
基于SRAP分子标记的不同种壳厚度油棕种质资源遗传多样性分析
2020-12-11 09:10周丽霞曹红星
南方农业学报 2020年9期
周丽霞 曹红星


摘要:【目的】分析不同種壳厚度油棕种质资源的遗传多样性及群体结构,为油棕种质资源的有效利用及新品种选育提供理论依据。【方法】选取厚壳种BM8和无壳种L2T对288对SRAP引物组合进行筛选,从中筛选出扩增条带清晰、稳定及多态性好的SRAP引物组合,利用其对46份不同种壳厚度的油棕种质材料进行多态性扩增,基于扩增结果,利用NTSYS 2.1的非加权组平均法(UPGMA)计算遗传相似系数并构建聚类图,利用POPGENE 1.32计算遗传多样性指数。【结果】从288对引物组合中共筛选出15对SRAP引物,利用其对46份油棕种质材料进行PCR扩增,共扩增出303条条带,其中多态性条带183条,平均每条引物扩增出12.2条,多态比率为60.4%。46份油棕种质材料的观测等位基因数(Na)为1.0982~1.3264,平均1.6024;有效等位基因数(Ne)为1.1092~1.15976,平均1.4803;Neis基因多样性指数(H)为0.0572~0.1093,平均0.1937;I为0.0927~0.1648,平均0.3115。薄壳种油棕的遗传多样性指数与无壳种油棕较接近,且二者均高于厚壳种油棕,说明薄壳种和无壳种油棕的遗传多样性高于厚壳种油棕。厚壳种油棕与无壳种油棕的遗传一致度最小(0.7728),但二者间的遗传距离最大(0.2556)。厚壳种油棕与薄壳种油棕间的遗传一致度最大,(0.8396),且二者间的遗传距离最小(0.1748)。在遗传相似系数为0.59时,46份供试油棕种质被分为4个类群,其中,厚壳种油棕种质材料均分布在第I类群,薄壳种油棕种质材料(除Eg14分布在第I类群外)和无壳种油棕种质材料均分布在II、III和IV类群。【结论】46份油棕种质材料的遗传多样性整体较丰富,其中薄壳种油棕和无壳种油棕的遗传多样性较厚壳种油棕丰富,二者可作为优良育种材料进行亲本选配,选育出高产油率的油棕新品种。
关键词: 油棕;种壳;厚度;SRAP;遗传多样性;群体结构
中图分类号: S565.9 文献标志码: A 文章编号:2095-1191(2020)09-2097-07
Genetic diversity analysis of oil palm germplasm resources with different seed shell thicknesses based on SRAP molecular marker
ZHOU Li-xia, CAO Hong-xing*
(Coconut Research Institute, Chinese Academy of Tropical Agricultural Science/Hainan Provincial Key Laboratory of Tropical Oil Crops Biology, Wenchang, Hainan 571339, China)
Abstract:【Objective】To study the population structure and genetic diversity of oil palm germplasm resources with different shell thicknesses and provide theoretical basis for the effective utilization of oil palm germplasm resources and the selection of new varieties. 【Method】A total of 288 SRAP primer pairs were selected with BM8(thick shell) and L2T (shellless). The 46 oil palm germplasm materials with different shell thicknesses were amplified using the SRAP primer with clear, stable and polymorphic. unweighted pair-group method with arithmetic mean(UPGMA) of NTSYS 2.1 was used to calculate genetic similarity coefficient and construct cluster graph, POPGENE 1.32 was used to calculate genetic diversity index. 【Result】A total of 15 primer pairs with clear amplification and rich polymorphism were selected from 288 pairs,and 303 clear bands were amplified totally from 46 oil palm germplasm materials by PCR,of which 183 were polymorphism sites with an average of 12.2 bands by each primer, and the polymorphic ratio was 60.4%. Observed number of alleles(Na) of 46 oil palm germplasm materials was from 1.0982 to 1.3264, with an average of 1.6024,the effective number of alleles(Ne) was from 1.1092 to 1.15976, with an average of 1.4803, Neis gene diversity(H) was from 0.0572 to 0.1093, with an average of 0.1937, Shannons information index(I) was from 0.0927 to 0.1648, with an average of 0.3115. The genetic diversity index of thin shell oil palm was close to that of shellless oil palm, and both of them were higher than that of thick shell oil palm, indicating that the polymorphism of thin shell and shellless oil palm were higher than that of thick shell oil palm. The genetic consistency between the thick shell and shellless oil palm was the lowest (0.7728), but the genetic distance between them was the largest(0.2556). The genetic consistency between thick shell and thin shell oil palm was the highest(0.8396), and the genetic distance between them was the smallest(0.1748). When the genetic similarity coefficient was 0.59, 46 oil palm germplasms were divided into 4 subgroups. Among them, thick shell materials were all distributed in subgroup I, thin shell materials(except Eg14 distributed in subgroup I) and shellless materials were distributed in subgroups II, III and IV. 【Conclusion】The genetic diversity of 46 oil palm germplasm materials is relatively rich, among which thin shell and shellless oil palm have more genetic diversity than the thick shell oil palm. They can be used as excellent breeding materials for parent selection and breeding new oil palm varieties with high oil yield.
Key words:oil palm; shell; thickness; SRAP; genetic diversity; population structure
Foundation item:Species and Varieties Conservation of Sector Project of the Ministry of Agriculture and Rural Affairs(125163015000160004);Basic Research Project of Chinese Academy of Tropical Agricultural Sciences(1630152017 008);Tropical Woody Oil Plant Industry Technology Innovation Group Project(17CXTD-13); Construction Project of Indonesia Experimental Agricultural Station of the Chinese Academy of Tropical Agricultural Sciences(SYZ2019-12)
0 引言
【研究意義】油棕(Elaeis guineensis Jacq.)属棕榈科多年生乔木植物,是世界上产油率最高的油料作物,也是热带地区主要的木本经济作物之一(曹红星等,2015),享有“世界油王”的美誉(冯美利等,2017;Sowmya et al.,2019)。油棕果实由外果皮、中果皮、内果皮(种壳)和种仁4部分组成,其中内果皮发育成熟后形成类似椰子壳的纤维环,且其厚度决定种仁的大小,薄壳种油棕的产油量更高(Teh et al.,2017;石鹏等,2018)。我国自1926年开始在海南引种试种油棕,早期引进的品种如杜拉、丹那拉等在我国具有良好的适应性,但产油率较低(熊惠波等,2009)。近年来我国又从国外引进高产型油棕新品种,对于这些引进品种的亲缘关系、种群结构等尚不明确,严重影响油棕育种进程。因此,研究我国油棕种质的遗传多样性及群体结构,对油棕种质资源的保存、开发利用及新品种培育均具有十分重要的意义。【前人研究进展】相关序列扩增多态性(SRAP)分子标记具有共显性高、操作简单和多态性丰富等优点,已广泛应用于作物的遗传多样性分析、种质资源鉴定等方面。颜平等(2014)利用SRAP分子标记对144份海南钻喙兰进行种内遗传多样性分析,结果发现海南岛上的野生海南钻喙兰种质资源具有较高的遗传多样性和种内变异。陈熙等(2016)利用40对SRAP引物分析50份陕西茶树种质资源的遗传多样性,结果发现材料间的遗传多样性处于较高水平。廖柏勇等(2016)开发了20个SRAP分子标记,并用于分析来自我国17个省(区)31个苦楝种质资源的遗传多样性,结果发现苦楝遗传变异主要来源于种源内,种源间基因交流有限,种源遗传多样性整体偏低,而部分山区种源遗传多样性较高。张景云等(2017)开发了14个SSR和33个SRAP分子标记,并用于分析46份苦瓜种质遗传多样性,结果发现其遗传多样性丰富。目前,我国对油棕遗传多样性的研究报道较少,周丽霞等(2017)开发了27个多态性SSR引物,并用于分析海南8个油棕品种的遗传多样性,结果发现种间杂交程度较小,物种亲缘关系较远,遗传多样性良好。【本研究切入点】目前利用SRAP分子标记分析不同种壳厚度油棕种质遗传多样性和群体结构的研究鲜见报道。【拟解决的关键问题】开发多态性SRAP分子标记,并用于分析不同种壳厚度油棕种质资源的遗传多样性和群体结构,为油棕种质资源的有效利用和新品种选育提供理论依据。
1 材料与方法
1. 1 试验材料
供试的46份油棕种质分别来自海南文昌、云南保山和广西防城港(表1)。经多年种植和性状调查,根据油棕果实种壳厚度(雷新涛和曹红星,2013),将供试材料分为3个类型:Eg01~Eg13为厚壳种(种壳厚2.0~8.5 mm),Eg14~Eg30为薄壳种(种壳厚0.5~2.0 mm),Eg31~Eg46为无壳种(无核壳)。主要试剂:Taq DNA聚合酶、dNTPs、Mg2+以及DNA提取所需试剂和电泳所需试剂均购自海南省合辉实业有限公司,DL2000 DNA Marker购自天根生化科技(北京)有限公司,SRAP引物由生工生物工程(上海)股份有限公司合成。主要仪器设备:低温高速离心机(Eppendorf centrifuge 5804R,德国)、梯度核酸扩增仪(TP1600,澳大利亚)、垂直电泳系统(北京六一仪器厂)和核酸凝胶成像系统(G-BOXF3)。
1. 2 DNA提取
采集油棕嫩叶,1片叶/株,采用CTAB法提取其DNA(周丽霞等,2013),-20 ℃保存备用。
1. 2 SRAP引物筛选
选取厚壳种BM8和无壳种L2T对本课题组设计的288对SRAP引物组合进行筛选。反应体系15.0 μL:DNA模板1.0 μL,2×Taq PCR Master-Mix(含染料)8.0 μL,10 μmol/L正、反向引物各1.0 μL,ddH2O补足至15.0 μL。扩增程序:94 ℃预变性5 min;94 ℃ 30 s,35 ℃ 1 min,72 ℃ 30 s,进行5个循环;94 ℃ 30 s,54 ℃ 1 min,72 ℃ 30 s,进行30个循环;72 ℃延伸7 min。扩增产物用8%非变性聚丙烯酰胺凝胶电泳进行检测,最后用银染法显色并拍照。
1. 3 统计分析
用Excel 2017统计扩增出的条带数量,相同位置上有清晰可辨条带记为“1”,无清晰可辨条带记为“0”,从而构建原始数据矩阵。应用NTSYS 2.1的非加权组平均法(UPGMA)计算遗传相似系数并构建聚类图。应用POPGENE 1.32计算遗传多样性指数:观测等位基因数(Observed number of alleles,Na)、有效等位基因数(Effective number of alleles,Ne)、Neis基因多样性指数(Neis gene diversity,H)和Sha-nnons信息指数(Shannons information index,I)。
2 结果与分析
2. 1 SRAP引物筛选结果
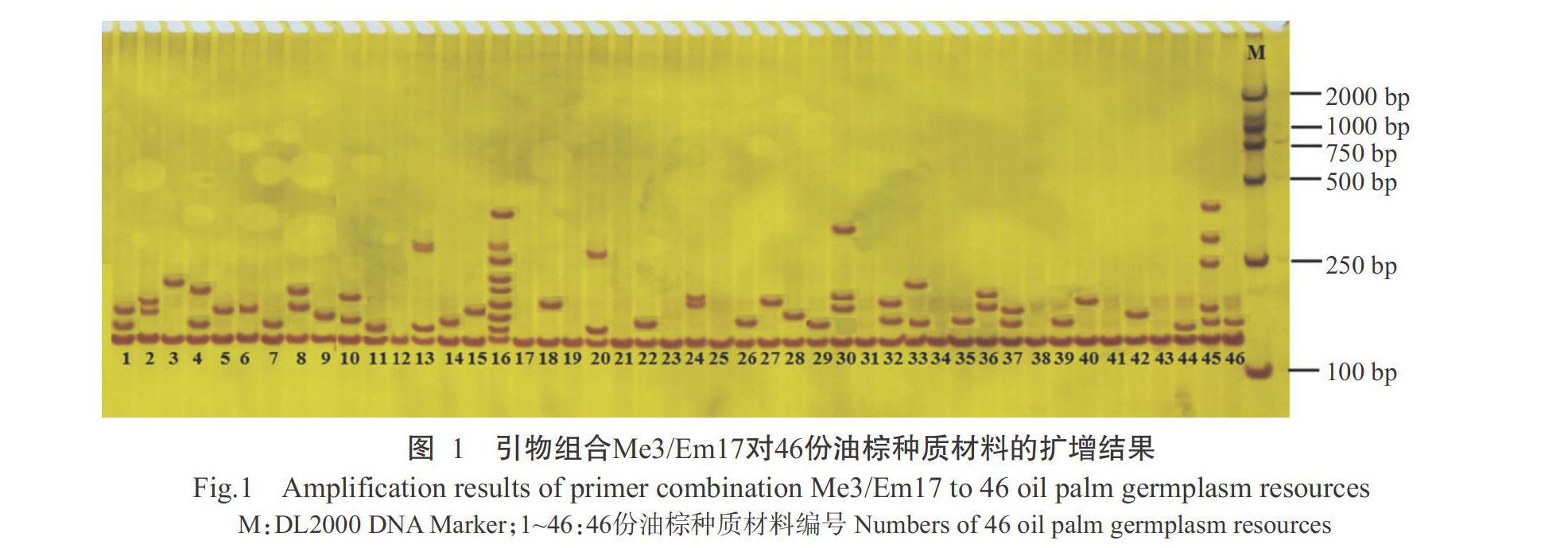
选取厚壳种BM8和无壳种L2T对288对SRAP引物组合进行筛选,根据扩增条带的清晰度、稳定性及多态性共筛选出15对SRAP引物组合,如表2所示。利用其对46份油棕种质材料进行PCR扩增,共扩增出303条条带,其中多态性条带183条,平均每对引物组合扩增出12.2条,多态比率为60.4%,表明供试油棕种质资源具有较丰富的遗传多样性。图1为引物组合Me3/Em17对46份油棕种质材料的扩增图谱,通过观察统计可发现不同油棕种质材料扩增结果存在明显差异,说明筛选出的SRAP分子标记多态性较好,可用于油棕种质资源的遗传多样性分析。
2. 2 油棕种质资源的遗传多样性分析结果
利用POPGENE 1.32计算46份油棕种质材料的遗传多样性指数,如表3所示。46份油棕种质材料的Na为1.0982~1.3264,平均1.6024;Ne为1.1092~1.15976,平均1.4803;H为0.0572~0.1093,平均0.1937;I为0.0927~0.1648,平均0.3115,说明46份油棕种质材料的遗传多样性较丰富。整体来说,薄壳种油棕的遗传多样性指数与无壳种油棕较接近,且二者均高于厚壳种油棕,说明薄壳种和无壳种油棕的多态性高于厚壳种油棕。
46份油棕种质材料的遗传一致度和遗传距离如表4所示。厚壳种油棕与无壳种油棕的遗传一致度最小,为0.7728,但二者间的遗传距离最大,为0.2556,说明厚壳种油棕与无壳种油棕的亲缘关系最远。厚壳种油棕与薄壳种油棕间的遗传一致度最大,为0.8396,且二者间的遗传距离最小,为0.1748,说明厚壳种油棕与薄壳种油棕的亲缘关系最近。
基于油棕SRAP分子标记的图谱矩阵结果,利用NYSTS 2.1计算遗传相似系数,结果(表5)显示,46份油棕种质材料的遗传相似系数为0.31~0.97,其中,BM8与UR1和BM119的遗传相似系数最小,均为0.31,说明二者亲缘关系最远;BM387与YA69的遗传相似系数最大,为0.97,说明二者的亲缘关系最近。
2. 3 油棕种质资源的聚类分析
为了解46份供试油棕种质的群体遗传结构,应用UPGMA对其进行聚类分析,结果如图2所示。在遗传系数为0.59时,46份供试油棕种质被分为4个类群,第I类群含有14份油棕种质材料,其中有13份厚壳种,1份薄壳种;第II类群含有9份油棕种质材料,其中6份薄壳种和3份无壳种;第III类群含有11份油棕种质材料,其中8份薄壳种和3份无壳种;第IV亚群含有12份油棕种质,其中2份薄壳种和10份无壳种。可见,厚壳种油棕均分布在第I类群,薄壳种油棕(除Eg14分布在第I类群外)和无壳种油棕均分布在II、III和IV类群,表明厚壳种油棕种质材料间的亲缘关系较近,而薄壳种油棕种质材料间和无壳种油棕种质材料间的亲缘关系均较远,且二者遗传多样性高于厚壳种油棕种质。整体来看,46份油棕种质材料的遗传多样性较丰富。
3 讨论
经大量研究证实,SRAP分子标记技术简单易操作,多态性好,成本低,且较稳定,广泛受得研究者的青睐。本研究从288对SRAP引物组合中筛选出15对扩增条带清晰、稳定且多态性丰富的SRAP引物组合,用于分析46份不同种壳厚度的油棕种质遗传多样性,结果显示,PCR扩增出303个条带,其中多态性条带183条,多态比率为60.4%,与甜瓜中的多态比率(58.0%)(高宁宁等,2019)相近,但高于小白菜中的多态比率(32.9%)(李桂花等,2017)和平菇中的多态比率(30.1%)(常艳等,2017),说明SRAP分子标记能有效分析多种植物的遗传多样性。
Na、Ne、H和I是衡量植物物种间种质资源遗传多样性丰富度的常用指标。本研究发现,46份油棕种质材料的Na为1.0982~1.3264,平均1.6024;Ne为1.1092~1.15976,平均1.4803;H为0.0572~0.1093,平均0.1937;I为0.0927~0.1648,平均0.3115,说明46份油棕种质材料的遗传多样性较丰富,与棉花(刘雅辉等,2015)和油菜(张羽等,2017)的研究结论相似。其中,无壳种油棕的遗传多样性指数略高于薄壳种油棕,二者均高于厚壳种油棕,结果表明无壳种和薄壳种油棕种质的遗传多样性均高于厚壳种油棕,可能是由于厚壳种油棕是我国最早期从印度尼西亚引进,来源地范围较小,遗传基础较狭窄,亲缘关系较近,而薄壳种和无壳种是从马来西亚、尼日利亚及哥斯达黎加等多个国家引进,来源范围较广,遗传背景较丰富,亲缘关系也较远。本研究聚类分析結果显示,46份油棕种质材料分为4个类群,其中,厚壳种油棕种质材料集中分布在第I类群,而薄壳种和无壳种种质材料分散分布于II、III和IV类群,说明薄壳种油棕和无壳种油棕的遗传多样性较高,厚壳种油棕的遗传多样性较低,与上述结论相一致。此外,厚壳种和无壳种油棕种质的遗传一致度最低,为0.7728,遗传距离最大,为0.2556,也说明厚壳种和无壳种油棕间的遗传差异大,亲缘关系较远。厚壳种和薄壳种油棕种质的遗传一致度最高,为0.8396,遗传距离最小,为0.1748,说明二者间的遗传差异小,亲缘关系近。由此可见,SRAP分子标记能有效地区分油棕种质亲缘关系的远近。由于丰富且优异的种质资源是新品种选育的材料基础,只有明确种质资源的群体结构及遗传背景,才能对其进行充分有效地利用。今后可将薄壳种和无壳种油棕种质作为优良育种材料进行亲本选配,选育出高产油率的油棕新品种。
4 结论
46份油棕种质材料的遗传多样性整体较丰富,其中薄壳种油棕和无壳种油棕的遗传多样性较厚壳种油棕丰富,二者可作为优良的育种材料进行亲本选配,选育出高产油率的油棕新品种。
参考文献:
曹红星,雷新涛,刘艳菊. 2015. 不同来源地油棕种质资源耐寒适应性初步研究[J]. 西南农业学报,28(5):1916-1919. [Cao H X,Lei X T,Liu Y J. 2015. Preliminary study on cold resistance adaptability of oil palm germplasm resources from different origins[J]. Southwest China Journal of Agricultural Sciences,28(5):1916-1919.]
陈熙,张羽,李佼,李秀峰,席彦军. 2016. SRAP标记分析陕西省主要茶树种质资源遗传多样性[J]. 西北林学院学报,31(3):143-147. [Chen X,Zhang Y,Li J,Li X F,Xi Y J. 2016. Analysis of SRAP marker-based polymorphism of the main tea germplasms in Shanxi[J]. Journal of Northwest Forestry University,31(3):143-147.]
常艳,金青,凌磊. 2017. 16种平菇遗传关系的SRAP分析[J]. 安徽农业大学学报,44(5):834-838. [Chang Y,Jin Q,Ling L. 2017. Phylogenetic relationship analysis of 16 Pleurotus ostreatus species by SRAP markers[J]. Journal of Anhui Agricultural University,44(5):834-838.]
冯美利,李爱基,曹红星,金龙飞. 2019. 油棕林下养鸡对土壤养分和微生物数量的影响[J]. 江西农业学报,31(11):43-46. [Feng M L,Li A J,Cao H X,Jin L F. 2019. Effects of raising chicken in oil palm forest on soil nutrients and soil microorganisms[J]. Acta Agriculturae Jiangxi,31(11):43-46.]
高宁宁,常高正,康利允,李晓慧,梁慎,李海伦,赵卫星. 2019. 基于SRAP标记的甜瓜耐盐种质资源遗传多样性分析[J]. 西北植物学报,39(1):68-75. [Gao N N,Chang G Z,Kang L Y,Li X H,Liang S,Li H L,Zhao W X. 2019. Genetic diversity analysis of salt tolerance germplasm resources of melon based on SRAP[J]. Acta Botanica Boreali-Occidentalia Sinica,39(1):68-75.]
雷新涛,曹红星. 2013. 油棕[M]. 北京:中国农业出版社. [Lei X T,Cao H X. 2013. Oil palm[M]. Beijing:China Agriculture Press.]
李桂花,陈汉才,张艳. 2017. 小白菜种质遗传多样性与亲缘关系的SRAP和SSR分析[J]. 广东农业科学,44(5):37-45. [Li G H,Chen H C,Zhang Y. 2017. Genetic diversity and phylogenetic relationships analysis of Chinese ca-bbage germplasm resources by SRAP and SSR[J]. Guangdong Agricultural Sciences,44(5):37-45.]
廖柏勇,王芳,陈丽君,刘明骞. 2016. 基于SRAP分子标记的苦楝种质资源遗传多样性分析[J]. 林业科学,52(4):48-58. [Liao B Y,Wang F,Chen L J,Liu M Q. 2016. Genetic diversity of germplasm resources of Melia azedarach revealed by SRAP markers[J]. Scientia Silvae Sinicae,52(4):48-58.]
刘雅辉,王秀萍,鲁雪林. 2015. 棉花耐盐相关序列扩增多态性(SRAP)分子标记筛选[J]. 江苏农业学报,31(3):484-488. [Liu Y H,Wang X P,Lu X L. 2015. Selection of sequence-related amplified polymorphism molecular marker associated with salt tolerance of cotton[J]. Jiangsu Journal of Agricultural Sciences,31(3):484-488.]
石鵬,夏薇,肖勇. 2018. 油棕种壳厚度控制基因SHELL的SNP分子标记开发[J]. 广西植物,38(2):195-201. [Shi P,Xia W,Xiao Y. 2018. SNP markers development of SHELL controlling shell thickness in oil palm(Elaesis guineensis)[J]. Guihaia,38(2):195-201.]
熊惠波,李瑞,李希娟. 2009. 油棕产业调查分析及中国发展油棕产业的建议[J]. 中国农学通报,25(24):114-117. [Xiong H B,Li R,Li X J. 2009. Investigation,analysis and the advice of palm industry in China[J]. Chinese Agricultural Science Bulletin,25(24):114-117.]
颜平,黄明忠,陆顺教,杨光穗,唐燕琼,尹俊梅. 2014. 基于SRAP标记的海南钻喙兰种质资源遗传多样性分析[J]. 热带作物学报,35(9):1677-1682. [Yan P,Huang M Z,Lu S J,Yang G H,Tang Y Q,Yin J M. 2014. Genetic diversity analysis of Rhynchostylis gigantean(Lindl.) Ridi.s germplasm resources based on SRAP markers[J]. Chinese Journal of Tropical Crops,35(9):1677-1682.]
张景云,黄月琴,万新建,关峰,繆南生. 2017. 基于SSR和SRAP标记苦瓜种质遗传多样性分析[J]. 上海交通大学学报(农业科学版),35(3):90-94. [Zhang J Y,Huang Y Q,Wan X J,Guan F,Liao N S. 2017. Genetic diversity of Momordica charantia germplasm based on SSR and SRAP marker[J]. Journal of Shanghai Jiaotong University(Agricultural Science),35(3):90-94.]
周丽霞,吴翼,肖勇. 2017. 基于SSR分子标记的油棕遗传多样性分析[J]. 南方农业学报,48(2):216-221. [Zhou L X,Wu Y,Xiao Y. 2017. Genetic diversity of Elaeis guineensis Jacq. based on SSR markers[J]. Journal of Southern Agriculture,48(2):216-221.]
周丽霞,肖勇,杨耀东,马子龙. 2013. 油棕基因组DNA 3种提取方法的比较研究[J]. 江西农业学报,25(8):9-11. [Zhou L X,Xiao Y,Yang Y D,Ma Z L. 2013. Comparison of three extract methods for genomic DNA of Elaeis guineensis[J]. Acta Agriculturae Jiangxi,25(8):9-11.]
张羽,张骁婿,陈进. 2017. 43份油菜菌核病抗性资源SCoT、SSR与SRAP标记分析[J]. 西北农林科技大学学报(自然科学版),45(1):45-52. [Zhang Y,Zhang X X,Chen J. 2017. Germplasm analysis of 43 Sclerotinia sclerotiorum resistant Brassica napus by SCoT,SSR and SRAP mar-kers[J]. Journal of Northwest A & F University(Natural Science Edition),45(1):45-52.]
Sowmya K,Robert M,Richard S. 2019. The palm oil industry and no communicable diseases[J]. Policy & Practice,97(2):118-128.
Teh C K,Muaz S D,Tangaya P,Fong P Y,Ong A L,Mayes S,Chew F T,Kulaveerasingam H,Appleton D. 2017. Characterizing haploinsufficiency of SHELL gene to improve fruit form prediction in introgressive hybrids of oil palm[J]. Scientific Reports,7(1):3118-3132.
(責任编辑 陈 燕)
猜你喜欢
中国美容医学(2022年5期)2022-07-12
读者·校园版(2020年4期)2020-06-01
意林·少年版(2019年21期)2019-12-17
长江蔬菜·学术版(2016年12期)2017-01-12
天津农业科学(2015年5期)2015-05-30
