
铁皮石斛FCA基因的表达及生物信息学分析
2020-04-22 09:38李良建王万军
生物学杂志 2020年2期
李良建, 胡 睿, 裴 薇, 王万军
(西南交通大学 生命科学与工程学院, 成都 610031)
开花是判断植株成熟的一个重要指标,适时开花能保证花朵在适宜的条件下授粉和正常结果,以保证质量和产量[1]。目前发现植物成花调控存在6条途径:光周期途径(Photoperiod pathway)[2]、春化途径(Vernalization pathway)、赤霉素途径(GA pathway)[3]、自主途径(Autonomous pathway)[4]、年龄途径(Age pathway)[5]和温度途径[6]。这些调控途径相互独立又相互配合,形成有序而复杂的成花调控网络,共同控制植物的生殖发育。
自主途径是植物根据自身发育状态而控制开花,植物在其营养生长达到一定阶段后会自然开花,现在比较清楚的自主途径是通过抑制FLC基因的表达来促进开花[7-8]。参与自主途径的调控因子主要有FCA、FPA、FLOWERINGLOCUSY(FY)、LUMINIDEPENDENS(LD)、FLOWERINGLOCUSD(FLD)、FVE、FLK和REF6[9]。FCA是自主途径重要调控因子。其选择性剪接和选择性加尾,导致FCA基因产生4种不同转录物:α、β、γ及δ,其中只有FCA-γ编码完整有功能的FCA蛋白,含有2个RNA识别结合域(RNA-recognition motif, RRM)和1个WW蛋白相互作用结构域[10-11]。研究发现,FCA-γ在拟南芥(Arabidopsisthaliana)所有组织中都有表达,但在茎尖中的表达水平最高[12];过量表达分析表明,只有过量表达FCA-γ转录本时,可以明显促进植物提前开花[11]。FY是FCA WW结构域的伴侣蛋白,FCA-WW结构域与FYPPLPP基序相互作用而阻止FLCmRNA的积累,进而促进开花;此外,还能促使FCA前体mRNA在第3个内含子处发生多聚腺苷化反应,形成较短无活性的FCA蛋白,从而实现FCA的自我负调控[13-14]。近年还在拟南芥中发现一个新的RNA结合蛋白HLP1,能特异结合到FCA转录本的两个Poly (A) 位点附近,并促进远端Poly (A) 位点的聚腺苷酸化,从而产生有生物学功能的全长FCA转录本来抑制FLC的表达,促进开花[15]。
铁皮石斛是兰科,石斛属植物,具有较高的观赏价值和药用价值[16]。通常情况下,从种子萌发到开花至少需要3~13年,并且还具有明确的季节特性[17]。同时,春石斛的花芽分化需要低温诱导,无低温诱导,不能完成花芽分化[18]。目前在铁皮石斛的研究中还没有开花相关基因FCA的报道。本研究首次从铁皮石斛中克隆得到开花相关基因FCA,并利用RT-qPCR技术,检测该基因在铁皮石斛不同组织内的表达水平,探索其时空表达模式;同时研究低温对铁皮石斛FCA基因表达的影响,为探寻DoFCA调节开花的机制奠定基础。
1 材料与方法
1.1 材料
铁皮石斛种子接种于1/2MS培养基,于25 ℃恒温培养室培养获得试管苗,选择成熟一致且发育良好的铁皮石斛植株,分别收集根、茎、叶、花,提取其RNA,于-80℃保存。
选取相同条件下,具4片完全伸展叶片的植株进行胁迫处理。低温处理:将备用植株放入4 ℃的培养箱中,其他的培养条件均保持一致,分别选取处理1、6、12、24、48和72 h的植株提取RNA,于-80 ℃保存。
1.2 方法
1.2.1 铁皮石斛总RNA的提取及第一链cDNA的合成
使用Plant RNA Kit试剂盒(Omega)分别提取备用植株的根、茎、叶、花以及低温处理的整苗植株总RNA。使用SYNERGY H1全功能酶标仪检测总RNA含量,选择A260/A280为1.8~2.0,且凝胶电泳检测结果清晰的总RNA进行逆转录。逆转录反应采用TaKaRa公司的Reverse Transcriptase M-MLV (RNase H-)合成cDNA第一链。
1.2.2 铁皮石斛FCA基因cDNA序列的获取
对铁皮石斛原球茎转录组二代测序数据进行分析,获得5个FCA基因片段;以这5个片段为探针,用Blastn在铁皮石斛转录组数据库中进行延伸;用DNAMAN拼接后获得铁皮石斛FCA基因cDNA序列。根据拼接所得cDNA序列,用Primer Primer 5.0设计该基因的巢式PCR上游引物,下游引物采用3RP和3RNP(表1),进行该基因3′ RACE,验证DoFCA是否完整。
选25 μL体系,使用EasyTaqDNA聚合酶 (TRANSGEN BIOTECH) 进行巢氏PCR,包括:cDNA模板1 μL、上下游引物(10 μmol/L)各1 μL、EasyTaqbuffer 2.5 μL、EasyTaqDNA聚合酶 0.15 μL、ddH2O 19.35 μL;扩增条件如下:94 ℃预变性4 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸3 min,共35个循环;72 ℃再延伸5 min,最后4 ℃终止反应。PCR产物进行1.0% 琼脂糖凝胶电泳检测,挑选目的条带进行胶回收纯化,纯化产物连接到pGEM-T easy载体上,转入感受态E.coliDH5α,用蓝白斑筛选出阳性菌落,进行菌落PCR检测,无误后,由成都擎科梓熙生物技术有限公司测序。

表1 引物序列
1.2.3DoFCA的生物信息学分析
利用BioXM的ORF功能预测DoFCA基因开放阅读框和蛋白质翻译;利用UniProtKB数据库的BLAST在线分析工具进行基因鉴定;利用GSDS2.0在线软件和NCBI的CDD数据库分别分析DoFCA的基因结构和蛋白结构域;利用Protparam软件对DoFCA蛋白序列进行理化性质的分析[19];用SOPMA软件对DoFCA蛋白进行二级结构预测;在UniProtKB数据库中选取其他物种同源蛋白,利用DNAMAN (LynnonBiosoft)对其进行多序列比对分析;再选取更多同源蛋白用MEGA7构建系统进化树[20]。
1.2.4DoFCA在不同组织以及低温胁迫条件下的基因表达分析
用RT-qPCR检测不同组织及低温胁迫条件下DoFCA的mRNA表达情况。根据测序获得的cDNA序列,用Primer Primer5.0设计定量引物(表1),选铁皮石斛的管家基因DoActin1为内参基因(表1)。将各类cDNA按10 μL体系进行扩增:SYBR green 5 μL,cDNA (100 ng/μL) 0.5 μL,ddH2O 2.9 μL,上下游引物(5 μmol/L)各0.8 μL;扩增条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火10 s,72 ℃延伸25 s,共45个循环;72 ℃再延伸30 s,4 ℃终止反应。每个实验样品进行3次技术重复和3次生物学重复,并设立阴性对照实验。数据用LightCycler® 96 SW 1.1进行技术重复误差分析,所有技术重复误差均≤0.25,采用2-ΔΔCt方法进行相对表达量分析。
1.2.5 DoFCA-WW/FY-PPLPP的同源建模及优化
以DoFCA-WW结构域序列通过BLAST工具检索蛋白质结构数据库RCSB PDB,选用1YWI和1ZR7作为模板,序列一致性分别为64%和62%。使用Discovery Studio(DS) 3.0进行DoFCA-WW结构域和FY-PPLPP相互作用复合物的同源建模,获得WW/PPLPP复合物模型。
初始模型利用DS 3.0的Side-Chain优化、Loop优化,最后用分子动力学模拟对复合物进行整体优化,选择能量最低构象进行评价和分析。
分子动力学模拟步骤:1)Minimization Ⅰ。将模型溶解于生理盐水环境,采用最陡下降法进行10 000步能量最小化,收敛标准0.1 kJ/mol。2)Minimization Ⅱ。采用共轭梯度法进行10 000步能量最小化,收敛标准0.05 kJ/mol。3)Heating。系统从50 K进行40 ps加热到300 K。4)Equilibration。在300 K系统进行60 ps的平衡。5)Production。在1 atm,300 K条件下以NPT系统进行300 ps模拟。
2 结果与分析
2.1 DoFCA基因序列的获得以及序列分析
电子克隆后得到2689 bp cDNA序列,3′RACE克隆结果与其匹配,且包含18个A的poly(A)尾。将完整的cDNA比对到铁皮石斛基因组数据[16],结果显示与XM_020833856.1的一致性达到99.9%,且具有5端和3端UTR,说明拼接结果正确且完整。利用BioXM软件对该序列分析,发现开放阅读框长度为2301 bp,编码766个氨基酸残基。基因结构分析显示基因共有19个外显子,18个内含子;蛋白质结构域分析表明DoFCA蛋白在N-端(131~210、220~298)包含两个RRM结构域,C-端(605~634)存在一个WW结构域。进一步分析发现两个RRM结构域跨越了2-13外显子,14和15外显子则编码了WW结构域。
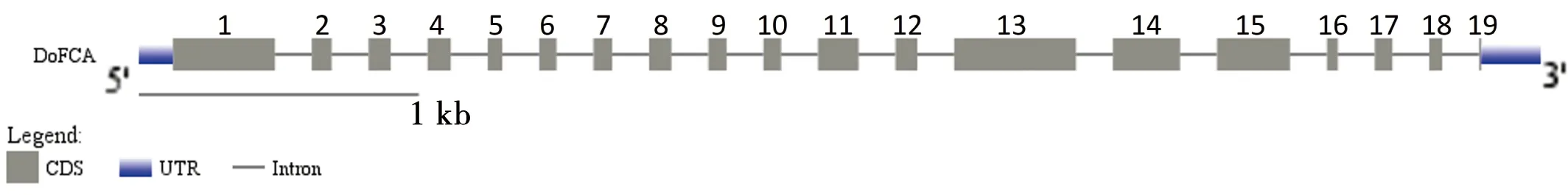
图1 DoFCA基因结构
从铁皮石斛基因组[16]中提取该基因TSS上游2000 bp启动子区,用PlantCARE进行调控元件分析。发现DoFCA启动子区存在WRKY、MYB和MYC 3种转录因子识别位点;热诱导顺式作用元件:STRE和AT-rich元件;激素应答元件:as-1和TGACG-motif;多种光响应元件。说明该基因可能对温度和激素有响应。此外,还包含CCGTCC-box、GCN4基序、‘CTCC’ 顺式作用元件3种元件,有研究证明这3个元件分别与植株的分生组织特异激活、胚乳表达、花药特异基因的表达调控密切相关[21-22]。反映该基因可能在铁皮石斛生殖发育中的重要地位。
2.2 DoFCA蛋白生物信息学分析
利用ExPASy Protparam在线软件对DoFCA所编码的蛋白质进行分析,该蛋白的分子量为82 920.34 ku,理论pI为8.40,原子总数为11 463,分子式为C3600H5628N1080O1130S25;不稳定系数Instability index为56.26,表明该蛋白质状态不稳定。带负电的残基总数(Asp + Glu)是54个,带正电的残基总数(Arg + Lys)为57个。亲水性平均值(GRAVY)为-0.602,为亲水性蛋白。
用SOPMA对DoFCA蛋白进行二级结构分析(图2),结果显示α螺旋(蓝色)占25.20%,延伸链(红色)占11.23%,β转角(绿色)占6.14%,无规则卷曲(紫色)占57.44%。无规则卷曲比重较大,说明该蛋白受侧链相互作用的影响较大。
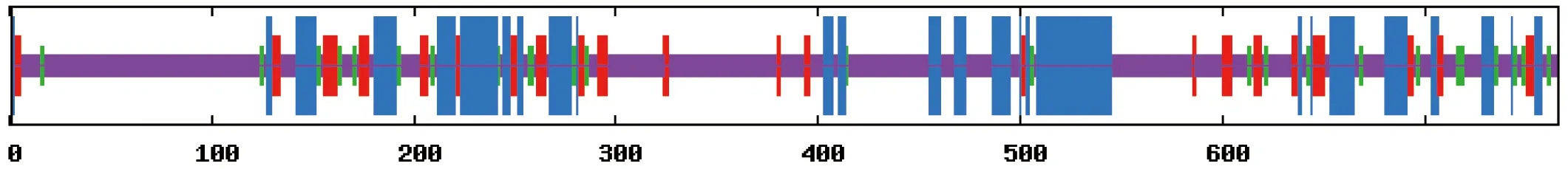
图2 DoFCA二级结构预测
Figure 2 Prediction of secondary structure of DoFCA

A0A2I0WFS5_9ASPA细茎石斛;A0A2I0ALZ0_9ASPA深圳拟兰; A0A2I0WRQ3_9ASPA细茎石斛; A0A2H3XUN6_PHODC海枣; A0A1U8AJT4_NELNU 莲
图3 不同物种FCA蛋白序列的多重比对
Figure 3 Multiple alignment of FCA protein sequences from different species
选择与DoFCA相似性较高的其他物种的蛋白序列,通过DNAMAN进行多序列比对(图3),一致性为68.71%(黑色表示完全一致,红色相似性较高,绿色相似性较低),比较发现保守区域主要集中在两个RRM结构域和WW结构域部分,表明该蛋白在不同物种中功能保守。
2.3 DoFCA系统发育分析
在UniProtKB数据库下载与DoFCA蛋白相似度较高的其他植物FCA蛋白,使用Clustalw进行多序列对齐,选择最大似然法进行系统发育树的构建(图4),氨基酸替换模型选择JTT Matrix-based model。
结果显示DoFCA蛋白与石斛类FCA蛋白的进化关系最近,并且单子叶植物FCA蛋白(蓝色)和双子叶植物FCA(红色)明显分为两支。说明FCA蛋白在单子叶和双子叶植物进化过程中出现了分化,与以往研究相符[13]。
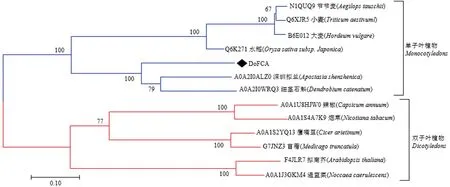
分支上的数字表示1000次重复搜索的自展百分比,标尺代表遗传距离
图4 不同物种的FCA蛋白序列系统发育树
Figure 4 Phylogenetic tree analysis of FCA protein sequences from different species
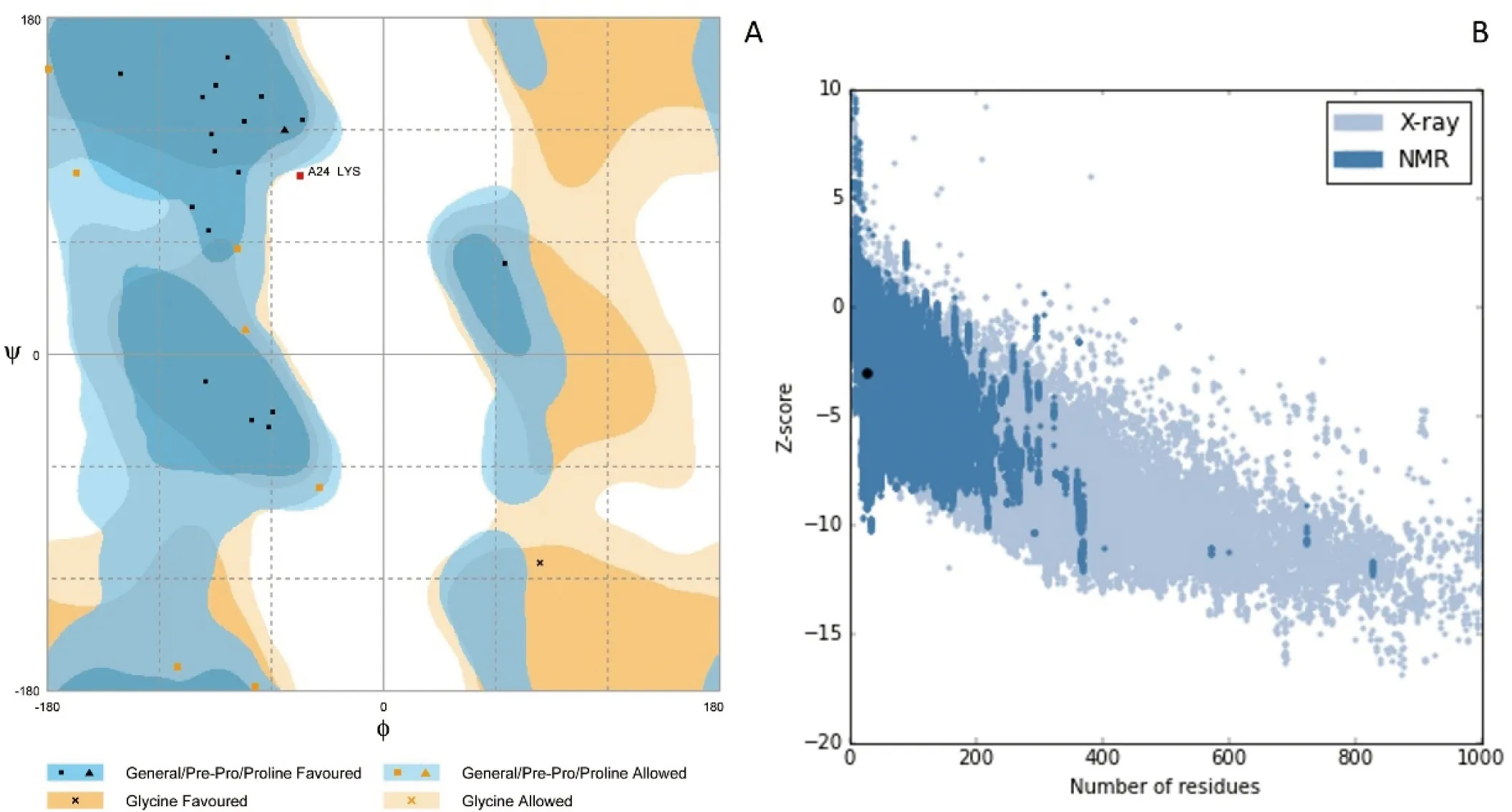
A:Ramachandran图;B:PROSA分析
图5 DoFCA-WW同源建模优化后评价结果
Figure 5 Evaluation results of DoFCA-WW homologous modeling optimization
2.4 DoFCA-WW/FY-PPLPP复合物结构及相互作用分析
同源建模和优化后,利用Ramachandran图(图5-A)检测模型立体化学的准确性,96.3%集中在最优区域和合理区域,仅3.7%分布在不合理区域,立体化学检测合格;PROSA分析(图5-B)反映蛋白质序列的长度和蛋白质所在的区域是否在晶体结构的合理区域,检测结果合格;Verify3D检测每个氨基酸残基侧链的合理性,DoFCA-WW每个残基平均分数均大于0.2,侧链结构合理。
图6-A为获得稳定的DoFCA-WW/FY-PPLPP复合物结构。通过对复合物结合模式的分析(图6-B),发现DoFCA-WW结构域中有8个相互作用结合位点:6His、10Glu、12Phe、14Tyr、16Tyr、22Glu、23Ser和25Trp。其中14Tyr、16Tyr、23Ser及25Trp 4个位点与FY-PPLPP相互作用最紧密,并且DoFCA-WW结构域的23Ser和FY-PPLPP基序的第2个Pro残基形成氢键。丙氨酸扫描8个结合位点也发现14Tyr、16Tyr、23Ser及25Trp的突变均会使复合物相互作用降低,说明这4个位置可能为关键结合位点。结合位点的饱和突变发现:6His变为Tyr、12Phe变为Trp、23Ser变为Thr或Val均能提高复合物稳定性;而25Trp的突变均会使复合物模型不稳定,反映了该残基的重要;同时14Tyr突变为苯丙氨酸外的氨基酸也会使复合物不稳定,可能为潜在关键位点。
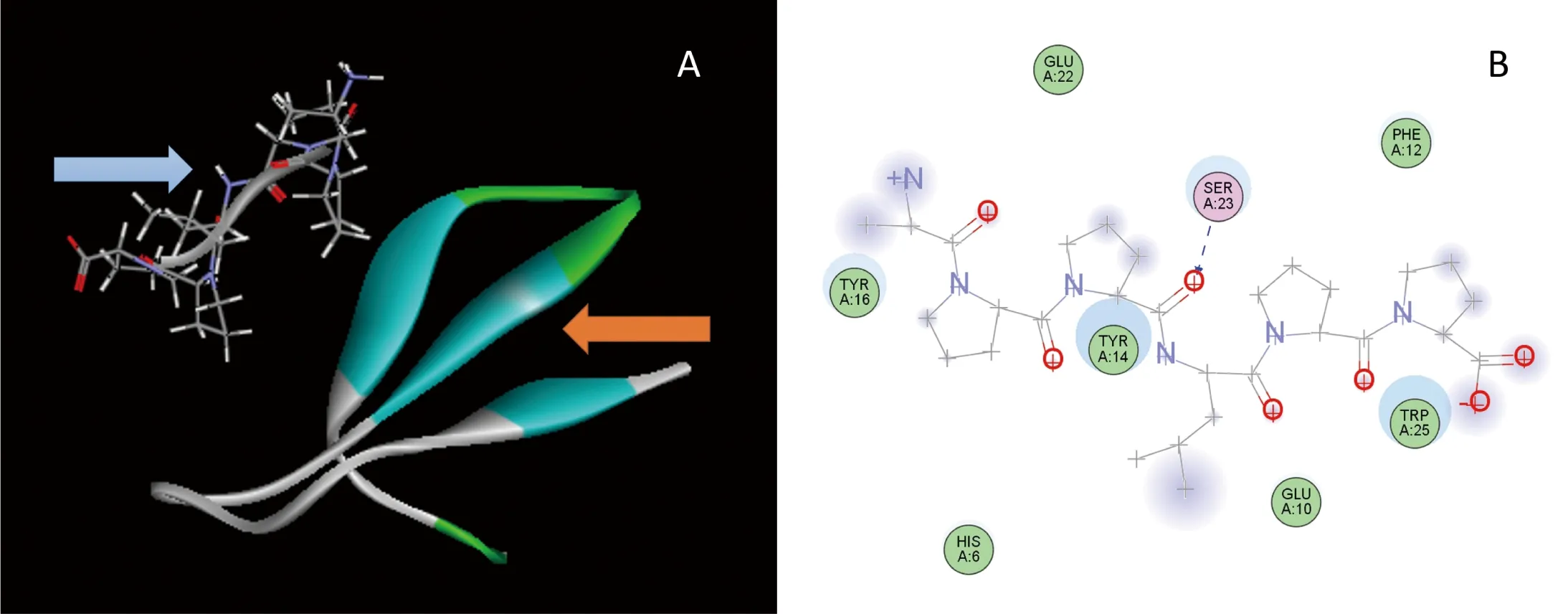
A:DoFCA-WW/FY-PPLPP复合物结构,FY-PPLPP配体(蓝色箭头),DoFCA-WW受体(橘色箭头);B:相互作用模式
图6 DoFCA-WW/FY-PPLPP复合物结构及相互作用模式
Figure 6 DoFCA-WW/FY-PPLPP complex structure and interaction pattern
2.5 DoFCA基因在不同组织以及低温胁迫条件下的基因表达分析
以DoActin1基因(表1)作为内参,进行RT-qPCR的分析,用2-ΔΔCt方法对原始Ct 值进行计算后得到DoFCA的相对表达量(不同组织以根的表达作为基准,低温胁迫以正常情况表达为基准),结果如图7所示。

A:不同组织DoFCA表达水平;B:低温条件下DoFCA表达水平
图7DoFCA在不同组织以及低温胁迫下的表达分析
Figure 7 Analysis ofDoFCAgene expression in different tissues and low temperature stress
DoFCA基因在不同组织中的表达结果显示,DoFCA在铁皮石斛花中表达量最高,而在根中的表达量最少,茎和叶中差别不大;表明DoFCA基因主要在花的形成和发育过程中发挥作用(图7-A)。低温胁迫的表达结果显示,初期DoFCA表达量逐渐减少,在处理到12 h时,表达量降到最低,之后表达量逐渐升高,高于正常值(图7-B)。
3 讨论与结论
目前铁皮石斛自主开花途径研究较少,FCA基因是自主开花途径中促进植物开花的保守基因,对其开展研究有助于更好地了解其调控机制。本研究获得了铁皮石斛FCA基因的完整开放阅读框,长度为2301 bp,编码766个氨基酸残基。基因结构分析显示其有19个外显子,18个内含子,这可能与该基因复杂的可变剪接有关。DoFCA蛋白的多序列比对显示,该蛋白在多个物种存在保守区域,保守区域主要集中在RRM结构域和WW结构域,说明该蛋白在多个物种中功能保守。系统发育分析指出在单子叶和双子叶植物中,FCA蛋白明显分为两支,说明FCA蛋白在单子叶和双子叶植物进化过程中出现了分化,与以往研究相符[13];同时,在单子叶植物FCA中,含有一个特异Glycine-Rich的结构域,在信号途径中调控G蛋白的功能,这可能是单子叶植物FCA功能多样性的原因之一[13]。
本研究发现DoFCA在铁皮石斛的表达存在组织特异性,在花中特异富集,其他组织中表达量较少。FCA基因在拟南芥几乎所有组织中表达,但在茎尖中的表达水平最高[12]。菜心BrcuFCA的强表达主要集中在花上[23]。对拟南芥FCA晚花突变体的研究也发现,FCA参与了植物营养生长时相转变的调控[24]。说明DoFCA对于花的发生和形成可能具有重要作用。
在温度对石斛兰花芽分化的研究中发现持续足够时间的低温是花芽分化的关键,低温诱导,能够促进花芽分化[25]。拟南芥开花研究发现,FCA可通过依赖FLC的春化途径或者不依赖FLC响应环境温度变化从而调控成花[14,26]。本研究发现DoFCA对低温胁迫有响应,且低温处理一段时间能促进DoFCA的积累。AI的研究也表明环境温度的变化可能改变PtFCA的表达模式[27]。说明DoFCA可能在低温诱导石斛兰花芽分化过程中发挥作用。
目前发现FCA至少有两个功能,即促进开花和负面调节自己的表达,这两个功能都需要完整的FCA-WW结构域与FY相互作用实现,并且WW结构域中的第二个色氨酸的突变会使其功能缺失[14]。对DoFCA-WW/FY-PPLPP相互作用模型进行分析,发现14Tyr、16Tyr、23Ser及25Trp 4个位点与FY-PPLPP相互作用最紧密。饱和突变发现25Trp的突变均会使复合物模型不稳定,反应了该残基的重要;同时14Tyr突变为苯丙氨酸外的氨基酸也会使复合物不稳定,可能为潜在关键位点,可进一步试验证明。
FCA除了与开花时间调控有关,目前还被证实与其他发育过程相关,比如种子休眠,种子千粒重和纤维产量等[28]。在拟南芥中FCA通过响应环境温度能促进miR172前体(pri-miR172),促进miR172积累,从而促进早花[12]。DoFCA启动子中也发现CCGTCC-box、GCN4基序、‘CTCC’ 顺式作用元件,被证实分别与植株的分生组织特异激活、胚乳表达、花药特异基因的表达调控密切相关[21-22]。反映FCA基因具有更多作用,有待进一步发掘。
本研究对铁皮石斛DoFCA基因进行了克隆,通过生物信息学分析和表达模式研究,发现该基因对铁皮石斛花的发生有重要的调控作用,为该基因在石斛中的功能研究奠定了基础,也为深入研究DoFCA提供了依据。
猜你喜欢
草堂(2022年4期)2022-11-11
贵州林业科技(2022年3期)2022-09-12
数学大王·中高年级(2020年12期)2020-12-31
趣味(语文)(2020年4期)2020-07-27
中成药(2018年7期)2018-08-04
中成药(2018年5期)2018-06-06
中成药(2018年3期)2018-05-07
现代园艺(2018年2期)2018-03-15
中成药(2017年5期)2017-06-13
安徽农业科学(2015年12期)2015-12-22
