
抗、感青枯病桑树根际细菌群落结构与多样性
2020-12-16 03:00武华周娄德钊涂娜娜卢芙萍吉训聪王树昌
福建农业学报 2020年9期
武华周,娄德钊,2,涂娜娜,2,耿 涛,卢芙萍,吉训聪,王树昌
(1. 中国热带农业科学院环境与植物保护研究所,海南 海口 571101;2. 海南大学,海南 海口 570228;3. 海南省农业科学院植物保护研究所 海南 海口 571100;4. 海南省植物病虫害防控重点实验室,海南 海口 571100)
0 引言
【研究意义】植物根际微生物对于植物生长、营养与健康有着深远的影响。植物根际包含有巨大数量微生物与无脊椎动物,被认为是地球上动态性最强、最有活力的一种交界面[1−2]。根际微生物的结构和丰度的改变是植物健康的重要预警器。已有研究表明植物在受到病虫害攻击时,会特异招募有益微生物,形成微生态系统-植物根部的“生物屏障”,达到阻止病原菌入侵的目的[3]。在自然条件下,植物及多种生态因子复杂的多变性,使得不同植物或同一物种不同基因型的根际微生物群落结构不同[4]。因此,研究植物不同基因型根际土壤的微生物结构和多样性对其抗病机制具有重要意义。【前人研究进展】研究表明每克植物的根部约含1011个微生物细胞,可以提供巨大的潜在功能,被称为寄主植株的“额外”基因组[5]。一是土壤微生物在营养元素循环、肥力提高、生态环境改善、植物生长发育、作物病虫害防治等方面具有重要作用[6−7]。二是微生物影响植物生长健康,如根际微生物群落结构与土传病害的发生有一定内在联系,某些微生物代谢产物抑制植物的生长等[8]。根际微生物在特定生态环境和植物类型存在的群落结构差异可促使植物产生特定的根际效应,根际效应直接影响着根际微生物的营养选择和富集[9]。Kwak 等研究结果表明抗青枯病番茄Hawaii 7 996 可以富集更多的黄杆菌基因组,抑制青枯菌的生长[5]。此外,小麦、水稻、西瓜、黄瓜等作物根际微生物与土传病害发生关系的研究也说明作物抗感品种间根际微生物群落结构存在差异[10−11]。因此,在农业生产中客观分析根际土壤微生物多样性以及微生物群落结构特征对研究和控制土传病害有很好的帮助。【本研究切入点】由青枯菌5 号小种引起的桑树青枯病是最主要的桑树细菌性病害之一,已成为制约我国蚕桑产业发展的重要因素[12]。由于桑树青枯病很难防治,一旦爆发会造成严重的损失,当前疫区主要利用抗性品种进行防控[13]。海南目前主要栽培叶桑树品种有抗青283×抗青10、抗青10、桂桑优62 及其他嫁接苗等[14]。前期调查发现,抗青10、抗青283×抗青10 对青枯病抗性强,较少有发病,桂桑优62 对青枯病抗性弱,在琼中地区多发病。这种差异与根际土壤微生物结构及多样性之间有何关系目前尚不清楚。【拟解决的关键问题】基于此,本研究对抗青枯病桑树种质抗青283×抗青10 和敏感种质桂桑优62,采用16S rRNA 高通量测序技术研究其根际细菌群落结构与多样性,拟找出抗、感桑树根际细菌群落的差异,为进一步研究桑树根际微生态特征、根际功能菌株的筛选和应用提供依据。
1 材料与方法
1.1 材料
抗青283×抗青10 杂交组合是湛江市蓖麻蚕科学研究所黄富等采用有性杂交、病地添菌、人工诱变等方法,于1994 年3 月17 日通过广东省农作物品种审定委员会桑桑专业组的审定,对青枯病达到中抗水平[15]。桂桑优62 是广西壮族自治区蚕业技术推广总站1995 年选育出的优良组合,于2000 年通过广西农作物品种审定委员会审定,耐高温,适于珠江流域等热带、亚热带种植[16]。桑树均为2016 年种植,树龄为3 年。
1.2 根际土壤样品的采集
样地位于海南省琼中县营根镇加钗农场品种示范基地(N19.02′29″,E109.47′22″;H: 306 m),样地 的 土 壤pH 为6.25,有 机 质39.63 g·kg−1,铵 态 氮54.60 g·kg−1,电 导 率605.00 us·cm−1,盐 分330.00 mg·L−1,总溶解固体物300.00 mg·L−1,全钾为24.77 g·kg−1,全磷为1.37 g·kg−1。2019 年7 月采集根际土壤样品,在抗青283×抗青10 和桂桑优62 种植行中随机选取4 行,每行选取5 株植株混合为一个样品,去除地表土壤后,取根际土壤样品,抗青283×抗青10 植株根际土壤样本编号为QZ2K1、QZ2K2、QZ2K3、QZ2K4,桂桑优62 植株根际土壤样本编号为QZ2G1、QZ2G2、QZ2G3、QZ2G4。所有样品置入冰盒,-80 ℃冰箱保存、备用。
1.3 土壤微生物DNA 提取
取充分混匀的土壤样品0.50 g,采用DNeasy Power Soil Kit(100)试剂盒提取根际土壤DNA。抽提DNA 浓度和纯度利用Nano Drop 2000 检测,要求A 260 /A 280 值在1.8~2.0。
1.4 基因扩增与测序
Takara 公司的Takara Ex Taq 高保真酶进行PCR,利用16S V3-V4 区域引物343F 5′- TACGGRAGGC AGCAG -3′和798R5′- AGGGTATCTAATCCT-3′扩增[17]。PCR 扩 增 体 系:25 μL 2x PremixTaq(Takara Biotechnology),引物各1 μL(10 mmol·L−1),DNA 模板3 μL(20 ng·μL−1),加dd 水 溶 解 定 容 至50 μL。PCR 反应程序:预变性:94℃ 5 min;变性:94℃30 s,退火:52℃ 30 s,延伸72℃ 30 s,30 个循环;延伸:72℃ 10 min。采样琼脂糖电泳检测,利用Qubit 2.0 DNA 检测试剂盒精确定量回收产物并进行测序。
1.5 数据分析
使用Trimmomatic(Version 0.35)软件对原始数据(raw data)质量部分剪切,拼接得到完整双端序列(paired end tags)。 使 用 QIIME 中 的 Split Libraries(Version 1.8.0)软件得到高质量序列(clean tags)。去除高质量序列中的嵌合体,得到优质序列(valid tags)。 对质控得到的优质序列使用Vsearch(Version 2.4.2)软件按照97%的相似度进行OTU 分类,以每个OTU 中丰度最大序列为该OTU 的代表序列,采用RDP Naive Bayesian Classifier分类算法进行注释。使用BioVenn 在线软件进行Venn 图绘制;使用Excel 2013 和SPSS 19.0 软件对相对丰度数据进行统计分析,绘制相对丰度柱状图;使用Qiime 软件计算观测物种数、Chao1 指数(Chao1)、香农、辛普森、谱系多样性指数(PD whole tree)和文库覆盖率(Goods-coverage)等多样性指数,稀释曲线和NMDS 图用R 软件绘制。通过LDA 对根际细菌群落进行LEFSE 分析(LDA Score 2.5)寻找抗、感青枯病桑树根际土壤细菌群落差异类群;使用PICRUSt 软件,预测已知微生物基因功能的构成。
2 结果与分析
2.1 根际细菌群落测序结果分析
细菌16S rRNA 高通量测序共获得优化序列304 272 条,平均长度431.48 bp(表1)。8 个样本序列均超过32 000 条reads,样品测序深度都超过94%,测序效果理想。
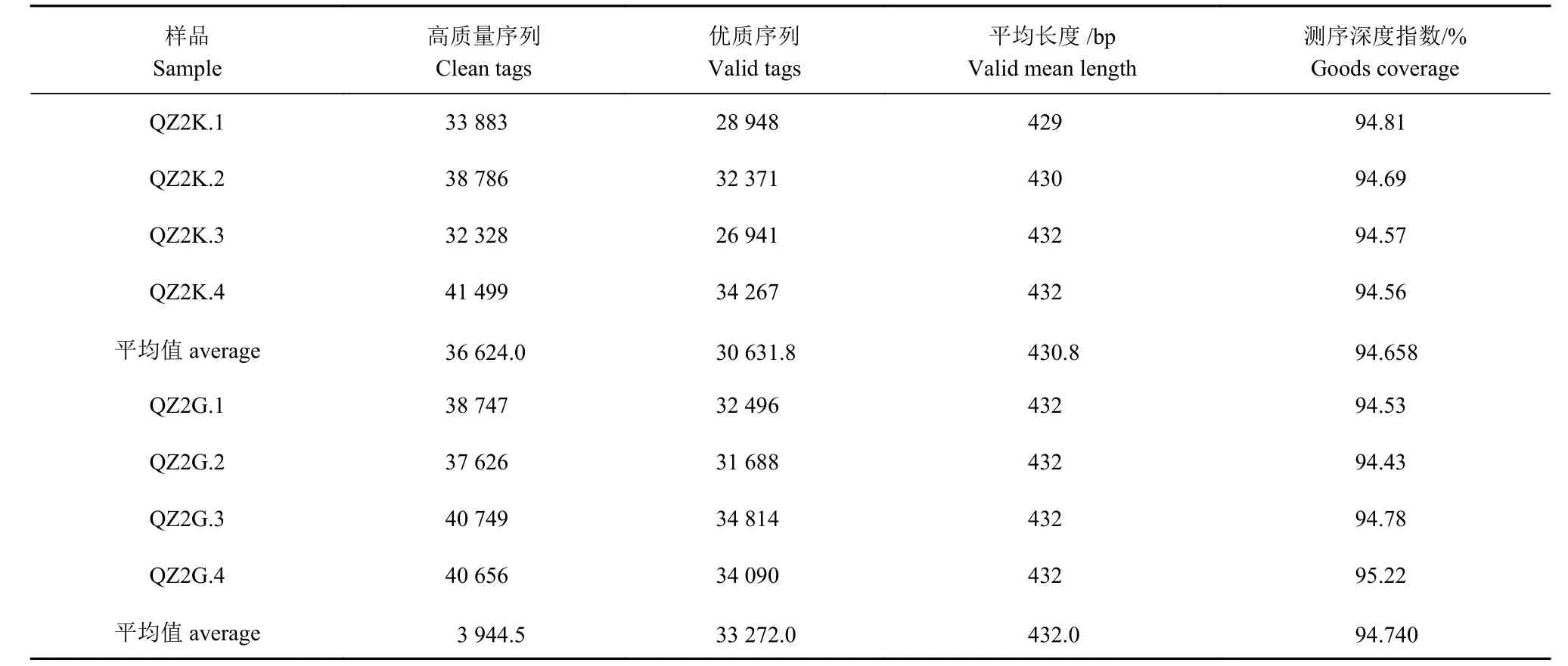
表 1 高通量测序数据概况Table 1 Overview of high-throughput sequencing data
2.2 细菌群落组成比较
抗青枯病桑树样品共鉴定出细菌28 个门、76 个纲、151 个目、230 个科、388 个属;易感青枯病桑树样品共鉴定出细菌27 个门、73 个纲、147 个目、225 个科、363 个属。根据Venn 图可知,共有OTUs数为2 089 个,其中QZ2K 特有的OTUs 为1 113 个,QZ2G 为898 个(图1)。
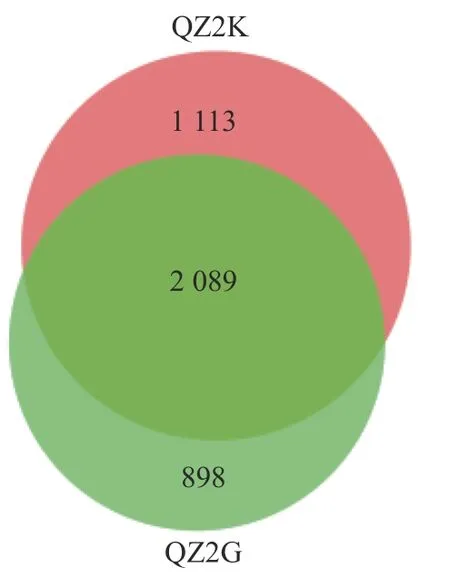
图 1 抗感青枯病桑树根际土壤细菌OTUs 韦恩图Fig. 1 Venn diagram of bacteria OTUs in rhizosphere soils at QZ2K and QZ2G
桑树根际土壤样品中细菌群落门水平上组成结构和丰度结果如图2-A 根际土壤细菌主要分布在以下15 个门:变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、 酸 杆 菌 门(Acidobacteria)、 芽单 胞 菌 门(Gemmatimonadetes)、 拟 杆 菌 门(Bacteroidetes)、厚壁菌门(Firmicutes)、硝化螺旋菌门(Nitrospirae)、匿杆菌门(Latescibacteria)、Entotheonellaeota、 Patescibacteria、 河 床 菌 门(Zixibacteria)、蓝细菌门(Cyanobacteria)、迷踪菌门(Elusimicrobia)、绿弯菌门(Chloroflexi)、罗克菌门(Rokubacteria),其中变形菌门最为丰富,QZ2K和QZ2G 土样中相对丰度分别为44.54%,46.23%,其次为放线菌门,QZ2K 和QZ2G 土样中相对丰度分别为23.01%,28.86%。其中QZ2K 根际土样中酸杆门、芽单胞菌门、厚壁菌门、硝化螺旋菌门、Patescibacteria、河床菌门、蓝细菌门、罗克菌门的含量高于QZ2G 根际土样的含量。
在属水平上如图2-B 所示,前十五的属分别为MND1、Gaiella、 硝 化 螺 菌 属 (Nitrospira)、Haliangium、链霉菌属(Streptomyces)、类诺卡氏菌属(Nocardioides)、芽胞杆菌属(Bacillus)、拟杆菌 属(Bacteroides)、Subgroup 10、Acidibacter、分支 杆 菌 属 (Mycobacterium)、 土 微 菌 属(Pedomicrobium)、Ellin6067、mle1-7、鞘氨醇单胞菌 属(Sphingomonas)。 芽 胞 杆 菌 属 在 QZ2K土样中占1.76%,而在QZ2G 土样中仅为0.41%。另外,土微菌属、拟杆菌属、mle1-7、鞘氨醇单胞菌属、类诺卡氏菌属、硝化螺菌属在QZ2K 土样中的含量也高于其在QZ2G 土样中的含量。
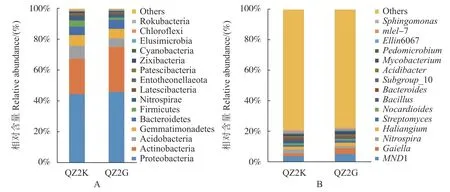
图 2 细菌群落门(A)和属(B)水平相对丰度Fig. 2 Phylum (A) and genus (B) distributions of OTUs
2.3 群落多样性分析
2.3.1 Alpha 多样性 对抗、感桑树根际土壤细菌群落进行6 种α 多样性指数计算(表2)。统计结果表明,抗感桑树根际细菌群落的各个多样性指数均无显著性差异。谱系多样性指数、Chao1 指数和观测物种数在抗青枯病桑树QZ2K 土样中高于感青枯病桑树QZ2G 土样。
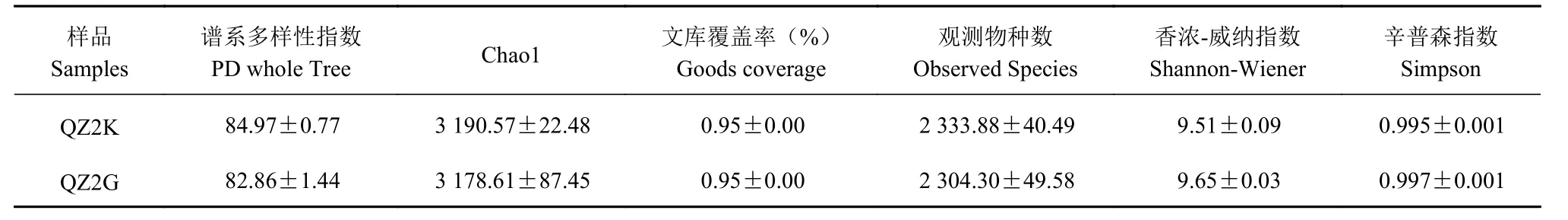
表 2 抗感青枯病桑树根际土壤细菌α 多样性指数(平均值±标准误,n=4)Table 2 Alpha diversity indices of bacteria in rhizosphere soils at QZ2K and QZ2G(Mean±SE,n=4)
2.3.2 Beta 多样性 Beta 多样性利用各样本序列间的进化关系及丰度信息来计算样本间距离,反映样本(组)间是否具有显著的微生物群落差异。基于加权UniFrac 距离的非度量多维标定法(Non - Metric Multi - Dimensional Scaling,NMDS)来研究不同基因型根际细菌群落之间的分离模式考虑了分类群丰度,对稀有的分类群更敏感。如图3 所示,在UniFrac NMDS 中,stress=0.005<0.05,说明很好地将抗青枯病桑树根际土壤QZ2K 与感青枯病桑树根际土壤QZ2G 样本区分开来,表明桑树不同基因型根际细菌群落之间存在显著的差异。
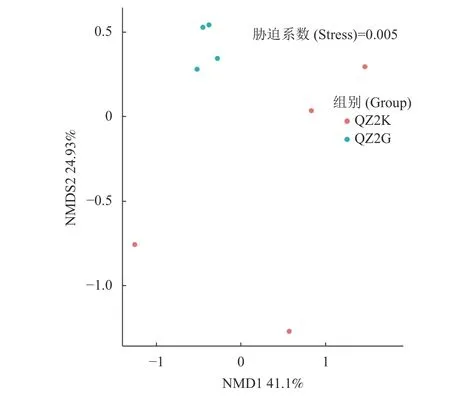
图 3 抗感青枯病桑树根际土壤细菌NMDS 分析Fig. 3 Non-MetricMulti-dimensional scaling by UUF metric
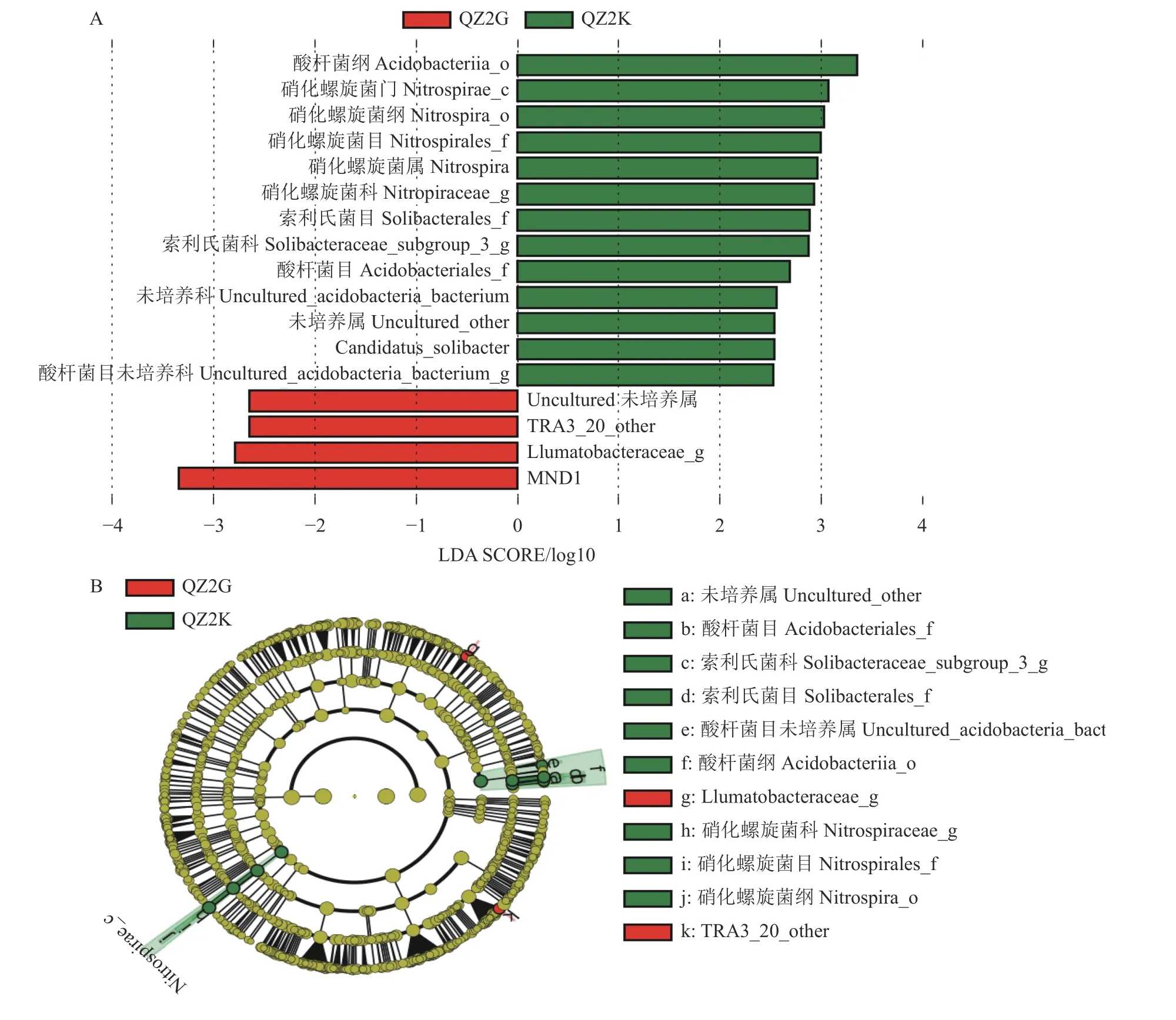
图 4 抗感青枯病桑树根际土壤细菌LEfSe 分析Fig. 4 LEfSe analysis on bacteria in rhizosphere soils at QZ2K and QZ2G
2.4 抗感青枯病桑树根际优势细菌差异分析
使用LEfSe 分析抗青283×抗青10 与桂桑优62 根际土壤细菌群落中主要差异物种(LDA Score 预设值为2.5,小于2.5 视为没有显著差异)(图4-A)。结果表明,酸杆菌纲(Acidobacteriia)、硝化螺旋菌门、硝化螺旋菌纲(Nitrospira)、硝化螺旋菌目(Nitrospirales)、硝化螺旋菌属(Nitrospira)、硝化螺旋菌科(Nitrospiraceae)、索利氏菌目、索利氏菌科(Subgroup 3)、 酸 杆 菌 目(Acidobacteriales)、Subgroup 2 未培养科未培养属、酸杆菌目未培养科、Candidatus Solibacter、Subgroup 2 未培养科这几类菌群在抗青283×抗青10 根际的相对丰度均显著高于桂桑优62 根际,抗青283×抗青10 根际酸杆菌纲LDA 值最大,桂桑优62 根际MND1 LDA 值最大,说明这两个类菌群的差异最大。从图4-B 中可以看出抗青283×抗青10 根际重要作用的细菌类群包括:硝化螺旋菌纲、酸杆菌纲、硝化螺旋菌目、索利氏菌目、酸杆菌目、硝化螺旋菌科、酸杆菌目未培养科、索利氏菌科(Subgroup 3)和Subgroup 2 未培养科;而桂桑优62 根际重要作用的细菌类群Ilumatobacteraceae 和TRA3 20 Other 两个科。
2.5 抗感青枯病桑树根际细菌群功能分析
基于16S rDNA 序列的PICRUSt 功能预测(图5),抗青283×抗青10 和桑优62 的根际细菌群落对应的COG 类目排名前10 的相同,分别为氨基酸运输与代谢(E)、转录(K)、信号转导机制(T)、能量产生与转化(C)、细胞壁/膜/包膜生物发生(M)、碳水化合物运输与代谢(G)、复制,重组和修复(Lr)、翻译,核糖体结构和生物源(J)、无机分子运输与代谢(P)、辅酶运输与代谢(H)(不包括未知功能和通用功能);两组根据Wilcoxon 算法差异COG 类目为763,仅占所有COG 类目的17.25%。
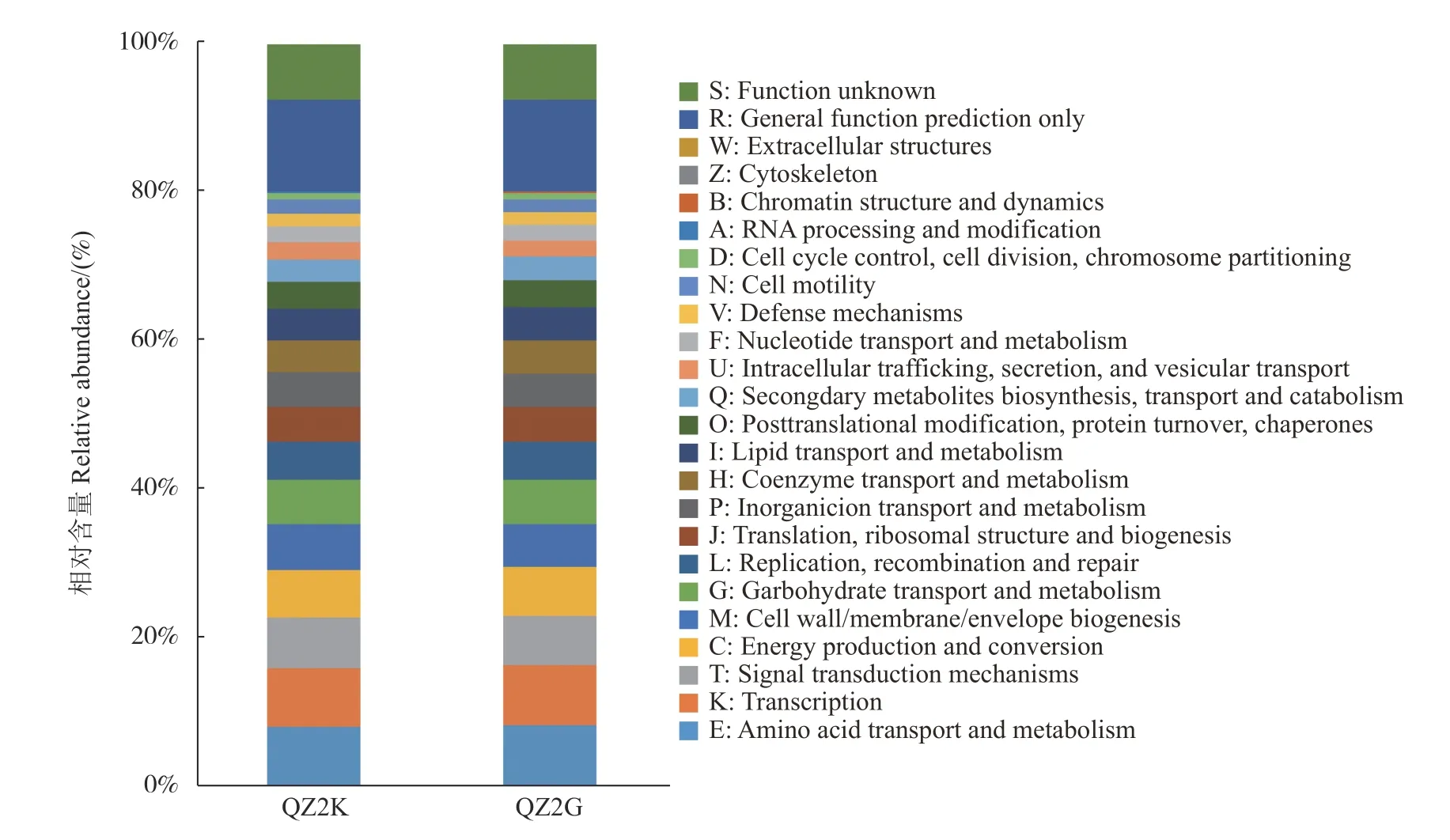
图 5 抗感青枯病桑树根际细菌预测得到的COG 相对丰度Fig. 5 Relative abundance of COG at QZ2K and QZ2G
3 讨论与结论
植物对青枯病的抗性是由数量性状基因介导的,但植物对其表型是可变的,并受环境因素影响,其中植物根际微生物在抗青枯病上具有重要作用[5,7]。本研究发现抗青283×抗青10 根际细菌在门、纲、目、科、属和OTU 分类水平均略高于桂桑优62 根际细菌群落,但对根际细菌物种多样性分析结果表明两种桑树根际细菌群落多样性差异不显著。这可能是因为两种基因型桑树种植在同一样地,管理方式一致,土壤在根系细菌组装过程中占主导地位。在门水平,变形菌门、放线菌门、酸杆菌门、芽单胞菌门、拟杆菌门、厚壁菌门等为根际土壤的优势菌门,分别占抗青283×抗青10 和桂桑优62 根际细菌总体的92.15%、94.27%。结果与邱洁等研究3 个品种桑树(粤椹大10、嘉陵30 号和红果1 号)根际细菌结构基本一致[18]。在属水平,主要分布在MND1、Gaiella、硝化螺菌属、Haliangium等15 个属,但抗、感桑树根际各属含量有差异,抗青283×抗青10 根际细菌在放线菌的类诺卡氏菌属和芽胞杆菌属均高于桂桑优62 根际细菌。邱洁等对3 种桑树根际细菌研究也发现,随着分类的细化,不同的桑树品种对细菌群落组成和分布的影响越大,在科属水平上差异更明显[18]。
进一步基于加权UniFrac 距离的非度量多维标定法Beta 多样性分析,抗青283×抗青10 和桂桑优62 的根际细菌群落可以显著区分开来。说明抗感桑树根际分别富集了不同的优势细菌菌群,可能是基因型通过植物差异生长性能、凋落物和根系分泌物等影响植物根际微生物群组装。通过LDA Score 进行LEfSe 分析,发现抗、感青枯病桑树根际重要作用的微生物类群存在差异。类诺卡氏菌属、芽胞杆菌属、鞘氨醇单胞菌属在抗性品种根际丰度也更高。研究表明类诺卡氏菌属和芽胞杆菌属分别属于放线菌和芽胞杆菌,是生防菌,通常代表土壤的健康状况对植物病害具有防病作用[19−20]。研究表明鞘脂单胞菌科(Sphingomonadaceae)在具有抑病性的烟草根际土壤中具有普遍存在的特征[18,21],鞘脂单胞菌科的某些菌株也被报道与根部固氮作用密切相关[22],因此,鞘氨醇单胞菌属也可能与植物对病原体攻击的抵抗力有关。抗青枯病桑树根际土壤富集了更多的类诺卡氏菌属、芽胞杆菌属、鞘氨醇单胞菌属,可能对其抵抗青桔病有关,这有待于研究证实。
本研究利用Illumina 高通量测序技术对抗感青枯病桑树的根际土壤样品进行了测序分析,同一地点抗、感青枯病桑树根际细菌群落多样性差异不显著,但其重要的细菌类群存在差异,抗青枯病桑树根际土壤富集了更多的类诺卡氏菌属、芽胞杆菌属、鞘氨醇单胞菌属等生防菌,这为进一步研究桑树青枯病的防治奠定了基础。有研究发现品种抗性差异可能通过根系分泌物对病原菌的化感作用差异来体现[23],根系分泌物通过刺激特定微生物的生长,富集特定微生物,在植物-微生物和微生物-微生物相互作用中发挥重要作用[24−26]。因此,为阐明抗感病桑树根际微生物差异的机制及原因,尚需进一步对抗、感青枯病桑树种质根系分泌物组分、含量差异及其对土壤微生物的影响作进一步测定分析。
猜你喜欢
音乐教育与创作(2022年1期)2022-04-26
中国土壤与肥料(2021年5期)2021-12-02
今日农业(2020年16期)2020-12-14
天然产物研究与开发(2018年3期)2018-05-07
小学生作文(中高年级适用)(2018年3期)2018-04-18
中国蔬菜(2016年8期)2017-01-15
学生天地·初中(2016年6期)2016-05-14
现代农业(2015年1期)2015-02-28
植物营养与肥料学报(2012年5期)2012-10-26
中国烟草学报(2012年5期)2012-04-12
