA1300-Cas9-TG2)。通过农杆菌介导转入日本晴水稻,经潮霉素抗性筛选获得TG1靶点株系和TG2靶点株系,共16株CRISPR/Cas9突变体植株。CRISPR/Cas9突变体植株在靶点序列的突变位点位置附近出现套峰,表明2个株系的植株均发生碱基突变,其中Y1、Y2、Y3和Y4突变体植株均为单碱基插入突变,最终导致编码的氨基酸发生移码突变,蛋白翻译提前终止。【结论】水稻OsC3H54基因CRISPR/Cas9突变体植株的获得为进一步研究水稻锌指蛋白生物学功能提供了良好材料。
关键词: 水稻;锌指蛋白;C3H54;CRISPR/Cas9;基因编辑;突变体植株;突变分析
中图分类号: S511.035.3 文献标志码: A 文章编号:2095-1191(2020)11-2607-07
CRISPR/Cas9 mutants construction of rice zinc finger protein gene and mutation analysis
YI Yong1, ZHENG Rui2, YANG Bo2, LUAN Wei-jiang2, GUO Si-bin3, SHA Ai-hua1*
(1College of Agriculture,Yangtze University/Hubei Collaborative Innovation Center for Industrialization of Major Grain Crops, Jingzhou, Hubei 434025, China; 2College of Life Science,Tianjin Normal University, Tianjin 300387, China;3Rice Research Institute, Guangxi Academy of Agricultural Sciences/Guangxi Key Laboratory
of Rice Genetics and Breeding, Nanning 530007, China)
Abstract:【Objective】The rice zinc finger protein gene(OsC3H54) was genetically edited based on CRISPR/Cas9 gene editing technology, and the mutant plants were screened and identified, which would provide materials for in-depth study of the biological functions of OsC3H54 as well as reference for the research of rice zinc finger protein. 【Method】The target sequence was designed on the exon of rice OsC3H54 gene through E-CRISP, and the target sequence was liga-ted to the OsU6SK vector to obtain the target link vector. Then the target link vector together with Cas9 were integrated to pCAMBIA1300 to obtain the CRISPR/Cas9 recombination binary vector, which was then transformed into Nipponbare rice callus with Agrobacterium-mediated method. The positive transformed plants were screened out using hygromycin, and the mutated bases at the target site and encoded amino acids were analyzed. 【Result】Two targets were found on the second exon of OsC3H54, which met the target design requirements, named TG1:5'-CCGCCGCGGCTGCCTTTGG ATAC-3' and TG2: 5'-CCTTCCCCAATGGCGGGGGTGGC-3'. The OsU6SK vector and recombinant vector with correct target sequence and the empty Cas9 vector were ligated to the pCAMBIA1300 binary vector, and the CRISPR/Cas9 recombinant binary vector(pCAMBIA1300-Cas9-TG1 and pCAMBIA1300-Cas9-TG2) was successfully obtained. The CRISPR/Cas9 recombinant vectors were transferred to Nipponbare based on Agrobacterium-mediated method with hygromycin resistance for transforms screening. A total of 16 CRISPR/Cas9 mutant lines of TG1 and TG2 target were obtained. A set of peaks near the mutation site of the target sequence in the CRISPR/Cas9 mutant lines were detected, indicating that base mutations happened in two transformed lines. Among them,the Y1,Y2,Y3 and Y4 mutant lines were all single-base insertions mutations that led to premature termination of amino acid traanslation with frameshift mutations. 【Conclusion】The obtained CRISPR/Cas9 mutant plant of OsC3H54 provides good materials for biological functions studying of rice zinc finger protein.
Key words: rice; zinc finger protein; C3H54; CRISPR/Cas9; gene editing; mutant plants; mutation analysis
Foundation item:National Key Research and Development Program of China (2018YFD0301301-5-2); Guangxi Natural Science Foundation (2017GXNSFDA198039); Guangxi Key Laboratory of Rice Genetics and Breeding Open Pro-ject (No.160-380-16-1)
0 引言
【研究意义】水稻(Oryza sativa)是全球主要的粮食作物之一,世界上近一半人口以稻米为主食,其对我国农业的可持续发展也发挥着举足轻重的作用(刘凯等,2019)。锌指蛋白是一类通过与Zn2+结合形成短且稳定、能自我折叠成“手指”形状结构域的蛋白,其结构域由3个半胱氨酸和1个组氨酸组成,在植物体内主要作为转录因子对基因表达发挥重要作用(刘丹等,2019)。该蛋白家族拥有众多成员,主要调控植物生长发育。C3H54是锌指蛋白家族的成员之一,克隆其编码基因并构建基因编辑突变植株,对研究锌指蛋白在水稻生长发育过程中的调控作用具有重要意义。【前人研究进展】锌指蛋白最早在非洲爪蟾中发现(Miller et al.,1985)。根据锌指结构功能及序列的不同,可将锌指蛋白分成9种类型:C2H2、C2HC、C2HC5、C3H、C3HC4、C4、C4HC3、C6和C8(Berg and Shi,1996),其中C3H型锌指蛋白的研究较少(刘小艳等,2015)。目前已有较多关于水稻锌指蛋白的研究报道:OsDOS是第一个在水稻中发现的锌指蛋白基因,其在水稻植株中过表达可延缓叶片衰老(Kong et al.,2006);通过水稻全基因组分析共发现67个C3H基因,通过构建系统发育进化树可将其分为8个亚家族(Wang et al.,2008a);水稻OsLIC基因能负调控油菜素内酯合成,其过表达会影响植株的形态建成(Wang et al.,2008b);C3H12基因编码的锌指蛋白是一种核酸结合蛋白,可通过茉莉酸途径正向调节水稻白叶枯病抗性(Deng et al.,2012);水稻OsTZF1基因过表达不仅可延缓水稻叶片衰老,还能阻滞水稻种子的萌发和生长(Jan et al.,2013);水稻Ehd4基因编码一种新型锌指蛋白,可调控植株光周期,进而影响开花,可导致长日照条件下植株不开花,短日照条件下植株可提前开花(Gao et al.,2013);水稻锌指蛋白基因OsZFP1在水稻中过量表达可提高植株稻瘟病抗性(李贺等,2015);非典型的串联锌指蛋白IIP4不仅能与促进水稻次生壁生成的NAC29/NAC31上游调节因子相互作用抑制下游调节因子,从而抑制次生壁的形成, 还可调控植株的机械强度(Zhang et al.,2018a)。此外,目前已有较多其他植物锌指蛋白的研究报道:橡胶树HbCZF1是一种非典型且具有C-X7-C-X5-C3-HCCCH基序和RNA识别基序的锌指蛋白,高度表达可激活hmg1基因的转录,参与天然橡胶的生物合成(Guo et al.,2015);拟南芥AtTZF5通过TZF基序与调控脱落酸(ABA)调节因子相互作用,在种子中高度表达并参与ABA信号转导(Bogamuwa and Jang,2016);辣椒CaC3H14基因上调表达可增强植株对病菌的抗性(Qiu et al.,2018);在甘蓝型油菜中C3H型转录因子BnZFP1与高油酸性状相关,其过表达可使油酸含量提高18.8%(Zhang et al.,2018b);甘薯IbC3H18是一种非串联的锌指蛋白,可作为核酸转录激活因子,其过表达增强甘薯的耐盐性和抗旱性(Zhang et al.,2019)。【本研究切入点】至今未见水稻C3H54锌指蛋白的相关研究报道。【拟解决的关键问题】通过CRISPR/Cas9基因编辑技术对水稻C3H54基因(OsC3H54)进行基因编辑,获得其稳定遗传突变体株系,并分析其靶点位置的碱基及编码的氨基酸突变情况,为深入研究OsC3H54的生物学功能提供良好材料,也为水稻锌指蛋白研究提供参考依据。
1 材料与方法
1. 1 试验材料
供试水稻品种为日本晴,由天津师范大学生命科学院提供。主要试剂:Bsa I酶购自纽英伦生物技术(北京)有限公司;质粒提取试剂盒购自天根生化科技(北京)有限公司,其他生化试剂购自武汉汉宇飞扬科技有限公司。主要仪器设备:SYSTEM GelDoc XR+IMAGELA-凝胶成像系统(Bio-Rad,美国)、T100TM Thermal Cycler PCR仪(Bio-Rad,美国)、Centrifuge 5430R离心机(Eppendorf,德国)和JY600C电泳仪(北京君意东方电泳设备有限公司)。
1. 2 靶点选择
在E-CRISP输入OsC3H54基因的编码区序列(CDS),找到含有20个碱基的靶点序列GN19NGG,其中NGG是识别靶点序列的原初间隔序列毗邻基序(Protospacer adjacent motif,PAM),N代表T、G、C或A,靶点序列位置必须设计在同一外显子上。通过E-CRISP评估分析靶点序列的打靶效率,从而选出合适的靶点序列。最后,利用NCBI数据库的BLAST对靶点进行特异性分析。
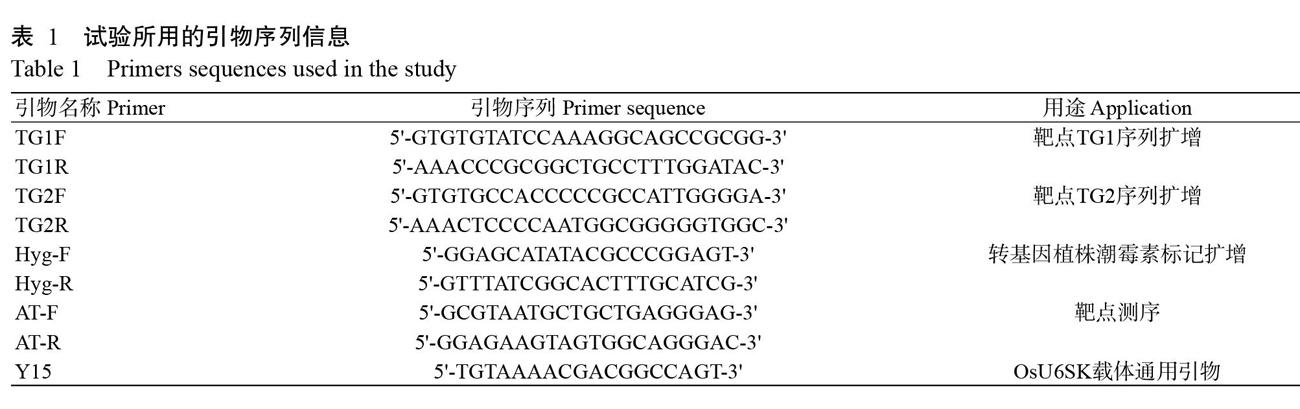
1. 3 CRISPR/Cas9重组双元载体构建
具体操作:(1)根据引物互补性,分别利用靶点引物TG1F/TG1R和TG2F/TG2R(表1)通过引物低温退火配对方式获得靶点序列TG1和TG2的Oligo二聚体。PCR扩增程序:95 ℃ 3 min;0.1 ℃/s速率降至22 ℃。(2)用Bsa I酶切OsU6SK载体过夜,回收载体片段,将其分别与靶点序列TG1和TG2连接,转化大肠杆菌DH5α感受态细胞。挑选单菌落,利用引物Y15(表1)和靶点序列下游引物(TG1R或TG2R)分别进行菌落PCR鉴定。PCR反应体系20.0 μL:10×Buffer 2.0 μL,Y15引物和靶点序列的下游引物各1.0 μL,5 U/μL Taq DNA聚合酶0.2 μL,2.5 mmol/L dNTP 1.5 μL,单克隆菌液1.0 μL,ddH2O补足至20.0 μL。扩增程序:94 ℃预变性4 min;94 ℃ 30 s;55 ℃ 30 s;72 ℃ 30 s,进行35个循环;72 ℃延伸7 min,12 ℃保存。挑選阳性单克隆菌株送至生工生物工程(上海)股份有限公司测序。将OsU6SK载体和靶点序列正确连接的重组载体与Cas9一起连接至pCAMBIA1300双元载体上,转化大肠杆菌DH5α感受态细胞,挑取单菌落,提取其质粒进行酶切鉴定,确保获得正确的CRISPR/Cas9重组双元载体(孔晓聪等,2019)。
1. 4 突变体植株的构建及分子鉴定
将构建的重组双元载体转化农杆菌后,通过农杆菌介导法侵染日本晴水稻愈伤组织,置于含潮霉素的培养基上培养,筛选出阳性愈伤组织,转至分化培养基上培养,再转至生根培养基上培养,最终获得突变体植株。将突变体植株种植于试验大田中,正常田间管理,待植株长大后收集幼嫩叶片,采用CTAB法提取其DNA,用潮霉素标记的特异引物Hyg-R和Hyg-F(表1)进行PCR鉴定,反应体系和扩增程序同1.3中的菌落PCR鉴定。
1. 5 突变体植株突变类型分析
根据OsC3H54基因序列设计能特异性检测CRISPR/Cas9的引物(AT-F/AT-R),其中上游引物AT-F在靶点序列上游约250 bp处,下游引物AT-R在靶点序列下游约250 bp处(表1)。利用AT-F/AT-R进行PCR扩增,模板为1.4中提取的突变体植株叶片DNA,反应体系同1.3中的菌落PCR鉴定;扩增程序略有修改,即退火温度和时间改为52 ℃和30 s,其余条件不变。PCR扩增产物经1%琼脂糖凝胶电泳验证后送至生工生物工程(上海)股份有限公司进行测序。测序结果用DNAMAN 6.0进行分析。
2 结果与分析
2. 1 靶点选择结果
由图1可知,OsC3H54基因含有7个内含子、8个外显子。依据靶点设计原则,结合OsC3H54基因的结构特点,在该基因的第2个外显子上找到符合要求的2个靶点。利用E-CRISP评估这2个靶点的打靶效率,再利用NCBI数据库中的BLAST对靶点特异性进行分析,结果显示这2个靶点的打靶效率和特异性均较好,可作为OsC3H54基因的编辑靶点,分别为TG1:5'-CCGCCGCGGCTGCCTTTGGATAC-3'和TG2:5'-CCTTCCCCAATGGCGGGGGTGGC-3'。
2. 2 CRISPR/Cas9重组双元载体构建結果
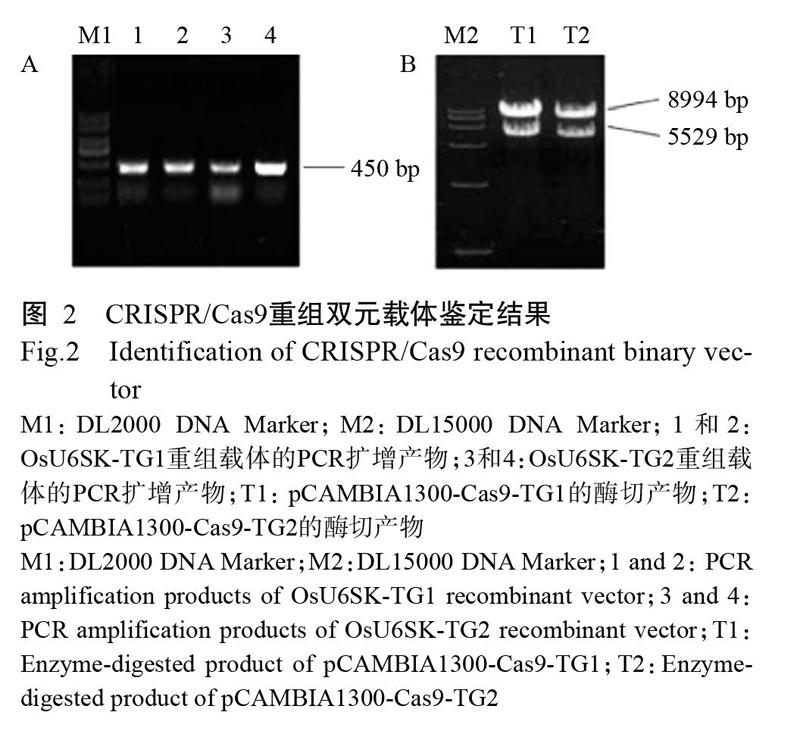
将靶点序列片段TG1和TG2分别与OsU6SK载体连接,然后转化大肠杆菌DH5α感受态细胞,挑取单菌落利用引物Y15(表1)和靶点序列下游引物(TG1R和TG2R)进行PCR鉴定,PCR扩增产物用1%琼脂糖凝胶电泳进行检测,成功获得2个含有靶点序列的重组载体(OsU6SK-TG1和OsU6SK-TG2),长度均450 bp(图2-A)。将测序正确的重组载体与Cas9一起连接至pCAMBIA1300双元载体上,并转大肠杆菌DH5α感受态细胞,提取其质粒进行酶切鉴定,酶切产物用1%琼脂糖凝胶电泳进行检测,结果(图2-B)显示,酶切产物大小分别为8994和5529 bp,与预期结果相符,表明pCAMBIA1300双元载体中已插入含靶点序列的重组载体和Cas9,成功获得含TG1和TG2靶点序列的CRISPR/Cas9重组双元载体(pCAMBIA1300-Cas9-TG1和pCAMBIA1300-Cas9-TG2)(图3)。
2. 3 CRISPR/Cas9突变体植株分子鉴定结果
将CRISPR/Cas9重组双元载体转化农杆菌,挑取阳性菌侵染日本晴水稻愈伤组织,将侵染成功的愈伤组织置于含有潮霉素的培养基上进行抗性筛选,经分化培养和生根培养,最终获得TG1靶点株系和TG2靶点株系,共16株CRISPR/Cas9突变体植株。利用潮霉素标记的特异引物Hyg-R和Hyg-F对其进行PCR鉴定,结果显示,这些植株均为阳性突变体植株(图4)。
2. 4 CRISPR/Cas9突变体植株OsC3H54基因碱基突变分析结果
采用CTAB法提取CRISPR/Cas9突变体植株DNA,以CRISPR/Cas9的引物(AT-F/AT-R)进行PCR扩增,PCR扩增产物测序结果显示,CRISPR/Cas9突变体植株在靶点序列的突变位点位置附近出现套峰(图5),表明2个株系的植株均发生突变。每个株系各挑2株突变体植株(Y1和Y2,Y3和Y4)用于分析OsC3H54基因碱基序突变情况,结果表明,Y1、Y2、Y3和Y4均为单碱基插入突变。
2. 5 CRISPR/Cas9突变体植株OsC3H54蛋白氨基酸突变分析结果
将4个突变体植株与野生型植株的OsC3H54蛋白氨基酸序列进行比对,结果发现4个突变体植株均出现移码突变,最终导致氨基酸翻译提前终止(图6)。以Y2为例,由于PAM附近插入单碱基,导致序列在441 bp处提前产生终止密码子,致使蛋白翻译提前终止于第147位氨基酸。
3 讨论
锌指蛋白是一个庞大的蛋白家族,不同锌指蛋白行使不同的功能,在植物抗病和非生物胁迫等方面具有重要作用。研究表明,锌指蛋白不仅可与RNA结合发挥作用,还能与其他蛋白质结合发挥功能。拟南芥中锌指蛋白可识别植株体内的RNA并与其结合调控植物的生长发育,其中HUA1是细胞核中的C3H型锌指RNA结合蛋白,在植株生长发育过程中参与调控花的发育(Cheng et al.,2003)。锌指蛋白也可与其他蛋白互作提高植株不同抗性,如棉花锌指蛋白GhZFP1可与GZIRD21A和GZIPR5结合,提高转基因烟草的耐盐性和抗真菌性(Guo et al.,2009)。锌指蛋白自身结构也能影响植物的生长发育,如锌指蛋白Ⅶ亚家族中的KH结构蛋白KHZ1和KHZ2能延缓拟南芥叶片衰老,调控植株延迟开花(Yan et al.,2017)。
目前,有关C3H基因的研究发现,水稻OsC3H54基因和OsC3H51基因可能是紧密同源的重复基因(Wang et al.,2008b);拟南芥C3H基因可对多种环境刺激响应,其中AtC3H54基因在冷胁迫时下调表达(Wang et al.,2008a);拟南芥中过表达AtC3H49/AtTZF3和AtC3H20/AtTZF2基因可提高植株对ABA的敏感性,减少叶片蒸腾,进而增强耐旱性,同时延缓茉莉酸途径介导的植株衰老过程(Lee et al.,2012)。C3H54作为锌指蛋白家族中的重要成员,但有关其功能研究鲜见报道。本研究通过CRISPR/Cas9基因编辑技术对OsC3H54基因进行基因编辑,获得其突变体植株,测序结果显示其主要为单碱基插入突变,其中A或T是主要的插入碱基类型,与Zhang等(2014)研究结果相似。水稻OsC3H54基因CRISPR/Cas9突变体植株的获得为进一步研究OsC3H54基因功能提供了良好材料。
4 結论
水稻OsC3H54基因CRISPR/Cas9突变体植株的获得为进一步研究水稻锌指蛋白的生物学功能提供了良好材料。
参考文献:
孔晓聪,邳瑞雪,石雨鹭,王荃,靳亚军,梁闪闪,张泗举,栾维江. 2019. 基于CRISPR/Cas9技术的水稻OsDUF1475突变体的创建与分析[J]. 农业生物技术学报,27(3):17-25. [Kong X C,Pi R X,Shi Y L,Wang Q,Jin Y J,Liang S S,Zhang S J,Luan W J. 2019. Obtainment and characteri-zation of rice(Oryza sativa) OsDUF1475 mutants based on CRISPR/Cas9 technique[J]. Journal of Agricultural Biotechnology,27(3):17-25.]
李贺,韩艺娟,林艺娟,刘丽华,张承康,张连虎,王宗华,鲁国东. 2015. 水稻锌指蛋白基因OsZFP1的功能分析[J]. 中国水稻科学,29(2):135-140. [Li H,Han Y J,Lin Y J,Liu L H,Zhang C K,Zhang L H,Wang Z H,Lu G D. 2015. Functional analysis of zinc finger protein gene OsZFP1 in rice[J]. China Rice Science,29(2):135-140.]
刘丹,李然红,陈鑫,王立凤. 2019. 狗枣猕猴桃AkSAP蛋白的生物信息学分析[J]. 河南农业科学,48(12):103-108. [Liu D,Li R H,Chen X,Wang L F. 2019. Bioinformatics analysis of AkSAP protein from Actinidia kolomikta[J]. Journal of Henan Agricultural Sciences,48(12):103-108.]
刘凯,严国红,张桂云,孙明法. 2019. 水稻滞绿突变分子遗传研究进展[J]. 江苏农业学报,35(2):484-488. [Liu K,Yan G H,Zhang G Y,Sun M F. 2019. Advances in molecular genetics of rice stay green mutation[J]. Jiangsu Journal of Agricultural Sciences,35(2):484-488.]
刘小艳,孙艳侠,王亚男,刘晓楠,刘坤,郗冬梅. 2015. 水稻CCCH型锌指蛋白亚家族Ⅰ基因的表达分析[J]. 山东农业科学,(2):7-11. [Liu X Y,Sun Y X,Wang Y N,Liu X N,Liu K,Xi D M. 2015. Expression analysis of CCCH-zinc finger protein subfamily Ⅰ genes in rice[J]. Shandong Agricultural Sciences,(2):7-11.]
Berg J M,Shi Y G. 1996. The galvanization of biology:A growing appreciation for the roles of zinc[J]. Science,271(5252):1081-1085.
Bogamuwa S,Jang J C. 2016. Plant tandem CCCH zinc finger proteins interact with ABA,drought,and stress response regulators in processing-bodies and stress granules[J]. PLoS One,11(3):e0151574.
Cheng Y L,Kato N,Wang W M,Li J J,Chen X M. 2003. Two RNA binding proteins,HEN4 and HUA1,act in the processing of AGAMOUS pre-mRNA in Arabidopsis tha-liana[J]. Developmental Cell,4(1):53-66.
Deng H Q,Liu H B,Li X H,Xiao J H,Wang S P. 2012. A CCCH-type zinc finger nucleic acid-binding protein quantitatively confers resistance against rice bacterial blight disease[J]. Plant Physiology,158(2):876-889.
Gao H,Zheng X M,Fei G L,Chen J,Jin M N,Ren Y L,Wu W X,Zhou K N,Sheng P K,Zhou F,Jiang L,Wang J,Zhang X,Guo X P,Wang J L,Cheng Z J,Wu C Y,Wang H Y,Wan J M. 2013. Ehd4 encodes a novel and oryza-genus-specific regulator of photoperiodic flowering in rice[J]. PLoS Genetics,9(2):e1003281.
Guo D,Yi H Y,Li H L,Liu C,Yang Z P,Peng S Q. 2015. Molecular characterization of HbCZF1,a Hevea brasiliensis CCCH-type zinc finger protein that regulates hmg1[J]. Plant Cell Reports,34(9):1569-1578.
Guo Y H,Yu Y P,Wang D,Wu C A,Yang G D,Huang J G,Zheng C C. 2009. GhZFP1,a novel CCCH-type zinc finger protein from cotton,enhances salt stress tolerance and fungal disease resistance in transgenic tobacco by interacting with GZIRD21A and GZIPR5[J]. New Phytologist,183(1):62-75.
Jan A,Maruyama K,Todaka D,Kidokoro S,Abo M,Yoshimura E,Shinozaki K,Nakashima K,Yamaguchi-Shinozaki K. 2013. OsTZF1,a CCCH-tandem zinc finger protein,confers delayed senescence and stress tolerance in rice by regulating stress-related genes[J]. Plant Physiology,161(3):1202-1216.
Kong Z S,Li M N,Yang W Q,Xu W Y,Xue Y B. 2006. A novel nuclear-localized CCCH-type zinc finger protein,OsDOS,is involved in delaying leaf senescence in rice[J]. Plant Physiology,141(4):1376-1388.
Lee S J,Jung H J,Kang H,Kim S Y. 2012. Arabidopsis zinc finger proteins AtC3H49/AtTZF3 and AtC3H20/AtTZF2 are involved in ABA and JA responses[J]. Plant and Cell Physiology,53(4):673-686.
Miller J,Mclachlan A D,Klug A. 1985. Repetitive zinc-bin-ding domains in the protein transcription factor IIIA from Xenopus oocytes[J]. The EMBO Journal,4(6):1609-1614.
Qiu A L,Lei Y F,Yang S,Wu J,Li J Z,Bao B J,Cai Y T,Wang S,Lin J H,Wang Y Z,Shen L,Cai J S,Guan D Y,He S L. 2018. CaC3H14 encoding a tandem CCCH zinc finger protein is directly targeted by CaWRKY40 and positively regulates the response of pepper to inoculation by Ralstonia solanacearum[J]. Molecular Plant Pathology,19(10):2221-2235.
Wang D,Guo Y H,Wu C A ,Yang G D,Li Y Y,Zheng C C. 2008a. Genome-wide analysis of CCCH zinc finger family in Arabidopsis and rice[J]. BMC Genomics,9(1):44.
Wang L,Xu Y Y,Zhang C,Ma Q B,Joo S H,Seong-Ki K,Xu Z H,Chong K. 2008b. OsLIC,a novel CCCH-type zinc finger protein with transcription activation,mediates rice architecture via brassinosteroids signaling[J]. PLoS One,3(10):e3521.
Yan Z Y,Jia J H,Yan X Y,Shi H Y,Han Y Z. 2017. Arabidopsis KHZ1 and KHZ2,two novel non-tandem CCCH zinc-finger and K-homolog domain proteins,have redundant roles in the regulation of flowering and senescence[J]. Plant Molecular Biology,95(6):1-17.
Zhang D M,Xu Z P,Cao S X,Chen K L,Li S C,Liu X L,Gao C X,Zhang B C,Zhou Y H. 2018a. An uncanonical CCCH-tandem zinc-finger protein represses secondary wall synthesis and controls mechanical strength in rice[J]. Molecular Plant,11(1):163-174.
Zhang H Q,Zhang Z Q,Xiong T,Xiong X H,Wu X M,Guan C Y,Xiao G. 2018b. The CCCH-type transcription factor BnZFP1 is a positive regulator to control oleic acid levels through the expression of diacylglycerol O-acyltransferase 1 gene in Brassica napus[J]. Plant Physiology and Biochemistry,132:633-640.
Zhang H,Gao X R,Zhi Y H,Li X,Zhang Q,Niu J B,Wang J,Zhai H,Zhao N,Li J G,Liu Q C,He S Z. 2019. A non-tandem CCCH-type zinc-finger protein,IbC3H18,functions as a nuclear transcriptional activator and enhances abiotic stress tolerance in sweet potato[J]. New Phytologist,223(4):1918-1936.
Zhang H,Zhang J S,Wei P L,Zhang B T,Gou F,Feng Z Y,Mao Y F,Yang L,Zhang H,Xu N F,Zhu J K. 2014. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation[J]. Plant Biotechnology Journal,12(6):797-807.
(責任编辑 陈 燕)
收稿日期:2020-01-20
基金项目:国家重点研发计划项目(2018YFD0301301-5-2);广西自然科学基金项目(2017GXNSFDA198039);广西水稻遗传育种重点实验室开放基金项目(160-380-16-1)
作者简介:*为通讯作者,沙爱华(1973-),博士,教授,主要从事植物分子生物学及作物育种研究工作,E-mail:aihuasha@163.com。易勇(1995-),研究方向为植物分子育种,E-mail:yy1111666 @163.com
猜你喜欢
世界中医药(2022年4期)2022-04-03
世界中医药(2021年22期)2021-01-03
热带作物学报(2020年7期)2020-09-02
中学生物学(2019年7期)2019-10-17
山东农业科学(2017年12期)2018-01-09
中学生物学(2017年7期)2017-08-23
科技资讯(2016年32期)2017-03-31
江苏农业科学(2015年11期)2016-01-27
热带农业科学(2014年8期)2014-09-23
