
K 亚群禽白血病病毒细胞受体的鉴定
2020-11-05 02:26邢立晓于蒙蒙常方方包媛玲王素艳祁小乐王笑梅高玉龙
中国预防兽医学报 2020年8期
邢立晓,俞 燕,于蒙蒙,常方方,包媛玲,王素艳,高 立,祁小乐,王笑梅,高玉龙*
(1. 中国农业科学院哈尔滨兽医研究所兽医生物技术国家重点实验室/禽免疫抑制病研究创新团队,黑龙江 哈尔滨 150069;2. 江苏省家禽科学研究所,江苏 扬州 225125)
囊膜病毒必须将病毒囊膜与宿主细胞膜融合才能启动感染[1]。在逆转录病毒中,这种膜融合是通过病毒囊膜糖蛋白(Envelope glycoprotein,Env)与宿主细胞受体相互作用而完成的[2]。Env 是由病毒囊膜表面亚单位(Surface subunit,SU:gp85)和跨膜亚单位(Transmembrane subunit,TM:gp37)二聚体组成的异源三聚体[3]。Env 通过SU 与细胞受体结合,介导病毒吸附。SU 对细胞受体的识别并结合决定了该病毒的宿主特异性[4]。与受体结合且经低pH 处理后,TM 发生构象改变,暴露出融合肽,将细胞膜与病毒囊膜拉近,完成膜融合[5]。无论是病毒的初始感染还是随后的病毒传播均取决于有效的结合细胞受体进入易感宿主的能力,因此鉴定细胞受体对于阐明病毒感染侵入机制至关重要。
作为囊膜病毒的成员之一,禽白血病病毒(Avian leukosis virus,ALV)属于逆转录病毒科(Retroviridae)α 逆转录病毒属(Alpharetrovirus)。根据其宿主范围、受体结合和干扰模式的不同,ALV 分为A~K 11个亚群[6]。其中A、B、C、D、J、K 为外源性病毒,E 为内源性病毒。A、B、J、K 亚群ALV 为常见感染家禽的外源性病毒。A 和B 亚群ALV 感染家禽通常引起淋巴细胞瘤[7],而J 亚群ALV(ALV-J)感染常引起髓细胞瘤[8]。ALV-K 是2012年首次从中国本地鸡——芦花鸡中分离得到的一个ALV 新亚群[9]。该病毒可能在我国本地鸡群中已长期存在,作为一种可垂直传播且易变异的逆转录病毒,给我国本地鸡群禽白血病的防控与净化带来挑战,也严重威胁我国本地鸡的种源安全和家禽产业的健康发展。不同ALV 亚群病毒通常利用宿主细胞的不同受体介导其感染。研究表明,鸡体内的低密度脂蛋白相关蛋白(Tva)由tva 基因编码,能够介导ALV-A 的感染[10]。tvb 基因编码Tvb 蛋白,能够介导ALV-B、ALV-D 和ALV-E的感染,属于肿瘤坏死因子受体家族[10-11]。Tvc蛋白由tvc 基因编码,能够介导ALV-C 的感染,属于免疫球蛋白超家族成员[12]。钠氢离子交换体1(NHE1)蛋白由鸡体内的管家基因nhe1 编码[13],能够介导ALV-J的感染。ALV-K是新出现的ALV亚群,其侵入细胞和感染机制还不清楚,本研究利用gp85蛋白受体交叉干扰试验、蛋白互作试验、受体结合和受体重建等一系列试验鉴定了ALV-K的细胞受体,以期为ALV侵入宿主细胞机制及筛选抗ALV 靶点研究奠定基础。
1 材料与方法
1.1 病毒、细胞株及质粒ALV-K(JS15SG01 株)为本实验室前期分离并保存。表达绿色荧光蛋白的重组病毒RCAS-K-GFP、RCAS-A-GFP 和RCAS-J-GFP以及HEK293T细胞、DF-1细胞、pCAGGS载体均为本实验室保存。pCAGGS-s-ALV-A-gp85-Fc、pCAGGSs-ALV-J-gp85-Fc 病毒蛋白真核表达质粒以及跨膜表达Tva 蛋白的pCAGGS-tva-Flag 质粒、可溶性表达Tva 蛋白的pCAGGS-tva-HA-His 质粒均为本实验室前期构建并保存[14]。
1.2 主要试剂鼠抗His 标签单克隆抗体(MAb)购自北京博奥龙生物公司;FITC 标记山羊抗人Fc IgG、鼠抗HA 标签MAb、鼠抗Flag 标签MAb、TRITC 标记山羊抗鼠IgG、FITC 标记兔抗鼠IgG、抗HA 琼脂磁珠均购自Sigma 公司;Steady-Glo Luciferase Assay System购自Promega 公司;质粒提取试剂盒购自QIAGEN 公司;Alexa Fluor 488 标记的驴抗小鼠IgG、蛋白Marker均购自Thermo scientific 公司;IRDye800CW 标记的山羊抗小鼠IgG购自LI-COR Bioscience公司;IRDye800CW直接标记山羊抗人Fc IgG 购自Sigma 公司;NP40 细胞裂解液、DAPI 染色液,购自上海碧云天生物技术有限公司;HiTrapTMProtein A HP-5 mL 预装柱、NiSepharose High Performance 预装柱、HiLoad 16/600 Superdex 200 prep grade 均 购 自GE 公 司;Polyjet 转 染 试 剂购自SignaGen Laboratories 公司;Lipofectamine 2000转染试剂购自Invitrogen 公司。
1.3 ALV-K gp85 蛋白真核表达质粒的构建与鉴定根据实验室分离的ALV-K JS15SG01 株gp85 基因序列合成其DNA片段。通过融合PCR在该序列5'端融合信号肽基因片段,同时在其3'端融合人IgG 恒定区基因片段(Fc),从而获得融合信号肽和Fc基因序列的可溶性gp85基因片段。将上述基因克隆到pCAGGS载体中构建真核表达质粒pCAGGS-s-ALV-K-gp85-Fc。重组质粒经菌液PCR 鉴定并经测序鉴定。
1.4 Tva 蛋白可溶性及跨膜表达的检测利用Lipofectamine 2000 转染试剂,按照说明书转染10 μg重组质粒pCAGGS-tva-HA-His 至铺有HEK293T 细胞的平皿中,同时以转染pCAGGS 空载体的细胞作为阴性对照,转染后48 h,收集细胞上清,常规处理后经12.5% SDS-PAGE 凝胶电泳,再转移至硝酸纤维素膜上。以鼠抗HA MAb(1∶3 000)为一抗,以IRDye800CW 标记的山羊抗鼠IgG(1∶20 000)为二抗,经western blot 检测可溶性Tva(sTva)蛋白的表达。
利用上述转染试剂转染2 μg 重组质粒pCAGGStva-Flag 至铺有HEK293T 细胞的激光共聚焦小皿中。转染后24 h,经4%多聚甲醛固定及5% BSA 封闭后,以鼠抗Flag MAb(1∶200)为一抗,以Alexa Fluor 488 标记的驴抗小鼠IgG(1∶200)为二抗,DAPI结合细胞核后,经Leica SP2 共聚焦系统检测Tva 在细胞表面的表达情况。
1.5 不同亚群ALV gp85-Fc蛋白的真核表达及gp85-Fc 和sTva 蛋白纯化效果的检测将pCAGGS-s-ALVA-gp85-Fc、pCAGGS-s-ALV-K-gp85-Fc、pCAGGS-s-ALV-J-gp85-Fc 重组质粒分别转染至密度为80%的HEK293T 细胞中,以转染pCAGGS 空载体的HEK293T 细胞做为阴性对照。48 h 后收集上清,经SDS-PAGE 电泳后,转移至硝酸纤维素膜上。以IRDye800CW标记的山羊抗人Fc IgG(1∶20 000)经western blot 检测ALV-A、ALV-K 和ALV-J gp85-Fc 蛋白的表达。确定上述ALV gp85-Fc 蛋白及1.4 中sTva 蛋白表达后按照制备型液相层析系统(AKTA Explore)操作说明分别对表达有ALV-A、ALV-K、ALV-J gp85-Fc 和sTva 的细胞上清进 行Protein A 和Ni 亲和层析纯化,制备纯化的gp85-Fc 和sTva 蛋白并经SDS-PAGE 检测纯化效果。
1.6 ALV-A 和ALV-K 的受体交叉干扰试验ALVK gp85 蛋白氨基酸序列与ALV-A gp85 蛋白氨基酸序列同源性最高,在80.8%以上。为了检测二者是否共用细胞受体,本研究进行了ALV-K 和ALV-A重组病毒的受体交叉干扰试验。将500 μL 纯化后的ALV-K gp85、ALV-A gp85 蛋白和本实验室前期表达的Fc 蛋白[14]分别稀释为50 μg/mL、100 μg/mL、200 μg/mL 3 种浓度,与DF-1 细胞4 ℃孵育1 h 后,将重组病毒RCAS-A-GFP 以104TCID50/孔的剂量感染不同浓度ALV-K gp85 孵育后的DF-1 细胞;将重组病毒RCAS-K-GFP 以104TCID50/孔的剂量感染不同浓度ALV-A gp85 孵育后的DF-1 细胞。将重组病毒RCAS-K-GFP 以104TCID50/孔的剂x 量感染Fc 孵育后的DF-1 细胞作为阴性对照,以相应重组病毒感染的正常DF-1 细胞(不加相应的gp85 蛋白)作为阳性对照。感染后24 h 在倒置荧光显微镜下观察各组细胞的荧光情况。
1.7 ALV-K gp85 与sTva 蛋白相互作用的Pulldown 检测利用Pull-down 试验体外验证ALV-K gp85 与sTva 蛋白的相互作用。将80 μL 抗HA 琼脂磁珠与20 μg 纯化后的sTva 蛋白在4 ℃孵育2 h 后,将磁珠分别与转染了pCAGGS-s-ALV-A-gp85-Fc、pCAGGS-s-ALV-K-gp85-Fc 和pCAGGS-s-ALV-Jgp85-Fc 质粒的HEK293T 细胞裂解液一起4 ℃孵育2 h 后,通过SDS-PAGE 分离结合的蛋白,转印至硝酸纤维素膜,以鼠抗HA MAb(1∶3 000)为一抗,以IRDye800CW 标记的山羊抗鼠IgG(1∶20 000)为二抗, 经western blot 检 测sTva 蛋 白 的 表 达。 以IRDye800CW 直接标记的山羊抗人Fc IgG 为抗体经western blot 检测gp85 蛋白是否被sTva 蛋白沉淀,从而确定sTva 和gp85 蛋白之间是否存在互作。将sTva蛋白与pCAGGS-s-ALV-A-gp85-Fc 转染细胞的裂解物孵育后作为阳性对照,将sTva 蛋白与pCAGGSs-ALV-J-gp85-Fc 转染细胞的裂解产物孵育后作为阴性对照。通过近红外荧光扫描成像系统进行Pulldown 结果分析。
1.8 ALV-K gp85 与sTva 蛋白相互作用的Co-IP 检测利用IP 试验验证ALV-K gp85 与sTva 蛋白在细胞内的相互作用。利用Lipofectamine 2000 转染试剂,将pCAGGS-s-ALV-A-gp85-Fc、pCAGGS-s-ALV-K-gp85-Fc、pCAGGS-s-ALV-J-gp85-Fc 分别和pCAGGS-tva-HA-His 重组质粒共转染6 孔板中培养的HEK293T 细胞(每孔2 μg)。转染后48 h,利用NP40 裂解液裂解各组细胞后,分别取40 μL 上清液检测各蛋白的表达情况。其余的上清液与抗HA 琼脂磁珠在4 ℃下过夜孵育,利用预冷的PBS 洗涤后,通过SDS-PAGE 分离蛋白质,按1.7 中抗体孵育方法及阴阳性对照设置,经western blot 检测ALV-K gp85 与sTva 在HEK 293T 细胞内 的互作。
1.9 ALV-K gp85 蛋白与跨膜表达的Tva 蛋白结合的检测将跨膜表达Tva 蛋白的pCAGGS-tva-Flag 重组质粒转染HEK293T 细胞后24 h,弃去培养液,PBS 洗涤3 次后,胰酶消化细胞,3 000 r/min 离心,将细胞沉淀用含5%FBS 的预冷PBS 洗涤3 次。将约2×106个细胞与500 μL ALV-K gp85 蛋白(200 ng/μL)冰上孵育1 h,以FITC 标记的山羊抗人Fc IgG(1∶200)与细胞作用1 h,以鼠抗Flag MAb(1∶200)为一抗,TRITC 标记的羊抗鼠IgG(1∶200)为二抗。按照上述操作,将跨膜表达的Tva 蛋白与ALV-A gp85 蛋白孵育后作为阳性对照,其与ALV-J gp85 蛋白孵育后作为阴性对照。通过流式细胞术(Fluorescence activated cell sorting,FACS)检测ALV-K gp85与跨膜表达的Tva 的结合情况。以ALV-A gp85 蛋白与跨膜表达的Tva 的结合率作为100%,计算ALV-K gp85 蛋白与跨膜表达的Tva蛋白的相对结合率。
1.10 Tva 介导RCAS-K-GFP 重组病毒进入非易感细胞中的检测为进一步验证Tva 蛋白是ALV-K 的细胞受体,本研究进行了ALV 非易感细胞的受体重建试验。将HEK293T 细胞按5×105个/孔传至12 孔板。待细胞长至80%,利用Lipofectamine 2000 转染试剂转染pCAGGS-tva-Flag 重组质粒(每孔1 μg),同时转染pCAGGS-Flag 质粒作为空载体对照。转染24 h 后,将重组病毒RCAS-K-GFP 按照104TCID50/的孔的剂量分别感染上述转染各重组质粒的HEK293T细胞。以重组病毒RCAS-A-GFP 按照104TCID50/孔的剂量感染转染pCAGGS-tva-Flag 的HEK293T细胞作为阳性对照,以重组病毒RCAS-J-GFP 按照104TCID50/孔的剂量感染转染pCAGGS-tva-Flag 的HEK293T细胞作为阴性对照。在感染后24 h 通过倒置荧光显微镜观察细胞荧光判断病毒的进入情况。
2 结 果
2.1 ALV-K gp85 真核表达质粒的构建与鉴定将经融合PCR 后获得的基因片段插入pCAGGS 载体,构建重组质粒pCAGGS-s-ALV-K-gp85-Fc。将重组质粒用特异性引物进行PCR 扩增,并测序鉴定。结果表明插入的目的基因序列正确,重组质粒正确构建。
2.2 不同亚群ALV gp85、Tva 蛋白可溶性及跨膜表达的检测结果将ALV-A、ALV-K 和ALV-J gp85重组质粒分别转染HEK293T 细胞,48 h 后,收取细胞上清经SDS-PAGE 检测结果显示,转染重组质粒的细胞上清均出现了分子量约为100 ku 的目的蛋白,而转染pCAGGS 空载体的阴性对照(NC)则无相应条带(图1A),这表明ALV-A、ALV-K 和ALV-J gp85 正确表达并分泌至上清。将pCAGGS-tva-HAHis 质粒转染HEK293T 细胞,48 h 后,收取细胞上清经western blot 鉴定sTva蛋白的表达。结果显示得到与预期分子量(35 ku)大小一致的sTva 蛋白(图1B)。分别利用Protein A亲和层析对各ALV gp85蛋白与sTva蛋白的纯化结果显示,纯化效果较好。将pCAGGS-tva-Flag 质粒转染HEK293T 细胞,24 h 后利用抗Flag MAb 和驴抗小鼠IgG 染色后,激光共聚焦显微镜观察显示,HEK293T 细胞膜上出现绿色荧光(图1C),表明Tva 蛋白跨膜表达在HEK293T 细胞膜上。
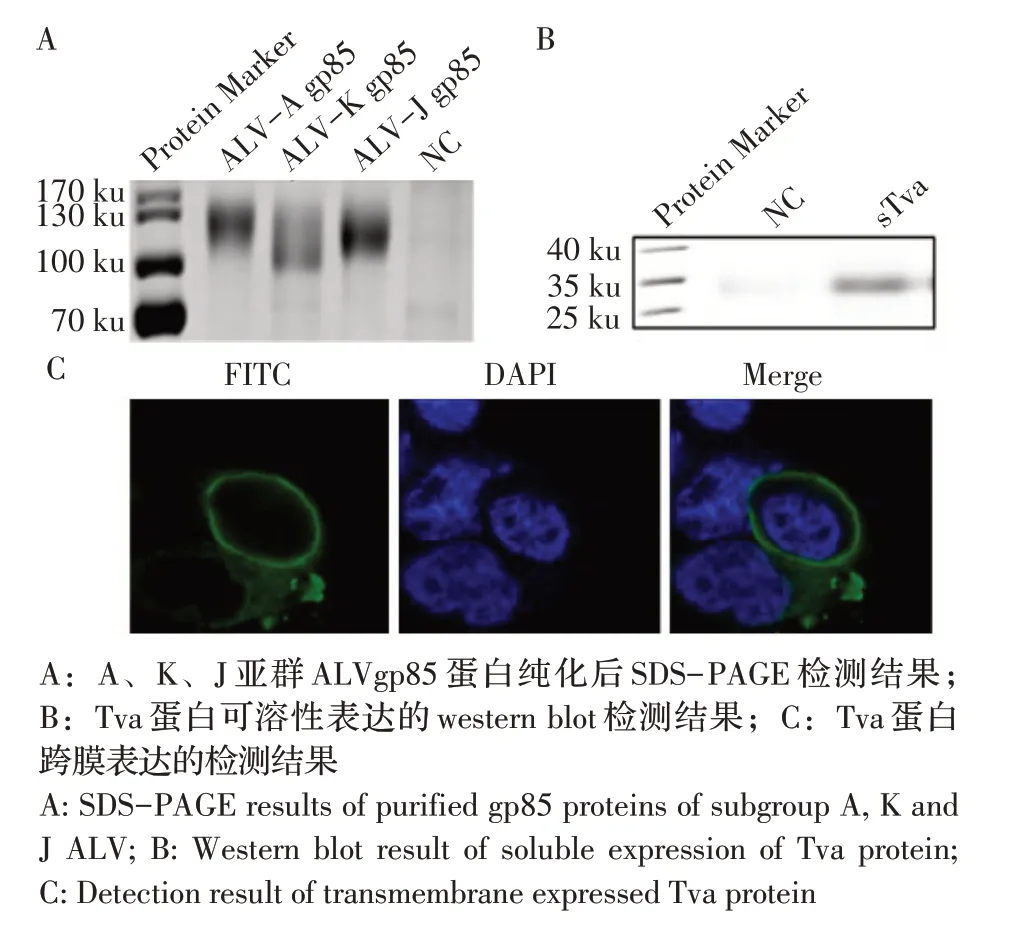
图1 不同亚群ALV gp85、Tva 蛋白可溶性及Tva 蛋白跨膜表达的检测结果Fig.1 Identification of protein expression of different subgroups ALV gp85,soluble and transmembrane Tva
2.3 ALV-A、ALV-K 重组病毒的受体交叉干扰试验结果将不同浓度的ALV-K gp85 蛋白和ALV-A gp85 蛋白分别与DF-1 细胞作用以封闭细胞表面病毒受体,再分别对应感染RCAS-A-GFP 和RCAS-KGFP 重组病毒,24 h 后在倒置荧光显微镜下观察可见,随着ALV-K gp85 蛋白浓度的升高,感染重组病毒的DF-1 细胞的绿色荧光逐渐减少,表明ALVK gp85 蛋白显著抑制了RCAS-A-GFP 重组病毒的感染;同样随着ALV-A gp85 蛋白浓度的升高,感染重组病毒的DF-1 细胞绿色荧光逐渐减少,表明ALV-A gp85 蛋白显著抑制了RCAS-K-GFP 重组病毒的感染;而RCAS-K-GFP 感染不同浓度Fc 蛋白作用后的DF-1 细胞产生的绿色荧光则无明显差异(图2)。与DF-1细胞共孵育后的ALV-A gp85蛋白能够明显抑制大部分ALV-K 的感染;同理,与DF-1 细胞共孵育后的ALV-K gp85 也可以竞争性抑制大部分ALV-A 的感染。表明ALV-A 和ALV-K 存在受体交叉干扰现象,它们可能共用一个细胞受体进入细胞。
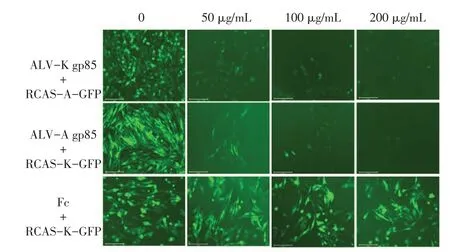
图2 ALV-A 和ALV-K 重组病毒受体的交叉干扰试验结果Fig.2 The result of cross-interference of ALV-A and ALV-Krecombination viruses
2.4 ALV-K gp85 与sTva 相互作用的Pull-down 和Co-IP 检测结果为验证ALV-K gp85 能否与sTva 蛋白互作,将纯化的sTva 蛋白分别与转染了ALV-A、ALV-K、ALV-J gp85 表达质粒的HEK293T 细胞裂解物共孵育后,利用Pull-down 试验检测sTva 蛋白与上述ALV gp85 的相互作用。结果显示,sTva 蛋白能够沉淀转染ALV-K gp85 表达质粒的HEK293T 细胞中的gp85 蛋白,sTva 蛋白也能够将转染ALV-A gp85 表达质粒的阳性对照中的gp85 蛋白沉淀,但sTva 蛋白不能将转染ALV-J gp85 表达质粒的阴性对照中的gp85蛋白沉淀(图3A)。表明ALV-K gp85 蛋白能够与细胞中的sTva 蛋白相互作用。
进一步利用IP 试验验证ALV-K gp85 与sTva 蛋白在细胞内的相互作用。利用ALV-K gp85 沉淀sTva的Co-IP 检测结果显示,共转染ALV-K gp85 和sTva表达质粒的HEK293T 细胞裂解物中,ALV-K gp85能够将sTva 沉淀(图3B),共转染ALV-A gp85 和sTva 表达质粒的阳性对照细胞中ALV-A gp85 也能将sTva 蛋白沉淀,但共转染ALV-J gp85 和sTva 表达质粒的阴性对照细胞中ALV-J gp85 则不能将sTva 蛋白沉淀。以上结果充分证明ALV-K gp85 蛋白和HEK293T 细胞中的sTva 蛋白存在相互作用。
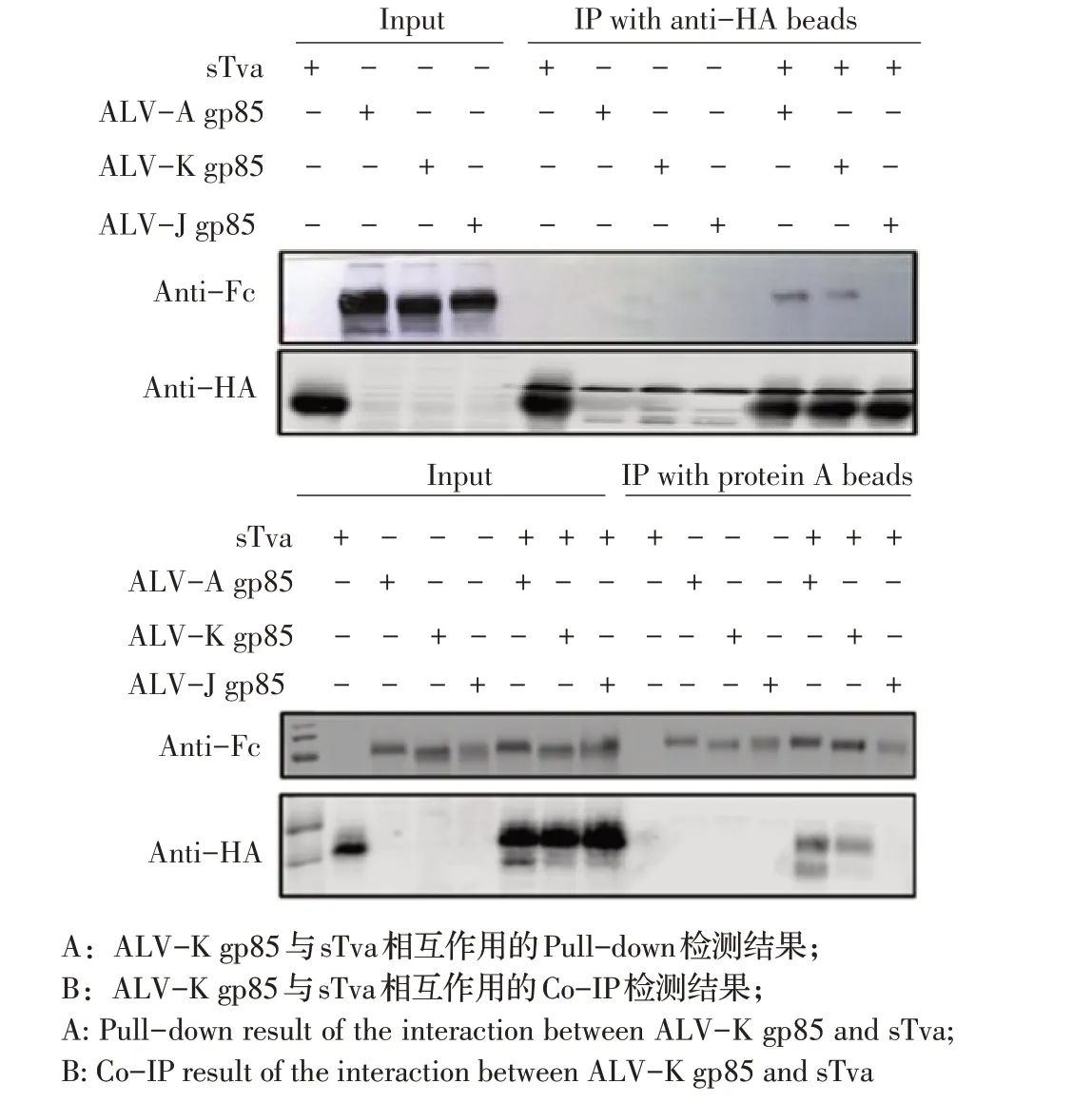
图3 ALV-K gp85 蛋白与sTva 蛋白互作的检测结果Fig.3 The result of the interaction between ALV-K gp85 and sTva
2.5 ALV-K gp85 与跨膜表达的Tva 结合的检测结果为进一步证明ALV-K gp85 与Tva 蛋白的结合能力,本研究将pCAGGS-tva-Flag 质粒转染HEK293T细胞36 h 后,孵育ALV-K gp85 蛋白,利用FACS 检测ALV-K gp85 与跨膜表达的Tva 蛋白的结合情况。经计算结果显示,ALV-K gp85 蛋白与跨膜表达的Tva 蛋白相对结合率超过90%,与阳性对照中ALV-A gp85 蛋白与跨膜表达Tva 蛋白的结合能力相近,而阴性对照中ALV-J gp85 蛋白与跨膜表达Tva 蛋白的结合率低于15%(图4)。表明ALV-K gp85 蛋白能特异识别并结合HEK293T 细胞膜表面的Tva 蛋白。

图4 ALV-K gp85 蛋白与跨膜表达的Tva 蛋白结合的检测结果Fig.4 The results of ALV-K gp85 binding to transmembrane expressed Tva proteina
2.6 Tva 介导RCAS-K-GFP 重组病毒进入非易感细胞的检测结果将pCAGGS-tva-Flag 重组质粒转染HEK293T 细胞,24 h 后,感染重组病毒RCAS-KGFP,通过倒置荧光显微镜观察可见,与阳性对照RCAS-A-GFP 感染表达Tva 蛋白的细胞相似,感染RCAS-K-GFP 的HEK293T 细胞也出现较强绿色荧光(图5)。而空载体对照组(RCAS-K-GFP 感染空载体pCAGGS-Flag 转染的HEK293T 细胞)和阴性对照组(RCAS-J-GFP 感染表达Tva 蛋白的HEK293T 细胞)均无绿色荧光(图5),上述结果表明Tva 蛋白能够特异性介导ALV-K 进入非易感细胞,进一步证明其是ALV-K 的受体。
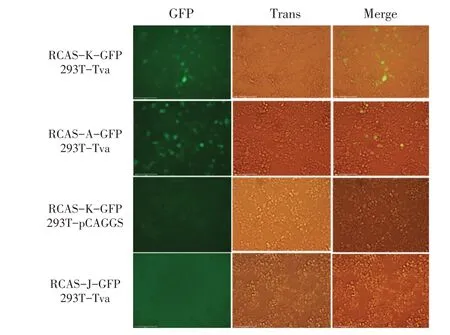
图5 Tva 介导RCAS-K-GFP 重组病毒进入非易感细胞的检测结果Fig.5 Tva mediates RCAS-K-GFP recombinant virus into non-susceptible cells
3 讨 论
病毒受体是存在于宿主细胞表面的细胞膜成分,在病毒感染过程中起着至关重要的作用,病毒识别并结合宿主细胞受体是开启病毒感染过程的第一步。ALV 囊膜蛋白表面亚单位gp85 蛋白识别并结合细胞受体,诱导跨膜亚单位gp37 蛋白的构象改变,形成六螺旋束,导致病毒囊膜与宿主细胞膜的融合,启动病毒的感染[15]。ALV-K 是新分离到的ALV,其gp85 基因与外源性ALV-A gp85 基因的同源性最高,而其pol、3'UTR 等基因与内源性ALV-E 有较高的同源性[16]。已有研究表明,ALV-K 的复制能力明显低于ALV-A[17],一方面可能是由于ALV-K 基因组中具有转录调控功能的长末端重复序列(LTR)与ALV-E同源性高,而内源性ALV-E 复制能力低下,其LTR转录调控能力弱,这导致ALV-K 基因组的转录翻译效率低;另一方面其复制能力低是否与其侵入易感宿主靶细胞的能力有关,还不得而知。为了阐明ALV-K 的侵入机制,本研究中对ALV-K 的细胞受体进行了鉴定。
ALV gp85 基因可划分为3 个可变区(Variable region,vr)和2 个宿主决定区(Host range region,hr),其中hr 是影响病毒与受体结合的主要区域[18]。ALVK gp85 蛋白全序列和受体决定区序列均与ALV-A 的同源性最高,为了确定ALV-K 是否与ALV-A 利用相同的细胞受体,本研究首先开展了ALV-A 和ALV-K 受体的交叉干扰试验。已有研究表明,当两种病毒共用同一受体时,在其易感细胞表面受体有限的前提下,一种病毒的表面蛋白与细胞受体结合,会竞争性地抑制另一种病毒吸附在该受体上,即前期结合的病毒蛋白会对后者产生“占位效应”[19]。本研究利用具有生物学活性的ALV-K gp85蛋白预先结合DF-1 细胞上的受体,模拟ALV-K 的感染,再接种RCAS-A-GFP 以模拟ALV-A 的超感染。通过荧光显微镜观察到ALV-K 和ALV-A 对宿主细胞的感染具有强烈的交叉干扰现象,即ALV-K gp85 蛋白与受体的结合抑制了ALV-A 的感染,造成ALV-A 感染力明显减弱。这种基于受体产生的交叉干扰现象提示这两种病毒很可能共用同一种细胞受体。逆转录病毒由于共用同一受体导致交叉干扰的现象之前也有报道,如长臂猿白血病病毒(GALV)与B 亚群猫白血病病毒(FeLV-B)共用相同细胞受体导致的受体交叉干扰[20]。一种病毒感染靶细胞后干扰另一种病毒的超感染也常被用来确定这两种病毒是否利用相同的受体[21]。
已有研究表明Tva 为ALV-A 的细胞受体[22]。为了进一步证明Tva 也能介导ALV-K 进入宿主细胞,本研究利用ALV-K gp85 蛋白与sTva 蛋白的Pulldown 和Co-IP 试验进一步证明二者有直接相互作用。并且受体结合试验表明ALV-K gp85 蛋白能够高效与跨膜表达的Tva 结合。受体重建试验结果表明Tva 能够介导ALV-K 感染非易感细胞。以上结果充分证明了ALV-K 的细胞受体也是Tva 蛋白。虽然ALV-A 和ALV-K 共用同一细胞受体,但ALV-K 感染的宿主范围以及感染后引起的临床症状与ALV-A存在差异,这可能与病毒利用受体中的不同位点介导其结合有关。这种现象类似于B、D、E 亚群ALV共用Tvb 受体,但是B 亚群ALV 主要与Tvb N 末端的L36、Q37、L41、Y42 产生相互作用,而E 亚群ALV主要与TvbS1中的Y67、N72、D73 3 个氨基酸组成的决定簇结合[23]。而哪些氨基酸决定Tva 的受体功能以及其与ALV-K gp85 蛋白的哪些氨基酸结合尚不清楚,需要进一步研究。总之,ALV-K 细胞受体的鉴定,对于理解ALV 的感染机制,ALV 与宿主的相互作用以及抗病毒靶点研究有着极为重要的意义。
猜你喜欢
工业水处理(2022年12期)2022-12-29
中华实验眼科杂志(2022年8期)2022-11-15
生殖医学杂志(2022年10期)2022-10-19
中国医药科学(2022年5期)2022-05-05
中风与神经疾病杂志(2021年9期)2021-11-08
山东医药(2020年19期)2020-12-29
医药前沿(2020年28期)2020-12-02
科学导报(2020年25期)2020-04-28
中小学德育(2020年11期)2020-03-18
标记免疫分析与临床(2016年9期)2016-11-21
