
H9N2 禽流感病毒全基因组密码子使用偏好性及影响因素分析
2020-06-02 08:53黄蔚虹陈永杰孙彦阔孙卫东
华南农业大学学报 2020年3期
黄蔚虹,陈永杰,孙彦阔,孙卫东
(1 南京农业大学 动物医学院,江苏 南京 210095; 2 广东省动物源性人兽共患病预防与控制重点实验室/华南农业大学 兽医学院,广东 广州 510642)
H9N2 禽流感在1994 年首次暴发于我国广东省,引起蛋鸡产蛋率降低(14%~75%)和肉鸡死亡(10%~40%)[1]。在过去20 多年中,H9N2 禽流感病毒(Avian influenza virus,AIV)迅速蔓延传播,成为我国流行最为广泛的AIV 亚型[2]。H9N2 AIV 与细菌、病毒等病原体共感染时,可造成家禽死亡率大幅升高,对家禽养殖业构成严重威胁。同时,H9N2亚型禽流感已出现跨种传播的现象,广东、香港、湖南和安徽均出现人感染H9N2 AIV 的病例[2]。
同义密码子的使用具有某种倾向性,并非是一个随机的过程。在蛋白质翻译过程中,部分同义密码子会比其他密码子使用度更高,这被称为密码子使用偏好性(Codon usage bias)[3]。若病毒在同一宿主长时间循环,则该病毒的密码子使用模式将趋同于宿主的密码子使用模式(即病毒使用与宿主相同的同义密码子以翻译某一氨基酸),以便更好地利用宿主细胞资源进行病毒复制、繁殖[4]。研究密码子使用模式有助于解释病毒在基因进化过程中所发生的适应性变化,进一步认识病毒的基因表达调控,有助于病毒疫苗的研制[5-6]。H9N2 AIV 对家禽养殖和人类健康的威胁日益加剧,但目前对H9N2 AIV 密码子使用模式的研究较少。
为了探讨近年来H9N2 AIV 的密码子使用偏好性及其影响因素,本研究选取90 株2010—2018 年H9N2 AIV 的全基因组为研究对象,分析其碱基组成特性、最优密码子、密码子使用偏好性的影响因素以及病毒对宿主密码子使用模式的适应性,以期为H9N2 AIV 基因进化、已有疫苗密码子的优化和新型疫苗(密码子去优化疫苗)研制提供理论依据。
1 材料与方法
1.1 序列来源
利用NCBI 数据库(http://www.ncbi.nlm.nih.gov/genomes/FLU/FLU.html)检索获取2010—2018 年中国H9N2 AIV 全基因组(内含8 个节段的完整编码序列),根据遗传进化分析结果,从中选取90 个H9N2 AIV 流行毒株的全基因组。每株病毒的编码序列按PB2+PB1+PA+HA+NP+NA+M+NS 顺序进行拼接。
1.2 密码子使用偏好性参数分析
利用CodonW 1.4.4 软件计算H9N2 AIV 基因组的密码子使用偏好性参数,包括同义密码子第3 位各碱基(A3S、U3S、C3S和G3S)含量,同义密码子第3 位的GC(GC3S)含量,有效密码子数(Effective number of codons,ENC),总平均亲水性(General average hydropathicity)和芳香性(Aromaticity)。用Puigbò等[7]的方法计算各碱基(A、U、C 和G)含量,GC 含量,密码子第3 位各碱基(A3、U3、C3和G3)含量,密码子第3 位的GC(GC3)含量,密码子第1、2 位的GC(GC12)平均含量,同义密码子相对使用度(Relative synonymous codon usage)和密码子适应指数(Codon adaptation index)。宿主鸡Gallus gallus 和人Homo sapiens 的密码子使用模式获取自密码子使用数据库(http://www.kazusa.or.jp/codon/)[8]。
1.3 有效密码子数绘图分析
ENC 为20~60,与密码子使用偏好性呈负相关,ENC≤35 说明密码子使用偏好性高,ENC>50 说明密码子使用偏好性低[9]。ENC 绘图是以ENC 为纵坐标、GC3S含量为横坐标绘制的二维散点图,若基因组位于标准曲线(即ENC期望与GC3S含量的函数关系)上或其附近,表明密码子使用偏好性仅受突变压力影响;若基因组位于标准曲线下方较远的位置,表明密码子使用偏好性受其他因素(如自然选择)影响。ENC期望与GC3S含量的函数关系为:
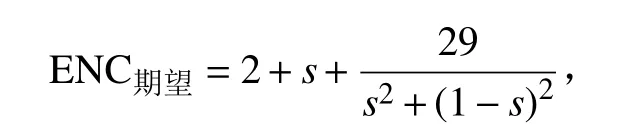
式中,s 为GC3S含量[9]。
1.4 中性绘图分析
采用中性绘图[w(GC3)为横坐标、w(GC12)为纵坐标]分析突变压力和自然选择对密码子使用偏好性的影响程度。若回归曲线斜率接近1,即基因组几乎沿对角线分布,表示密码子使用偏好性仅受突变压力影响;随着斜率逐渐减小,甚至减小为0,自然选择对密码子使用偏好性的作用逐渐增大[5]。
1.5 PR2 绘图分析
以w(A3)/w(A3+U3)为纵坐标、w(G3)/w(G3+C3)为横坐标绘制二维散点图,若w(A3)=w(U3) 且w(C3)=w(G3),基因组均匀分布于图中心(横、纵坐标均为0.5),表明突变压力和选择率(替代率)对密码子使用的作用相等;反之,两者作用存在差异[5]。
1.6 密码子适应指数分析
密码子适应指数为0~1,值越高,病毒对宿主的适应性越强[10]。密码子适应指数和ENC 之间的相关性分析可确定突变压力和自然选择对密码子使用偏好性的相对影响。若自然选择优于突变压力,则密码子适应指数和ENC 之间的相关性高,相关系数(r)趋向于-1;相反,若突变压力更为重要,则r 应接近0,密码子适应指数和ENC 之间无相关性[11]。
1.7 对应性分析
同义密码子相对使用度是其实际使用频率与期望频率(某一特定氨基酸的所有同义密码子能被均等使用的情况下该密码子的使用频率)的比率。同义密码子相对使用度等于1 代表对应氨基酸无密码子使用偏好性,大于1 代表对应氨基酸对该密码子的使用较多,小于1 代表对该密码子的使用较少。对应性分析(Correspondence analysis)是常用于研究密码子使用模式的多元统计分析方法,基于各基因组的同义密码子相对使用度,将每个基因组依密码子使用频率在1 个59 维的向量空间内表示[12]。
1.8 最优密码子分析
各基因组的ENC 从高到低依次排序,从上限区域和下限区域各取10%的基因组作为高、低表达组,使用t 检验分析两组之间密码子使用的差异显著性。比较两组的同义密码子相对使用度,若差异≥0.0 8,并且高表达组显著高于低表达组(P<0.01),则该密码子被定义为最佳密码子[9,13]。
1.9 统计学分析
使用CodonW 1.4.4 软件对同义密码子相对使用度进行对应性分析,通过SPSS 23.0 软件完成t 检验和Spearman 相关性分析,使用SigmaPlot 12.5 软件绘图。
2 结果与分析
2.1 碱基组成特性
由2010—2018 年H9N2 AIV 基因组的碱基组成分析结果(表1) 可知,各碱基含量为A(32.92%~33.03%)>G(24.65%~24.72%)>U(22.88%~22.90%)>C(19.41%~19.49%)。2010—2018 年GC 含量整体呈下降趋势,且均少于50%,提示H9N2 AIV 基因组中AU 含量多于GC 含量,碱基组成更倾向于使用AU,随着时间推移,该倾向性逐渐加大。同义密码子第3 位碱基含量为A3S(41.42%~41.80%)>U3S(30.76%~30.95%)>G3S(27.74%~28.11%)>C3S(26.51%~26.73%),GC3S含量变化与GC 相同,说明以A 或U 作为末位碱基的同义密码子的使用度较大,随着时间推移,其使用度将逐渐增大。
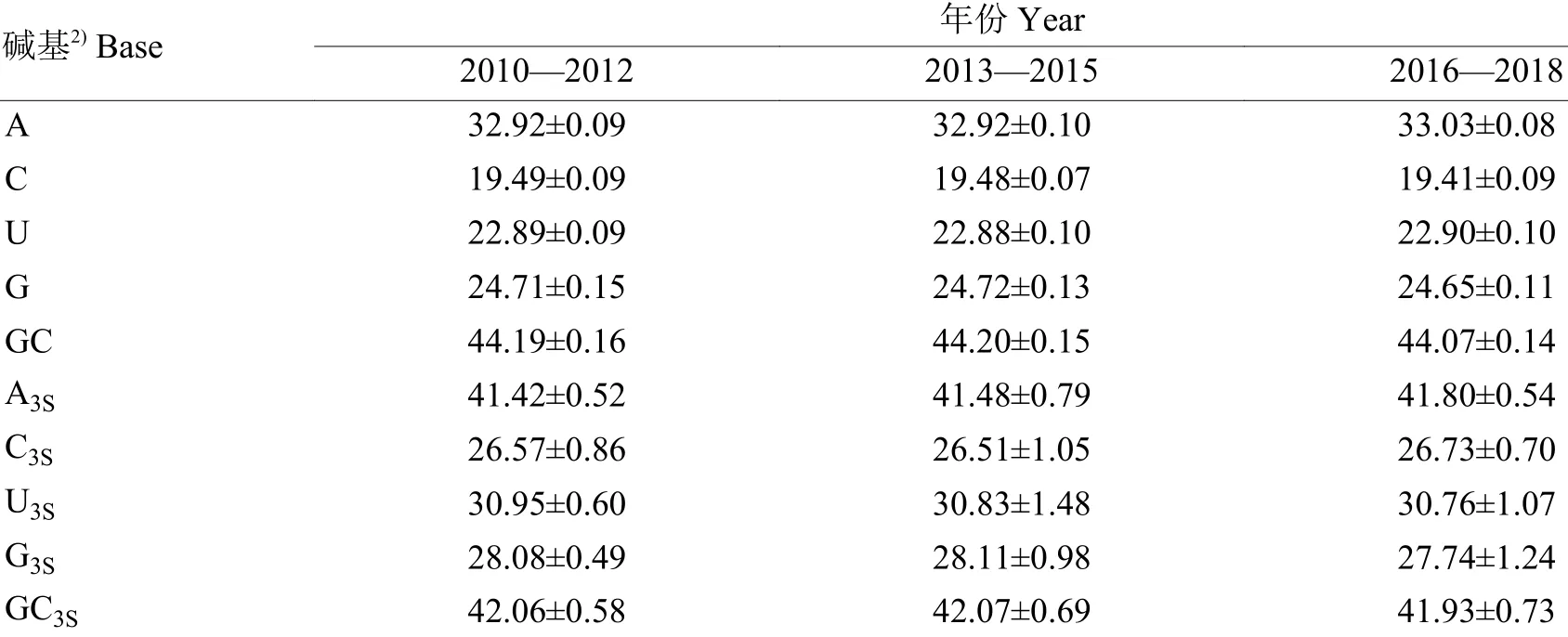
表 1 H9N2 禽流感病毒基因组的碱基组成分析1)Table 1 Base composition analysis of H9N2 avian influenza virus genomes %
2.2 最优密码子分析
以ENC 为依据形成的高、低表达组数据如表2所示,大部分同义密码子相对使用度差异较小且无显著性差异,仅有10 个密码子(UUU、GUU、GUA、GCU、GCC、GAU、CGU、CGA、GGU 和GGC)满足相对使用度差异≥0.08 且高表达组相对使用度显著高于低表达组(P<0.01)的条件,因此这10 个密码子可判定为最优密码子。其中有8 个最优密码子均以A 或U 结尾,仅有2 个以C 结尾,说明最优密码子的形成可能主要受到碱基组成限制,可能存在突变压力影响密码子使用偏好性。
2.3 突变压力和自然选择共同作用于密码子使用偏好性
2010—2018 年H9N2 AIV 的ENC 为51.90~55.01,平均值52.86,标准差0.64,说明H9N2 AIV 的密码子使用偏好性较低。在ENC 绘图(图1)分析中,所有点均聚集于标准曲线的下方,说明H9N2 AIV 密码子使用偏好性的主导因素并非突变压力,而是其他影响因素如自然选择。
PR2 绘图分析结果(图2)显示,2010—2018 年H9N2 AIV 基因组主要聚集于图中心(横、纵坐标均为0.5)的右上方,基因组对AU 的使用度大于GC,其中w(A3)>w(U3)且w(G3)>w(C3),这表明密码子使用偏好性可能由突变压力和其他因素(如自然选择)共同决定,但两者作用存在差异。
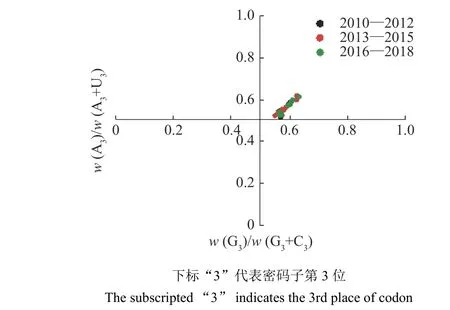
图 2 不同年份H9N2 禽流感病毒的PR2 绘图分析Fig. 2 PR2 plot analysis of H9N2 avian influenza virus in different years
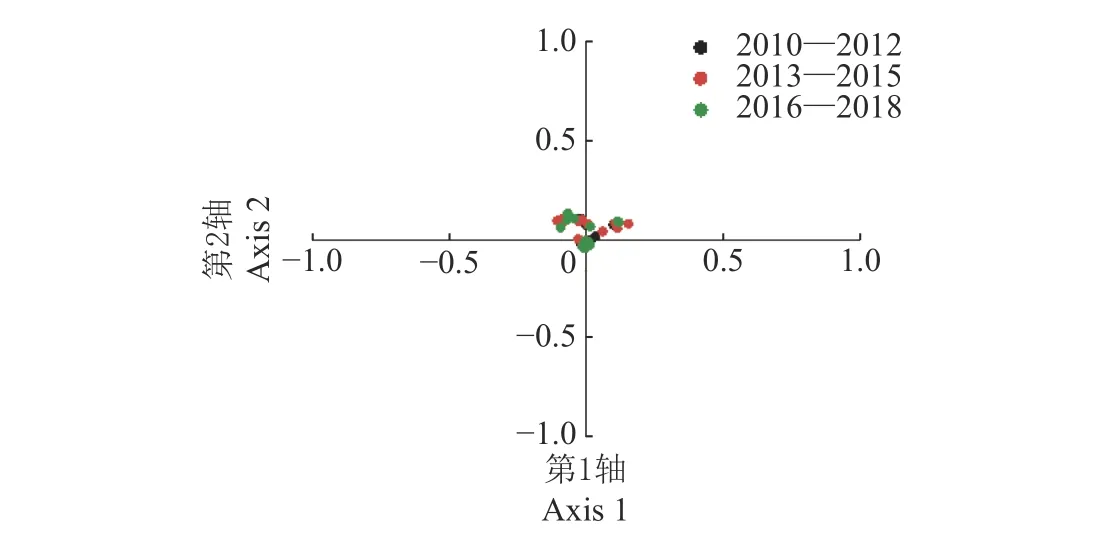
图 3 不同年份H9N2 禽流感病毒的对应性分析Fig. 3 Correspondence analysis of H9N2 avian influenza virus in different years
2.3.1 H9N2 禽流感病毒密码子使用偏好性受突变压力的影响 对2010—2018 年H9N2 AIV 进行对应性分析(图3),结果显示第1、2 轴分别解释34.65%和21.34%的变异,大部分基因组都聚集在原点(横、纵坐标均为0)周围,彼此之间不存在太大差异,这提示各基因组间的密码子使用偏好性差异相对较小。第1、2 轴与碱基组成指数进行相关性分析(表3),结果显示第1、2 轴与ENC 均呈极显著正相关(r=0.633、0.216,P<0.001);第1、2 轴与大部分碱基组成指数呈显著相关,其中第1 轴与GC 含量呈极显著正相关(r=0.510,P<0.001),第2 轴与G3S含量呈极显著负相关(r=-0.717,P<0.001)。这提示碱基组成对密码子使用偏好性产生影响。
2.3.2 H9N2 禽流感病毒密码子使用偏好性受自然选择的影响 第1、2 轴与总平均亲水性、芳香性进行相关性分析(表3),结果显示第1、2 轴与总平均亲水性、芳香性均存在显著相关性,这说明氨基酸的疏水性和芳香性均显著影响密码子使用偏好性。此外,对2010—2018 年H9N2 AIV 进行密码子适应指数分析(表4),结果显示H9N2 对宿主的密码子适应指数平均值随时间变化不大,但H9N2 AIV 对鸡的密码子适应指数平均值(0.766~0.768)均高于对人的(0.739~0.741)。同时,密码子适应指数与ENC 相关性分析发现,除了2013—2015 年H9N2 AIV 对人的密码子适应指数与ENC 相关性不显著(r=-0.223,P=0.237),其余密码子适应指数均与ENC 呈显著负相关,说明自然选择对H9N2 AIV 的密码子使用偏好性产生重要影响,但r≠-1,所以也存在突变压力产生影响。
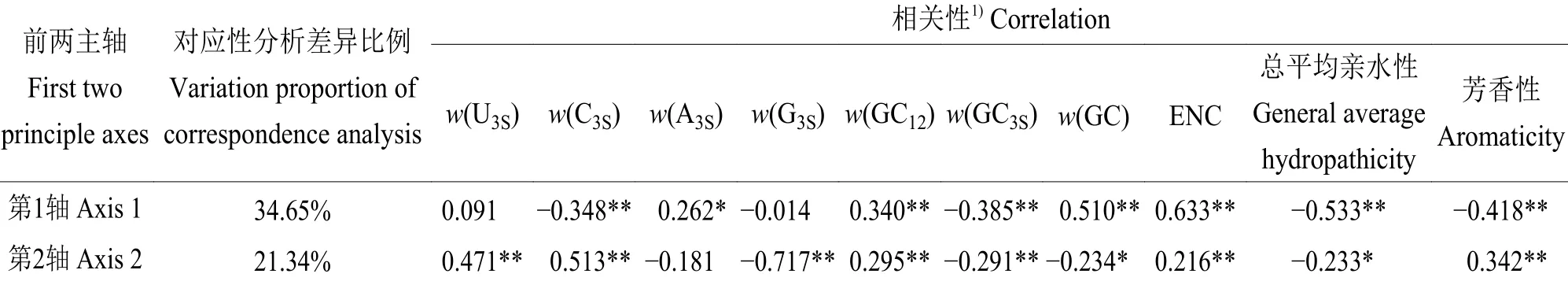
表 3 前两主轴对应性分析及与密码子使用偏好性主要参数的相关性分析Table 3 Correspondence analyses of the first two principle axes and correlation analyses between the first two principle axes and main parameters of codon usage bias

表 4 H9N2 禽流感病毒基因组的密码子适应指数分析1)Table 4 Codon adaptation index analysis of H9N2 avian influenza virus genomes
2.3.3 突变压力和自然选择对密码子使用偏好性的相对作用比例 由中性绘图分析和GC12含量与GC3含量相关性分析(图4)可知,2010—2012、2013—2015 和2016—2018 年的GC12含量与GC3含量均呈极显著负相关(r=-0.720、-0.826、-0.595,P<0.001),提示密码子的这3 个位置均受到突变压力的影响。对2010—2012、2013—2015 和2016—2018 年的GC12含量与GC3含量进行线性回归分析发现,斜率分别为-0.238 5、-0.382 1 和-0.346 4,表明突变压力对H9N2 AIV 密码子使用偏好性的影响分别为23.85%(2010—2012 年)、38.21%(2013—2015 年) 和34.64%(2016—2018 年),所以自然选择对密码子使用偏好性的影响较大,分别为76.15%(2010—2012 年)、61.79%(2013—2015 年) 和65.36%(2016—2018 年)。

图 4 不同年份H9N2 禽流感病毒的中性绘图分析Fig. 4 Neutrality plot analysis of H9N2 avian influenza virus in different years
3 讨论与结论
H9N2 AIV 全基因组的碱基组成特性和最优密码子结果显示,基因组中AU 含量较多且偏好使用以A 或U 为末位碱基的同义密码子,这提示密码子使用偏好性可能对H9N2 AIV 的进化产生影响。ENC 与密码子使用偏好性呈负相关,密码子使用偏好性大小可通过ENC 值来进行评估,本研究发现H9N2 AIV 的ENC 平均值为52.86,说明不同序列间的基因组组成保守且相对稳定,H9N2 AIV 的密码子使用偏好性较低,这与其他A 型流感病毒和一些RNA 病毒相似,如H3N8(ENC=52.09)[14]、H5N1(ENC=50.91)[15]、H1N1pdm(ENC=52.5)[16]、H3N2(ENC=53.05)[3]、寨卡病毒(ENC=53.93)[17]和马尔堡病毒(ENC=54.2)[5]。低密码子使用偏好性病毒与宿主的合成竞争较低,可在与其密码子使用模式不相同的宿主细胞内进行有效的复制,从而在宿主体内维持其存活周期[5,16]。密码子使用偏好性可能会受到突变压力、自然选择、基因长度、GC 含量、基因表达水平和RNA 结构等因素的影响,其中突变压力和自然选择为主要影响因素[18-19]。
为了确定突变压力对密码子使用偏好性的作用,对应性分析的第1、2 轴与碱基组成指数的相关性分析结果显示第1、2 轴与大部分碱基组成指数呈显著性相关,提示碱基组成对密码子使用偏好性产生影响,即突变压力为H9N2 AIV 密码子使用偏好性的主要影响因素之一。但当病毒试图适应宿主细胞时,密码子使用偏好性往往会受到自然选择而降低[19]。对应性分析的第1、2 轴与总平均亲水性、芳香性之间均呈现显著相关性,这说明氨基酸的疏水性和芳香性均显著影响着密码子使用偏好性,自然选择作用于H9N2 AIV 的密码子使用偏好性。PR2 绘图分析发现基因组对AU 的使用度大于GC,揭示H9N2 AIV 密码子使用偏好性可能由突变压力和其他因素(如自然选择)共同决定,但两者作用不相同。综合对应性分析、PR2 绘图和相关性分析结果,可得出结论:突变压力和自然选择为主要影响因素,共同作用于H9N2 AIV 密码子使用偏好性;两者作用的相对大小仍为未知。
为了衡量突变压力和自然选择对密码子使用偏好性作用的相对大小,ENC 绘图分析发现所有基因组均聚集于标准曲线的下方,提示突变压力并非是H9N2 AIV 密码子使用偏好性的主导因素,而是其他影响因素(如自然选择)起主导作用。但有报道[11]指出突变压力和自然选择可能影响ENC,使其不能有效评估突变压力和自然选择对密码子使用偏好性的相对作用;此外,基因组为AU 偏向的物种,其密码子使用偏好性将与基因组为GC 偏向的存在差异,因此ENC 绘图分析可能具有误导性。
密码子适应指数是研究自然选择影响密码子使用偏好性的不二选择,可用于评估病毒对宿主的适应性,所以利用ENC 和密码子适应指数之间的相关性来研究突变压力和自然选择对密码子使用偏好性的影响[20]。H9N2 AIV 对鸡和人的密码子适应指数平均值偏高,提示H9N2 AIV 已适应鸡和人的密码子使用模式,能够更好地利用宿主细胞资源进行病毒复制和繁殖,有利于病毒在宿主内存活和进化;而且H9N2 AIV 可能会发展为对人类有致病性的病毒,已有多例H9N2 AIV 感染人类事件发生[2,21],随着H9N2 AIV 逐渐适应人类密码子使用模式,对人类的致病性将逐渐增强,因此需要密切监测密码子适应指数变化。同时,密码子适应指数与ENC 相关性分析发现大部分密码子适应指数与ENC 均呈显著负相关且r≠-1,这说明自然选择对密码子使用偏好性的作用大于突变压力,但突变压力的作用也不能忽视。
进行中性绘图分析以揭示两者作用比例,从GC12含量与GC3含量的线性回归分析发现,突变压力对H9N2 禽流感病毒密码子使用偏好性的作用分别为23.85%(2010—2012 年)、38.21%(2013—2015 年)和34.64%(2016—2018 年),而自然选择对密码子使用偏好性的作用较大,分别为76.15%(2010—2012年)、61.79%(2013—2015 年) 和65.36%(2016—2018 年)。中性绘图分析、ENC 绘图分析、密码子适应指数及其相关性结果揭示,在H9N2 AIV 密码子使用偏好性中,自然选择为主导作用(所占比例为61.79%~76.15%),而突变压力为次要作用(所占比例为23.85%~38.21%)。与H9N2 同为A 型流感病毒的H3N2[3]和H3N8[22]的密码子使用偏好性也是以自然选择为主要影响因素。
综上所述,H9N2 AIV 全基因组为AU 偏向且最优密码子多以A 或U 结尾,而且H9N2 AIV 基因组的密码子使用偏好性较低,主要受到突变压力和自然选择的共同作用,其中自然选择为主导作用(所占比例为61.79%~76.15%),而突变压力为次要作用(所占比例为23.85%~38.21%)。H9N2 AIV 已适应人类的密码子使用模式,可能对人类有致病性且致病性在逐渐增强。
猜你喜欢
福建农业学报(2021年6期)2021-08-18
考试与评价·七年级版(2021年2期)2021-08-14
科学(2020年3期)2020-11-26
教学考试(高考生物)(2020年4期)2020-11-18
初中生学习指导·中考版(2020年7期)2020-09-10
当代水产(2020年3期)2020-06-15
发明与创新·中学生(2019年6期)2019-06-26
安徽农业科学(2018年1期)2018-05-14
小星星·阅读100分(高年级)(2015年11期)2015-11-28
医学研究杂志(2015年12期)2015-06-10
