
马尾松毛虫肠道细菌的分离鉴定与产蛋白酶细菌的筛选
2020-08-30 12:38江宇航李宏伟蔡赛波林连兵张棋麟
浙江农业学报 2020年8期
江宇航,李宏伟,蔡赛波,林连兵,张棋麟,*
(1.昆明理工大学 生命科学与技术学院,云南 昆明 650500; 2.云南省高校饲用抗生素替代技术工程研究中心,云南 昆明 650500)
蛋白酶是一类催化蛋白质分解为蛋白胨、多肽和游离氨基酸的重要水解酶[1]。目前,已广泛应用于食品、造纸、清洁剂、饲料添加剂、医药、酿造、化妆品等多个领域,是最重要的工业用酶之一[2-3]。蛋白酶种类繁多,来源广泛,按其来源可大致分为植物、动物和微生物蛋白酶3大类[4]。目前在工业上广泛应用的蛋白酶主要是植物蛋白酶和动物蛋白酶,而这2类蛋白酶主要来源于动物内脏、植物果实与茎叶[5],易受到自然因素、环境变化和产酶源自身生理特性等多种因素的影响,使其具有分离提纯复杂,不易获得、产量低下等劣势[6-7]。随着现代化工业对蛋白酶需求量的逐渐增大,寻找到具有高活性、酶学性质稳定、产量高、成本低廉等优势的蛋白酶来源变得刻不容缓[8]。人们通过对微生物次级代谢产物的不断挖掘和认识,发现部分微生物在代谢过程中能产生多种蛋白酶,且大量产蛋白酶菌株存在于独特的环境中(如高海拔、重金属暴露和加工食品等)。例如,在青藏高原采集的土壤中获得了耐低温(0~40 ℃能保持较高酶活性)皮氏类芽孢杆菌(Paenibacilluspeoriae)产蛋白酶菌株[3],废铬革屑堆积土壤中的耐Cr3+蜡样芽孢杆菌(Bacilluscereus)产蛋白酶菌株[9],以及从某些谷类豆制品中分离得到的具有拮抗营养因子的枯草芽孢杆菌(Bacillussubtilis)产蛋白酶菌株[10]。此外,细菌产生的蛋白酶还具有类型丰富、稳定性好、易大量生产等特点[3,9-11]。
昆虫作为自然界中对环境适应能力最强、分布最广、肠道微生物资源较为丰富的动物类群,其肠道内的细菌具有较高的多样性和独特性,显示出多种生物学功能[12-13]。研究表明,昆虫肠道细菌能分泌多种水解酶协助昆虫进行食物消化、营养吸收,并降解利用淀粉、脂肪、纤维素、蛋白质等大分子能量物质[14]。其中,蛋白酶是昆虫分解食物中蛋白质的一类关键水解酶,大多为胞外蛋白酶,与植物蛋白酶相比,具有来源广、菌株选育方便、菌体易培养、产量高、酶学性质稳定等诸多优点[15]。目前,已从昆虫肠道中筛选出一批酶活性能优良的产蛋白酶菌株。例如,Zhou等[16]从黑胸散白蚁(Reticulitermeschinensis)肠道中筛选获得的霍氏肠杆菌(Enterbacterhormaechei)和地衣芽孢杆菌(Bacilluslicheniformis)展现了高效的木质素降解能力,且在55 ℃时仍保持较高活性;Wang等[17]从家蚕(Bombyxmori)中分离获得多株不动杆菌属(Acinetobacter)产碱性蛋白酶菌株,其具有良好的酸碱耐受性。随着对昆虫肠道菌群资源进行深入挖掘,筛选具有酶活性高和酶学特性优良的蛋白酶菌株是一项具有应用前景的工作。
马尾松毛虫(Dendrolimuspunctatus)隶属鳞翅目、枯叶蛾科、松毛虫属,是我国历史性的林业害虫之一,广泛分布于四川、安徽、陕西、海南、云南等多个省份[18]。目前,国内外对松毛虫的研究主要集中在将其作为有害生物进行防治,对其肠道微生物资源的研究与利用较少。研究表明,棉铃虫(Helicoverpaarmigera)[19]、烟夜蛾(Helicoverpaarmyworm)[20]、大豆夜蛾(Anticarsiagemmatalis)[21]等多种鳞翅目害虫幼虫肠道中存在芽孢杆菌(Bacillus)、假单胞菌(Pseudomonas)、肠杆菌(Enterobacter)等多个种属的产蛋白酶细菌。马尾松毛虫也是鳞翅目昆虫,本研究目标是通过传统微生物分离纯化技术,对马尾松毛虫幼虫肠道细菌进行体外分离培养研究,以期从中筛选获得高产蛋白酶细菌,并对多个蛋白酶活性较强的菌种进行基本酶活特性(热稳定性和酸碱耐受性)分析。
1 材料与方法
1.1 马尾松毛虫幼虫肠道细菌的分离与纯化
供试虫源于2019年8月采自贵州省天柱县,4~5龄健康幼虫,通过cox1分子标记进一步确认种名。取36头龄期一致的健康幼虫,试验前饥饿处理24 h,置于超净工作台上。用无菌水冲洗幼虫体表3次,以去除异物,用75%乙醇均匀清洗幼虫表面,再用0.1%的HgCl2清洗幼虫表面30 s进行消毒,最后用无菌水对幼虫体表冲洗3次。于解剖镜(奥林巴斯,日本)下迅速取出其肠道,装入含有500 μL无菌水的1.5 mL无菌EP管中,研磨棒研磨匀浆,加无菌水至1 mL。迅速用无菌水按10倍梯度稀释至10-6,取10-1、10-3、10-63个梯度菌液各100 μL,用涂布棒均匀涂布于牛肉膏蛋白胨培养基(NA)、牛肉膏蛋白胨葡萄糖培养基(BPDA)、LB培养基和哥伦比亚血琼脂培养基(CA)上,于37 ℃恒温培养箱中培养24 h。试验独立重复3次。长出菌落后,利用无菌接种环挑取单菌落,经4次平板划线最终得到纯培养菌株。最后,将纯化得到的单菌落进行编号并置于装液量为25 mL的无菌玻璃试管中,于37 ℃、150 r·min-1恒温摇床培养12 h,添加50%的甘油置于-80 ℃超低温冰箱中保存。
1.2 细菌DNA的提取与菌种鉴定
将已编号的马尾松毛虫肠道细菌按照细菌基因组DNA提取试剂盒(天根,中国)的操作步骤进行DNA提取。以提取获得的细菌基因组DNA为模版,选择27F和1492R 2种通用引物进行PCR,反应体系25 μL:MIX 12.5 μL,DNA模板1 μL,27F、1492R引物各0.5 μL,其余体系用无菌水补齐。PCR扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸50 s,30个循环;72 ℃延伸10 min。利用1.5%的琼脂糖凝胶进行电泳,并送至生工生物工程(上海)股份有限公司进行切胶纯化与双向测序直至测通。将测序结果利用NCBI的BLAST在线服务器(https://blast.ncbi.nlm.nih.gov/Blast.cgi)与细菌数据库进行比对分析,选取得分最高、序列一致性在99%及以上的已知细菌种类确定目标物种。
1.3 产蛋白酶菌株筛选
利用酪蛋白选择培养基,配方为:酵母粉2.5 g、蛋白胨5.0 g、琼脂20.0 g、酪蛋白10.0 g、KH2PO40.3 g、NaCl 1.0 g、MgSO40.5 g、无菌双蒸水1 L。将鉴定到的单菌落接种于酪蛋白选择培养基上,于37 ℃恒温培养24 h,加入适量的三氯乙酸处理10 min,观察有无水解圈,并利用游标卡尺准确测量记录。对初筛获得的产蛋白酶细菌进行3次独立重复试验。产酶量以水解圈直径(D)与菌落直径(d)之比(D/d)表示,具有酪蛋白水解圈的菌株确定为产蛋白酶细菌,进行后续试验。
1.4 细菌蛋白粗酶液制备
挑取具有产蛋白酶能力的细菌单菌落,37 ℃、150 r·min-1恒温振荡培养12 h,按5%的接种量接种于装有酪蛋白液体选择培养基的250 mL玻璃锥形瓶中,于37 ℃、150 r·min-1恒温振荡培养24 h。取发酵液置于装液量为50 mL的无菌离心管中,10 000×g离心5 min,收集上清液,弃去菌体沉淀,得到蛋白粗酶液,密封放置于4 ℃冰箱中保存备用。
1.5 酶活性测定
将各细菌产生的蛋白粗酶液按照GB/T 23527—2009《蛋白酶制剂》标准检测方法进行酶活性测定:将酪蛋白溶液pH调节至7.0,吸取蛋白粗酶液和底物(酪蛋白溶液)各l mL,振荡混匀,于40 ℃恒温水浴锅中反应10 min,加入2 mL 0.4 mol·L-1的三氯乙酸终止反应,再10 000×g离心5 min,静置10 min,弃去菌体沉淀;取上清液1 mL加入新的无菌玻璃管中,再加入5 mL 0.4 mol·L-1的Na2CO3溶液和1 mL的Folin-酚试剂,振荡混匀,置于40 ℃恒温水浴锅中保温发色20 min,利用722N可见分光光度计(上海仪电,中国)测定660 nm的吸光度。以添加三氯乙酸终止起始反应的一组为对照组,每组设置3个重复。酶活性单位:将每min水解酪蛋白产生1 μg酪氨酸所需的酶量定义为1个酶活性单位(U)。
1.6 蛋白酶热稳定性和酸碱耐受性分析
选取蛋白酶活性最高的3个菌种(如同一菌种包含不同酶活性的菌株,则选取酶活性最高的菌株作为该菌种代表,开展后续试验),将各菌株单菌落接种于装有酪蛋白液体选择培养基的200 mL锥形瓶中,于37 ℃、150 r·min-1恒温摇床培养24 h,获得蛋白粗酶液,按以下处理进行粗酶液热稳定性和酸碱耐受性处理:(1)分别在30、35、40、45、50、55、60 ℃恒温水浴处理10 min,自然冷却至室温,最高酶活性定义为100%,其余酶活性以所占最高酶活性的百分比表示;(2)分别调节粗酶液pH至4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0,于37 ℃温育1 h,pH调回7.0,最高酶活性定义为100%,其余酶活性以所占最高酶活性的百分比表示。进行单因素方差分析和多重比较,每组设置3个重复。
1.7 数据分析
产酶量和蛋白酶活性均以平均值±标准差表示。利用Microsoft Excel 2007对所测得的数据进行整理分析与绘制图表,用IBM SPSS 22.0软件对所测数据进行显著性检验。采用one-way ANOVA的单因素方差分析和最小显著差异法(least-significant difference, LSD)测验不同处理的蛋白酶活性与差异显著性;采用Student’s t-test检验细菌菌落直径与水解圈直径的差异显著水平。
2 结果与分析
2.1 肠道细菌的分离
从LB、NA、BPDA和CA培养基上培养获得马尾松毛虫幼虫肠道细菌96株,经16S rDNA分子鉴定发现,这96个菌株为18个不同种属的细菌,其中LB培养基14种,NA培养基6种,BPDA培养基7种,CA培养基5种(表1)。18种细菌分别隶属于厚壁菌门(Firmicutes)中的芽孢杆菌属,变形菌门(Proteobacteria)中的亚硫酸杆菌属(Sulfurobacillus)、葡萄球菌属(Staphylococcus)、嗜冷杆菌属(Psychrobacter)、泛菌属(Pantoea)、假单胞菌属、不动杆菌属(Acinetobacter)和柠檬酸杆菌属(Citrobacter)。
2.2 产蛋白酶细菌的筛选与酶活性
在酪蛋白选择培养基上获得6株具有清晰水解圈的菌株,其菌落直径显著(P<0.05)小于水解圈直径,将其作为产蛋白酶细菌(表2)。6个菌株均隶属于芽孢杆菌属,分别为解淀粉芽孢杆菌(Bacillusamyloliquefaciens)、枯草芽孢杆菌(Bacillussubtilis)、地衣芽孢杆菌(Bacilluslicheniformis)。产酶量和酶活性测定结果显示:地衣芽孢杆菌(MW-39)的蛋白酶活性最佳,其产酶量和蛋白酶活性分别为6.95±0.49和(255.36±1.32)U·mL-1;其次为解淀粉芽孢杆菌(MW-1和MW-50),其产酶量和酶活性最高为6.45±0.83和(193.85±1.86)U·mL-1;枯草芽孢杆菌(MW-4、MW-6、MW-32)菌株产酶量和酶活性最高分别达6.25±0.68和(165.81±1.23)U·mL-1。
2.3 蛋白酶的热稳定性与酸碱耐受性
地衣芽孢杆菌(MW-39)、解淀粉芽孢杆菌(MW-50)和枯草芽孢杆菌(MW-4)的蛋白粗酶液经不同温度处理后,在30~60 ℃蛋白粗酶液均保持较高活性,50 ℃时MW-39产生的蛋白酶活性为86.69%,MW-4产生的蛋白酶活性为74.65%,MW-50产生的蛋白酶活性为69.98%。经60 ℃处理后MW-39、MW-4和MW-50产生的蛋白酶活分别下降至51.94%、42.63%和36.26%。且MW-39产生的蛋白粗酶液在45 ℃时,酶活性达到最高,为(295.36±2.63)U·mL-1(100%),而MW-4和MW-50产生的蛋白粗酶液在40 ℃时,酶活性分别达到最高,为(196.25±1.85)U·mL-1和(223.68±1.69)U·mL-1(100%)(图1-a)。经过单因素方差分析与多重比较发现,MW-39、MW-4和MW-50产生的蛋白酶活性在不同的温度间差异有统计学意义(P<0.05)(表3—5)。3个菌株产生的粗蛋白酶在pH 4.0~11.0均保持一定的蛋白酶活性,在pH 4.0时,MW-39产的蛋白酶活性下降至58.37%,MW-4下降至41.89%,MW-50下降至49.65%;在pH 11.0时,MW-39、MW-4和MW-50产生的蛋白酶活性分别下降至36.56%、21.58%和28.76%。
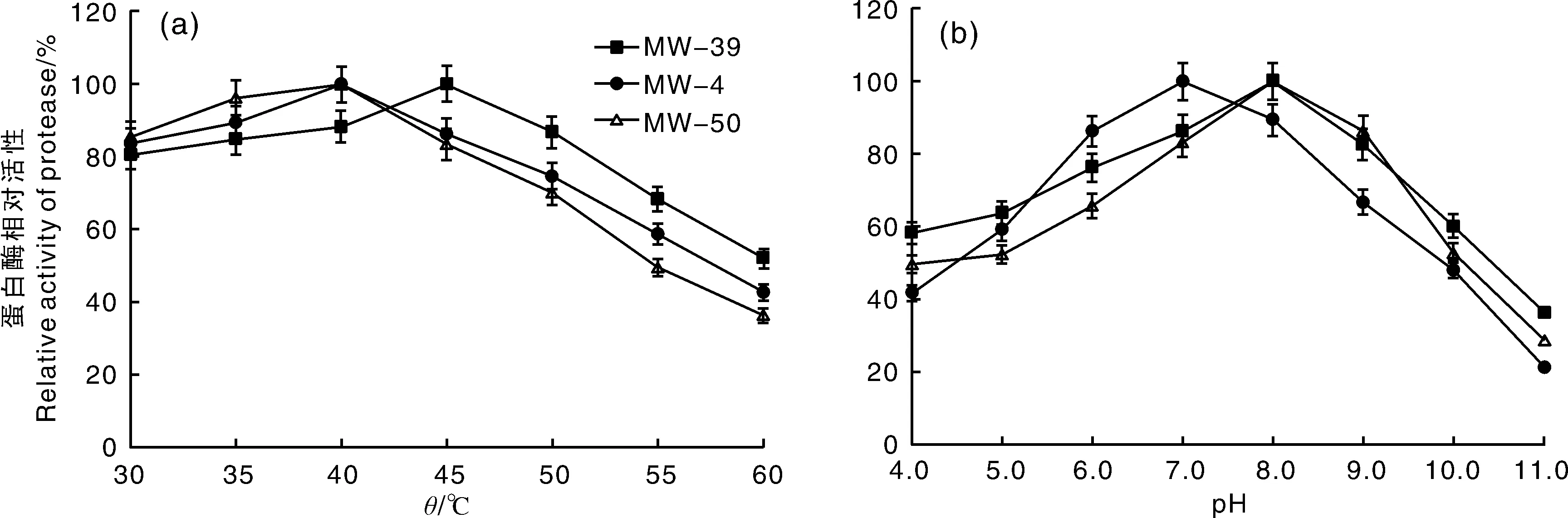
MW-39,地衣芽孢杆菌;MW-4,枯草芽孢杆菌;MW-50,解淀粉芽孢杆菌。MW-39, Bacillus licheniformis; MW-4, Bacillus subtilis; MW-50, Bacillus amyloliquefaciens.图1 马尾松毛虫肠道3种细菌产生的粗蛋白酶在不同温度(a)和pH(b)的相对活性Fig.1 Relative activity of crude protease from three species bacteria in intestine of Dendrolimus punctatus larvae under different temperatures (a) and pH (b)
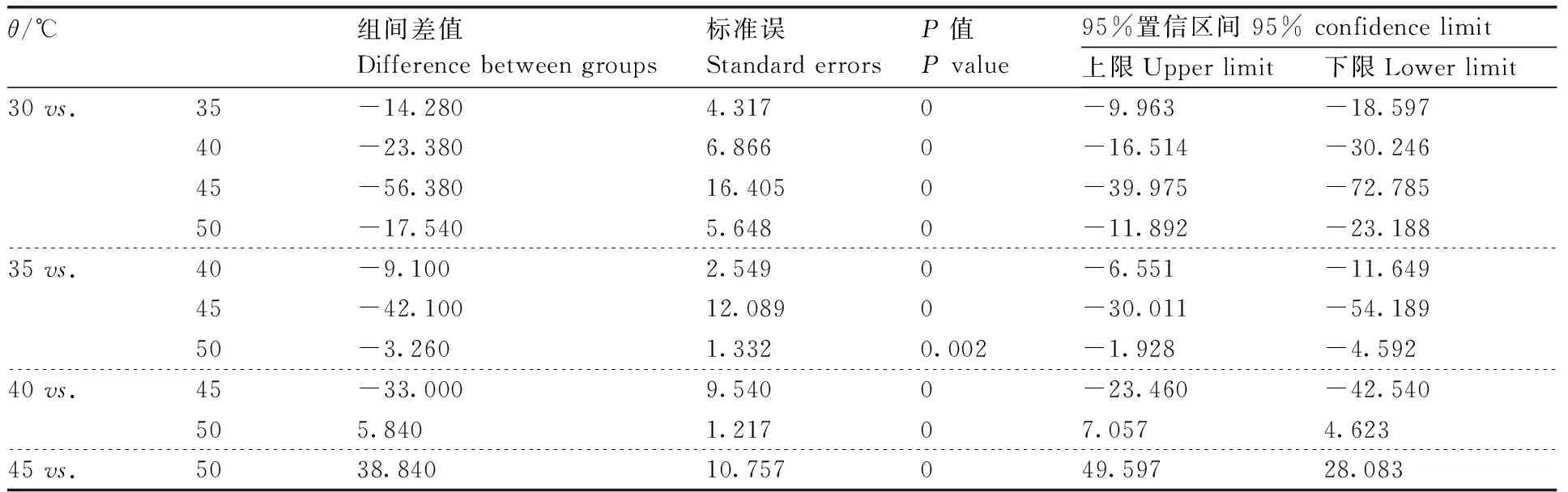
表3 不同温度下地衣芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
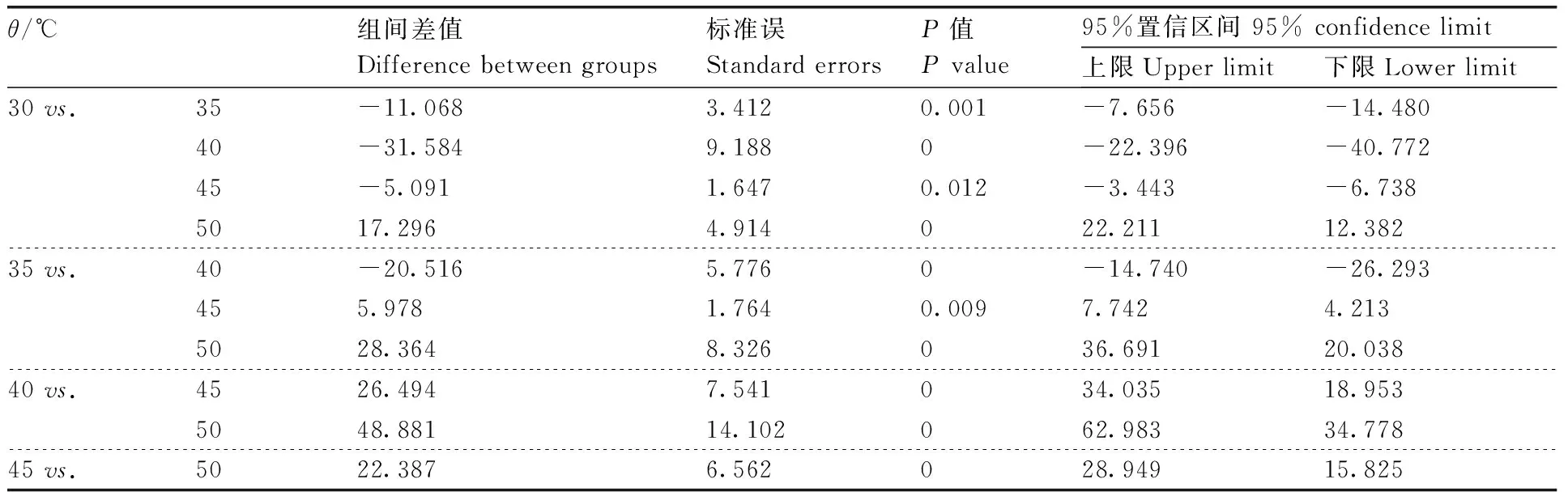
表4 不同温度下枯草芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
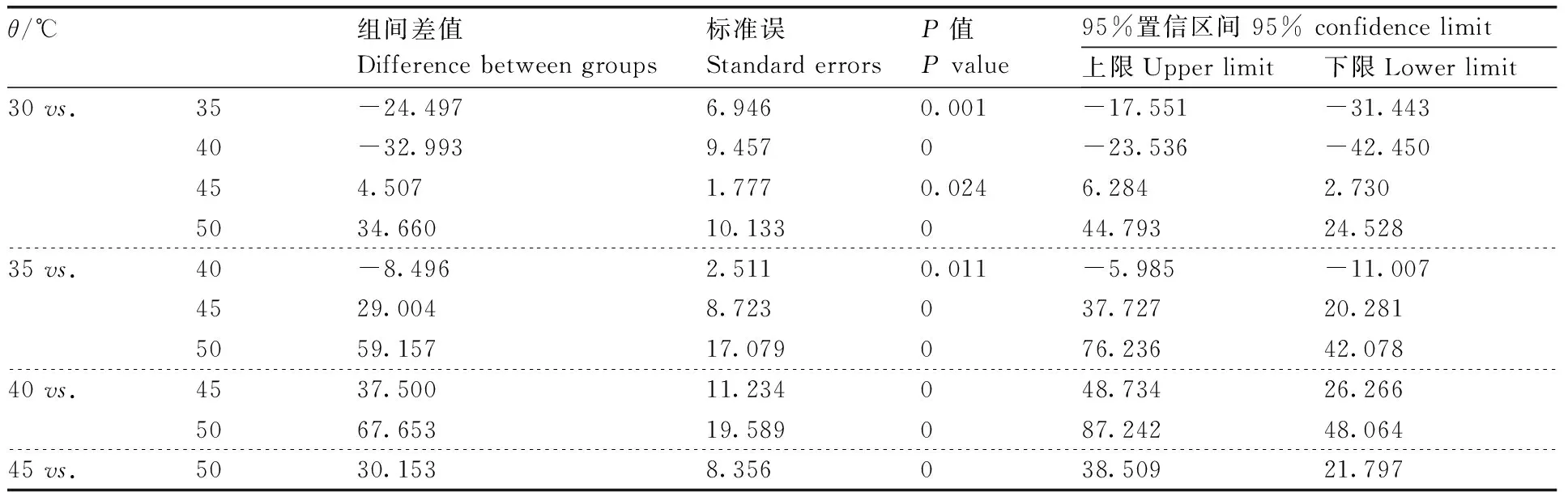
表5 不同温度下解淀粉芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
同时,MW-39和MW-50产生的蛋白粗酶液在pH 8.0时酶活性最高,分别为(285.65±1.93)U·mL-1和(218.39±2.39)U·mL-1(100%),而MW-4产生的蛋白粗酶液在pH 7.0时酶活性达到最高,为(183.46±1.23)U·mL-1(图1-b)。通过对pH 5.0~9.0的蛋白酶活性进行单因素方差分析与多重比较发现,MW-39、MW-4和MW-50产生的蛋白酶活性在不同的pH下差异有统计学意义(P<0.05)(表6,7和8)。
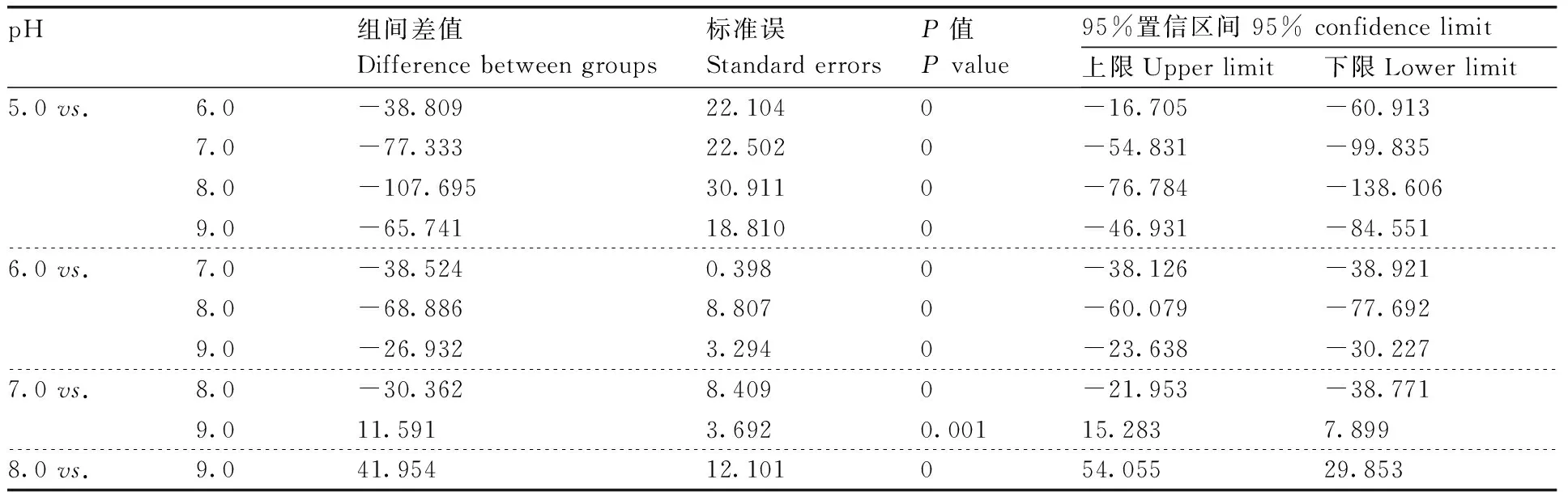
表6 不同pH下地衣芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
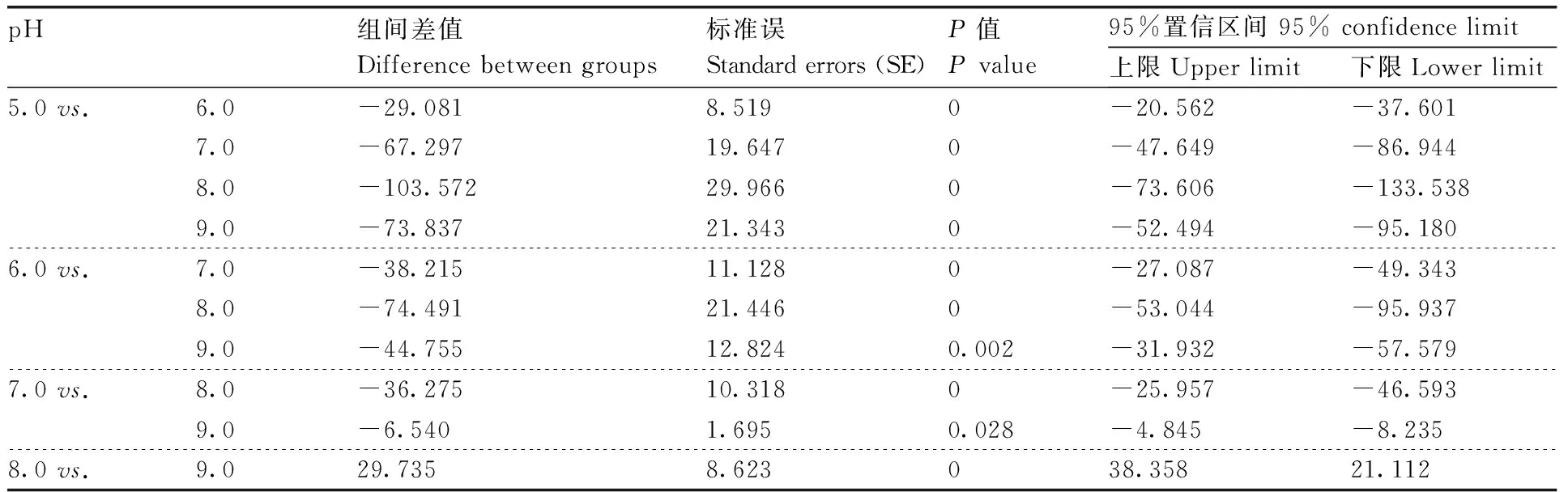
表7 不同pH下枯草芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
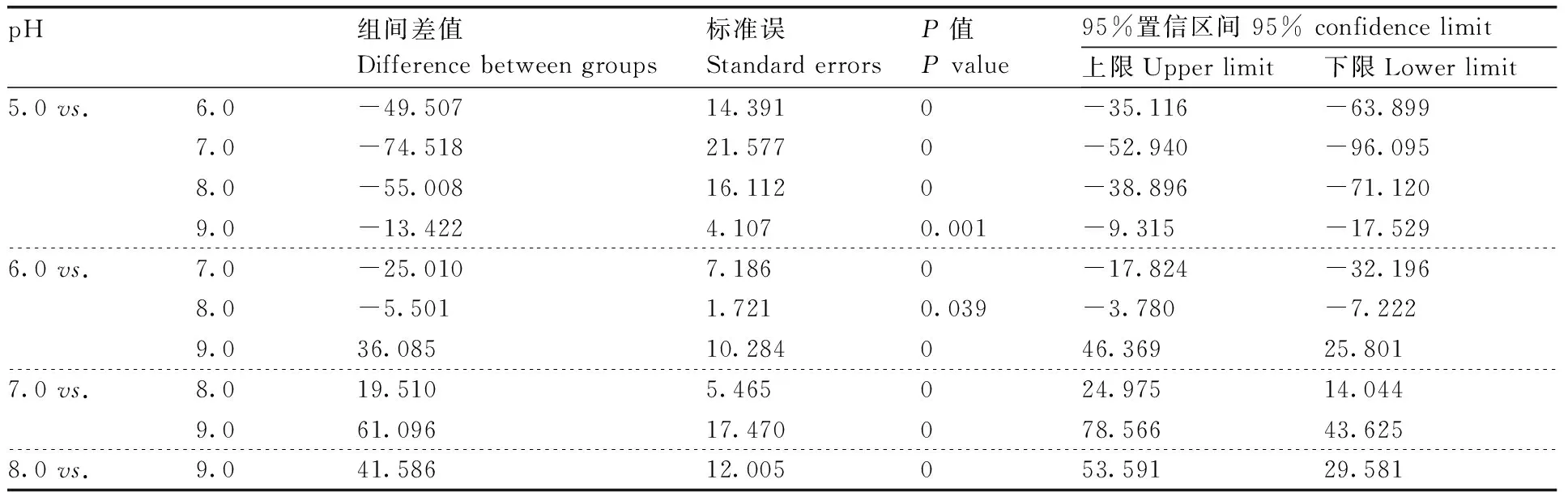
表8 不同pH下解淀粉芽孢杆菌蛋白酶活性的单因素方差分析与多重比较
3 讨论
本研究共分离获得2门18种马尾松毛虫幼虫肠道细菌,包括变形菌门和厚壁菌门,这与其他动物类群肠道细菌组成具有一定相似性,特别是在科和属的分类单元水平[22-23]。以前的研究对云南松毛虫(Dendrolimushoui)幼虫[24]肠道菌群分离结果显示,肠杆菌科、芽孢杆菌属和假单胞菌属细菌是其肠道中占比较大的优势菌群。对草原毛虫(Gynaephoraaureate)幼虫[25]肠道细菌的分离发现,芽孢杆菌属和葡萄球菌属细菌是其肠道的优势菌群,在这些鳞翅目昆虫肠道中还分离到了嗜冷杆菌属和亚硫酸杆菌属细菌。此外,本研究在马尾松毛虫幼虫肠道内鉴定到的柠檬酸杆菌属和泛菌属等非肠道优势菌群在其他昆虫肠道菌群中也有报道。例如,林胜等[26]从褐飞虱(Nilaparvatalugens)若虫肠道中分离获得泛菌属的成团泛菌(Pantoeaagglomerans),黄振东等[27]从德国小蠊(Blattellagermanica)肠道中分离获得多株柠檬酸杆菌属细菌。可见,不同昆虫肠道内存在部分相同细菌,这些细菌类群可能在昆虫祖先时期就已定殖于肠道,为较原始的昆虫肠道菌,这为我们了解昆虫肠道菌群演化提供了信息。然而,由于地理环境、生存条件、取食方式等多种因素的影响,不同类群的昆虫塑造了各自独特的肠道菌种类。值得注意的是,由于分离培养法无法准确模拟出昆虫肠道菌的生长环境,大量的肠道细菌种类还不能进行宿主体外培养,可能有部分极具价值的产细菌素细菌资源被忽视。随着基于高通量测序的肠道微生物多样性和宏基因组测序技术的发展,人们正在从海量基因组学数据中探索细菌的生物学特性,从而开发出匹配的培养基。如2015年,有研究者在《Nature Communications》上发表了根据16S rDNA序列直接对目标菌株进行培养基预测的数据库(http://komodo.modelseed.org/default.htm)[28],为挖掘和利用肠道微生物资源提供了新的思路。此外,由于培养条件的限制,尚未对马尾松毛虫肠道中厌氧细菌进行筛选。本研究所鉴定到的菌株大多为好氧细菌,其次为兼性厌氧细菌(如柠檬酸杆菌)。肠道是厌氧环境,厌氧菌的数量较为庞大,故挖掘肠道产蛋白酶厌氧菌菌株是一项有意义的工作。
芽孢杆菌是产蛋白酶的常见细菌属,它们广泛存在于昆虫肠道和自然环境中,能分解蛋白质等大分子物质[29]。以往研究表明,芽孢杆菌中的许多细菌都具有产蛋白酶能力,主要包括解淀粉芽孢杆菌、枯草芽孢杆菌、地衣芽孢杆菌、苏云金芽孢杆菌(Bacillusthuringiensis)和蜡状芽孢杆菌(Bacilluscereus)等[30]。本研究中蛋白酶活性最高的3种细菌均为芽孢杆菌,且其种类和以往研究报道的具有产蛋白酶能力的芽孢杆菌一致,表明芽孢杆菌在马尾松毛虫幼虫肠道蛋白质分解中起关键作用,昆虫肠道中存在大量产蛋白酶的芽孢杆菌,尤其地衣芽孢杆菌、解淀粉芽孢杆菌和枯草芽孢杆菌。另外,本研究在葡萄球菌和肠杆菌等细菌中,未发现产蛋白酶菌株,这与丰硕等[31]从小菌虫(Alphitobiuslaevigatus)肠道中只获得产蛋白酶芽孢杆菌的结果类似。邹昌瑞等[32]在柞蚕(Anthereapernyi)幼虫肠道中筛选获得的葡萄球菌和肠杆菌也不产蛋白酶。也有研究表明,肠杆菌属的许多细菌能介导昆虫对芽孢杆菌的敏感性,并能帮助昆虫降解蛋白质。因此,一些肠道细菌(如肠杆菌等)可能介导了马尾松毛虫肠道对食物中蛋白质的分解。此外,部分产蛋白酶细菌种属显示了在不同昆虫类群肠道间的特异分布。例如,本研究的马尾松毛虫肠道中产蛋白酶细菌为芽孢杆菌,云南松毛虫肠道中产蛋白酶细菌为霍氏肠杆菌(Enterobacterhormaechei)[33],德国小蠊肠道中产蛋白酶细菌为Chryseobacteriummassiliae和粘质沙雷菌(Serratiamarcescens)[27],家蚕肠道中产蛋白酶细菌为不动杆菌属(Acinetobacter)[17]。这些证据表明,昆虫肠道中产蛋白酶的细菌种类具有较高的多样性和一定的种属特异性。
酪蛋白选择培养基和蛋白酶活性测定结果显示,马尾松毛虫幼虫肠道的多株芽孢杆菌能产生高活性的蛋白酶,尤其以地衣芽孢杆菌蛋白酶活性表现最好,是Nadeem等[34]和刘海进等[35]发现的地衣芽孢杆菌产生的蛋白酶活性的1.5~2.5倍;解淀粉芽孢杆菌蛋白酶活性次之,与宁豫昌等[36]发现的解淀粉芽孢杆菌产生的高产蛋白酶活性相似;枯草芽孢杆菌蛋白酶活性相较前两者低,但是王东等[37]报道的枯草芽孢杆菌蛋白酶性的1.2~1.5倍。马尾松毛虫肠道细菌产生的蛋白酶在pH 5.0~9.0具有相对较高的蛋白酶活性。其中,地衣芽孢杆菌和解淀粉芽孢杆菌产生的蛋白酶在pH 8.0时活性达到最高,为碱性蛋白酶;枯草芽孢杆菌产生的蛋白酶在pH 7.0时活性最高,为中性蛋白酶。3种蛋白酶均能耐受pH 4.0和11.0的酸碱度,表明不同芽孢杆菌来源的蛋白酶最适酸碱条件不同。在生产应用中,可根据实际酸碱环境选择不同菌种来源的芽孢杆菌蛋白酶。此外,供试的3种芽孢杆菌蛋白酶活性在60 ℃仍然保持35%以上,尤其地衣芽孢杆菌蛋白酶活性仍高达50%以上,表明地衣芽孢杆菌、枯草芽孢杆菌、解淀粉芽孢杆菌所产生的蛋白酶具有较好的热稳定性。3种菌产生的蛋白酶活性均在40 ℃左右达到最高,这一结果与湖泊中发现的地衣芽孢杆菌[38]、豆粕中发现的枯草杆菌[10],以及猪肠道内发现的解淀粉芽孢杆菌[36]的适宜蛋白酶活性温度相似,表明不同生态环境和动物肠道来源的芽孢杆菌具有相似的蛋白酶热力学性质。单因素方差分析发现,供试芽孢杆菌蛋白酶在偏碱性条件下的活性显著高于偏酸性条件,与以往报道的地衣芽孢杆菌[39]、枯草芽孢杆菌[37]和解淀粉芽孢杆菌[36]蛋白酶为中性或碱性酶的研究结果一致。以上结果表明,马尾松毛虫幼虫肠道中的芽孢杆菌来源蛋白酶具有较好的热稳定性和酸碱耐受性,在工业化制备酶制剂方面显示了巨大的应用潜力。因此,未来细菌蛋白酶基因工程改良、优质菌株选育和蛋白酶大规模发酵制备等,应优先关注地衣芽孢杆菌、枯草芽孢杆菌和解淀粉芽孢杆菌等芽孢杆菌属细菌,尤其是能产高活性且具有较强耐热和耐酸碱性能蛋白酶的地衣芽孢杆菌。
本研究通过传统体外培养方法对马尾松毛虫肠道细菌进行研究,分离鉴定了一批不同种类的肠道细菌,以芽孢杆菌为主,其中地衣芽孢杆菌的蛋白酶活性最佳,枯草芽孢杆菌和解淀粉芽孢杆菌次之,明确了多株/种细菌所产蛋白酶的适宜温度和pH。值得注意的是,本研究所测试的蛋白酶学性质仅为温度和酸碱耐受性等基本指标。根据实际生产和试验的需要,还应关注更多与蛋白酶应用目标密切相关的酶学特性(如对重金属、有机溶剂和农药等耐受能力)。另外,肠道产蛋白酶细菌帮助昆虫消化食物,本研究发现的高产蛋白酶细菌作为马尾松毛虫肠道益生菌,可考虑将其作为防控马尾松毛虫灾害的靶标。例如:针对产蛋白酶菌株,筛选其特异的噬菌体、克隆制备噬菌体裂解酶,开发低毒特异化学杀菌剂等,对马尾松毛虫为害林区进行规律喷洒,破坏其肠道产蛋白酶细菌功能,使马尾松毛虫失去或减弱消化能力,为马尾松毛虫防治提供新的思路。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
疯狂英语·新读写(2022年1期)2022-11-23
中国饲料(2022年5期)2022-04-26
新农业(2021年12期)2021-11-29
南方农业·中旬(2021年5期)2021-10-25
农产品加工(2020年17期)2020-10-22
食品安全导刊·中旬刊(2020年2期)2020-06-01
绿色科技(2019年5期)2019-11-29
文苑(2018年22期)2018-11-19
作文与考试·小学低年级版(2016年18期)2016-09-10
