
丛枝菌根真菌与有机肥配施对甜玉米根际土壤氮素转化及氮循环微生物功能基因的影响
2020-12-16 03:00左明雪徐如玉袁银龙顾文杰卢钰升徐培智解开治
福建农业学报 2020年9期
左明雪,孙 杰,徐如玉,袁银龙,顾文杰,卢钰升,徐培智 ,解开治
(1. 甘肃农业大学资源与环境学院,甘肃 兰州 730070;2. 广东省农业科学院农业资源与环境研究所,广东 广州 510640)
0 引言
【研究意义】甜玉米(Zea maysL.)富含多种氨基酸、维生素、蛋白质,含糖量高,享有“水果玉米”和“保健水果玉米”的美称[1−2]。据不完全统计,我国甜玉米种植面积占世界种植面积的21%,其中广东省是我国甜玉米主产区和消费区[3]。氮素是甜玉米需要量最高、影响甜玉米生长和产量品质的必需矿质营养元素。但目前甜玉米氮肥利用率低于全国玉米氮肥利用率水平,盲目增施氮肥追求高产不仅造成资源浪费还会带来一系列的环境问题[4]。丛枝菌根真 菌(Arbuscular mycorrhizal fungi, AMF)能 与90%以上植物形成菌根共生体,并且促进宿主对氮磷钾等营养元素的吸收和利用[5]。但也有研究指出AMF只有当自身N 素被满足后才可能将多余氮素传递给宿主,增加碳源能刺激AMF 共生体内的N 吸收转运[6]。有机无机肥配施作为氮肥高效管理的关键施肥措施,通过增加土壤有机碳含量为氮循环微生物提供碳源,来间接或直接调控土壤氮素循环关键环节,从而提高土壤氮肥利用效率以及作物产量,减少化学氮肥投入和削减氮素损失对环境的负面影响。现有研究表明,有机无机肥配施能实现作物高产稳产,增强土壤氮素有效性,提高氮素利用率,具有改良土壤、保水保肥、减肥增产等作用[7−8]。研究AMF 配施有机肥对甜玉米根际土壤氮素转化以及氮循环微生物功能基因的影响有助于为甜玉米增产、氮素减施提供理论依据。【前人研究进展】有机肥含有大量有机碳源,能促使微生物将更多的铵态氮同化进入到土壤活性有机碳库[9],提高氮素矿化同化速率,通过后续微生物作用转变为植物有效氮,有效固氮降低氮素损失[10−11]。有机肥通过调节土壤碳氮比能使土壤中的异氧微生物在吸收利用NH4+-N的同时也进行NO3−-N 同化,激发自养硝化和异氧硝化,为反硝化提供底物[12−14]。Leininger 等[15]研究表明,有机无机肥配施提供的基质、养分有利于氨氧化细菌(Ammonia-oxidizing bacteria, AOB)以及氨氧化古菌(Archaea-oxidizing ammonia, AOA)的生长,显著提高土壤中AOB 的丰度。而AMF 在玉米减肥增产方面具有重要的作用。赵乾旭等[16]通过盆栽试验证明丛植菌根真菌能与玉米形成良好共生体,增加作物含氮量和生物量,提高19.51%的氮吸收利用率;刘铭铭等[17]通过盆栽试验表明增施AMF能提高玉米27.8%的氮肥利用率。李芳等[18]通过大田试验,菌根化育苗指出接菌植株根系的菌根侵染强度和丛枝丰度均显著高于不接菌植株,但焦点着力于作物对于磷的吸收利用。【本研究切入点】目前对于AMF 促进化肥减量主要局限于盆栽试验,并且大部分学者关注磷肥减施。而AMF 配施有机肥的研究也主要集中在土壤微生物群落多样性差异以及具体的某一个氮循环转化环节,针对有机无机肥配施后土壤氮素转换以及氮循环微生物功能基因的研究甚少。【拟解决的关键问题】本研究采用大田区组设计,利用功能基因芯片(GeoChip 5.0)技术,研究AMF 配施有机肥对甜玉米根际土壤氮素转化以及氮循环微生物功能基因的影响,探索其对甜玉米-土壤氮循环的微生物学过程,为氮肥利用率提高、化学氮肥减施和氮肥增效等提供技术支撑。
1 材料与方法
1.1 试验地概况
本次定位试验在广东省惠州市惠阳区农业技术推广中心试验基地(N23°8′46.19″, E113°20′47.84″)进行。于2018 年春季开始,一年春秋两季栽培,结束于2018 年秋季。试验地属亚热带季风气候区,土壤类型为砖红壤。试验初始基本地力养分指标为有机质含量22.8 g·kg−1,碱解氮191.54 mg·kg−1,速效磷49.56 mg·kg−1,速效钾149.41 mg·kg−1,pH 值5.18。
1.2 试验材料
供试甜玉米(Zea MaysL.)品种为广东省农业科学院作物研究所提供的粤甜28 号。供试菌种为购自北京市农林科学院植物营养与资源研究所的变形球囊霉菌(Glomuse versiforme, BGC GD01C)。其菌剂本身含有被侵染的玉米植物根段、菌根真菌孢子及根外菌丝的根际沙土混合物,经检测,其孢子密度为38 个·g−1。参照毕银丽等[19]的室内盆栽砂土扩繁方式进行扩繁。基质采用风化煤∶砂∶蛭石∶珍珠岩=1∶1∶1∶1 组合,经过高压灭菌(121℃,30 min)后备用。采用10% H2O2对甜玉米种子进行10 min 的消毒后洗净置于培养箱28℃催芽备用。将2.0 kg 灭菌基质放入盆钵(75%酒精消毒)中,浇水至土壤持水量75%后将15 g 菌剂平铺,放入6 粒催芽后的甜玉米种子,最后用0.5 g 基质覆盖住种子;出苗一周后每盆定苗4 株,采用自然光照、空调控温的方式,白天控温(25±2)℃,夜间(19±2)℃,相对湿度保持在65%。在播种后45 d 每盆分别补施100 mL Hoagland 营养液,最后于甜玉米收获期,采集土壤并测定孢子含量,作为大田试验备用。
1.3 试验方法
1.3.1 试验处理 根据当地高产栽培方式优化最佳施肥量为:氮肥(N)330 kg·hm−2、磷肥(以P2O5计)150 kg·hm−2、钾肥(以K2O 计)225 kg·hm−2。试验共设7 个处理(表1),每个处理3 个重复,各小区面积67.5 m2(4.5 m×15 m),随机区组排列。各处理磷、钾肥施用量相同,施磷肥(以P2O5计)150 kg·hm−2、钾肥(以K2O 计)225 kg·hm−2,磷肥品种为过磷酸钙(P2O512%),钾肥品种为硫酸钾(K2O 50%)。氮肥使用尿素(N 46%),在基肥、苗期、拔节期分别以2∶3∶5 的比例施入,有机肥的主要养分为N 2%、P 1.5%、K 1.5%,有机肥和磷肥一次性基施,钾肥在基肥和拔节期分别以5∶5 的比例施入。所有基肥按试验设计施用量起垄撒施旋耕。甜玉米幼苗移栽时每棵定殖增施7 g 变形球囊霉(F. versiforme)菌剂,种植密度为45 000 株·hm−2,起垄种植。其他的田间管理按照当地常规管理。试验于2019 年9 月18 日移苗,12 月10 日收获。
1.3.2 样品采集方法 供试土壤、植株、根系样品均于2019 年秋季甜玉米成熟期(12 月10 日)采集。供试植株(根系)样品选取各处理中10 株长势均匀的植株整株挖出(避免损坏根系),地上部分称重、105 ℃下杀青30 min、75 ℃烘至恒重计算干物质重量。挑选不同代表性部位的烘干样品粉碎后测定地上部分全氮含量。地下部分(甜玉米根系)用清水洗净后分为两份,一份用于侵染率和侵染强度的测定,一份置于−80 ℃冰箱保存,用于测定氮代谢相关酶活性。按照Zhao 等[20]的方法采集根际土,均匀混合后分为两份,一份2 000 g 室温保存用于常规理化性质以及孢子量测定;一份500 g 放入无菌密封袋后用冰盒带回实验室−80℃保存,用于基因芯片检测。
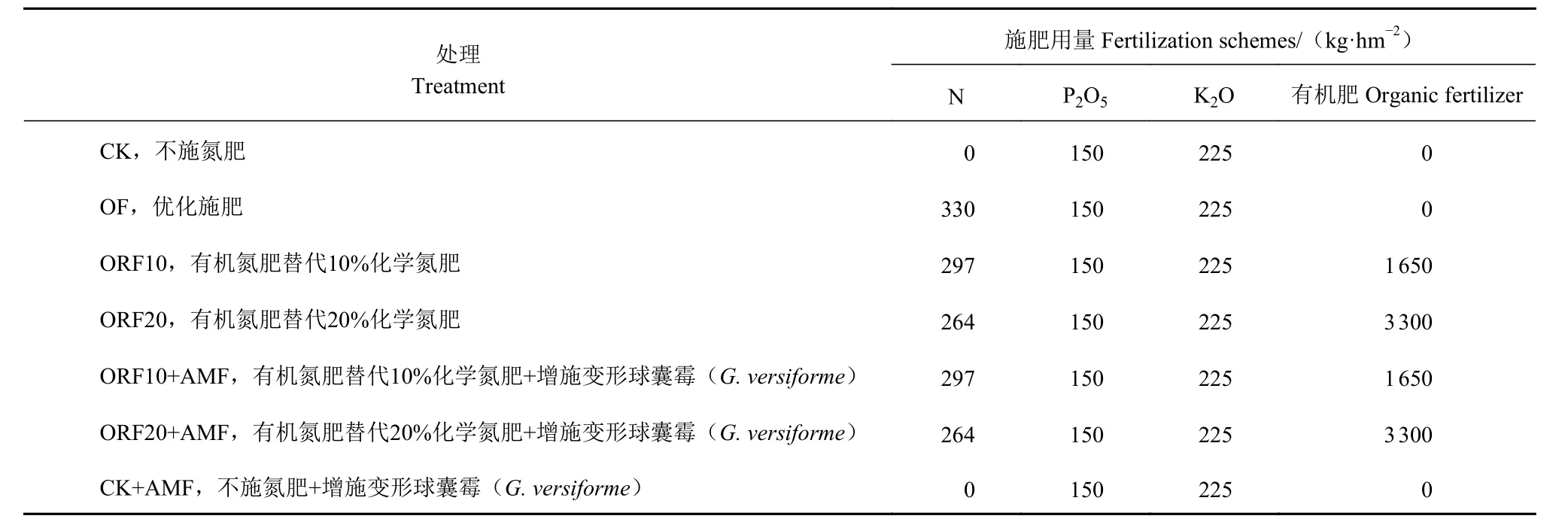
表 1 不同处理间施肥方案Table 1 Fertilization treatments
1.3.3 酶活性、土壤理化性质、侵染率和孢子量测定
硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)和谷氨酸合酶(GOGAT)采用北京索莱宝科技有限公司试剂盒进行检测[21]。常规理化性质等环境因子按照鲍士旦土壤农化分析方法测定[22]。土壤呼吸采用室内密闭培养法进行测定[23];采用氯化钾浸提流动注射分析法对硝态氮和铵态氮进行测定[23]。采用台盼蓝染色法进行染色、脱色、制片、镜检[24]对菌根侵染率和强度进行测定。土壤孢子量测定采用湿筛倾注蔗糖离心法[25]。
1.3.4 土壤氮利用率和侵染统计 氮素利用效率相关参数的计算方法[26−27]:
氮肥农学效率(NAE,kg 籽粒/kgN)=(施氮区产量−对照产量)/施氮量
氮肥偏生产力(PFP,kg 籽粒/kgN)=施肥区鲜苞产量/施氮量
氮肥吸收利用率(NRE,%)=(施氮区地上部吸氮量−对照区地上部吸氮量)×100/施氮量
氮素生理利用率(NPE,kg 籽粒/kgN)=(施氮区鲜苞产量−对照区鲜苞产量)/(施氮区地上部吸氮量−对照区地上部吸氮量)
根系侵染率以及侵染强度的计算方法[28]:
根系中的菌根侵染率/%=有菌根根段数/总根段数×100
根系中的菌根侵染强度/%=(95×侵染率90%以上根段数+70×侵染率50%至90%的根段数+30×侵染率10%至50%的根段数+5×侵染率10%以下1%以上根段数+侵染率1%以下根段数)/总根段数×100
1.3.5 根际土微生物功能基因芯片 采用土壤DNA提取试剂盒(MinkaGene Soil DNA Kit)对甜玉米根际土壤样品进行总DNA 的提取,利用NanoDrop One检测DNA 的完整性、纯度和浓度。采用Cy3 染料和DNA 聚合酶I 对纯化后的DNA 进行标记,采用QIAquick PCR Purification Kit(Qiagen, Valencia, CA,USA)纯化标记的DNA,置于Labconco Centrivap浓缩器中50 ℃干燥45 min,DNA 稀释至相同浓度后在95 ℃下孵育5 min,并在42 ℃下进行杂交16 h,将标记的DNA 置于阵列上,然后在杂交站(BioMicro Systems, Inc., Salt Lake City, UT)将阵列预热至42 ℃(5 min),最后用NimbleGen MS200 扫描仪进行预处理和信号强度的标准化。
1.4 数据统计与分析
图表制作和统计分析采用Excel 2010 和SPSS 20.0,施菌处理和有机无机配施水平及其交互作用的差异显著性采用双因素方差分析(Two-way ANOVA)进行检验,不同处理之间的差异显著性采用Duncan’s多重检验法进行检验(P<0.05)。除趋势对应分析和不相似性分析使用R 软件(R-3.2.2)进行分析绘图;RDA 分析和绘图采用QIIME2 和ggplot2 包。
2 结果与分析
2.1 根际土壤氮素转化及利用率分析
2.1.1 不同处理AMF 在甜玉米根际土壤中的定殖与侵染 如表2 所示,每个处理的甜玉米根系都有不同程度的AMF 侵染。单施菌肥CK+AMF 处理侵染率、侵染强度最高,对照CK 处理最低。CK+AMF处理侵染率、侵染强度分别达81.0%和45.34%。变形球囊霉配施有机肥处理ORF20+AMF 处理孢子密度和侵染强度与OF、ORF10、ORF20、ORF10+AMF处理之间达到显著水平(P<0.05),但侵染率无显著差异(P>0.05)。增施变形球囊霉配施有机肥处理 组(CK+AMF、 ORF10+AMF、 ORF20+AMF)中的AMF 孢子密度、侵染强度显著高于相应未施菌处理组(CK、OF、ORF10、ORF20),增施变形球囊霉能极显著影响孢子密度和侵染强度(P<0.001),且与有机无机肥配施存在正交互效应(P<0.05),有机无机配施组间无显著差异。说明增施AMF 有助于提高根际土壤孢子量和侵染率,增加侵染强度。
2.1.2 不同处理甜玉米根际土壤氮素利用率 各处理间氮素养分吸收利用存在一定差异(表3)。变形球囊霉配施有机肥处理组(ORF20+AMF、ORF10+AMF)的氮肥农学效率(NAE)和氮肥偏生产力(PFP)显著高于对应未施菌处理组(ORF20、ORF10),氮肥吸收利用率表现为变形球囊霉配施处理ORF20+AMF 处理显著高于ORF20 处理,氮素生理利用率表现为变形球囊霉配施有机肥处理组(ORF20+AMF、ORF10+AMF、CK+AMF)大于对应不施菌处理组(ORF20、ORF10、CK),组间差异不显著。综合来看,ORF20+AMF 氮肥利用率最高,显著高于包括优化施肥的其他6 个处理。增施变形球囊霉和有机无机配施均极显著影响氮肥农学效率、氮肥偏生产力,但二者无显著的交互效应。
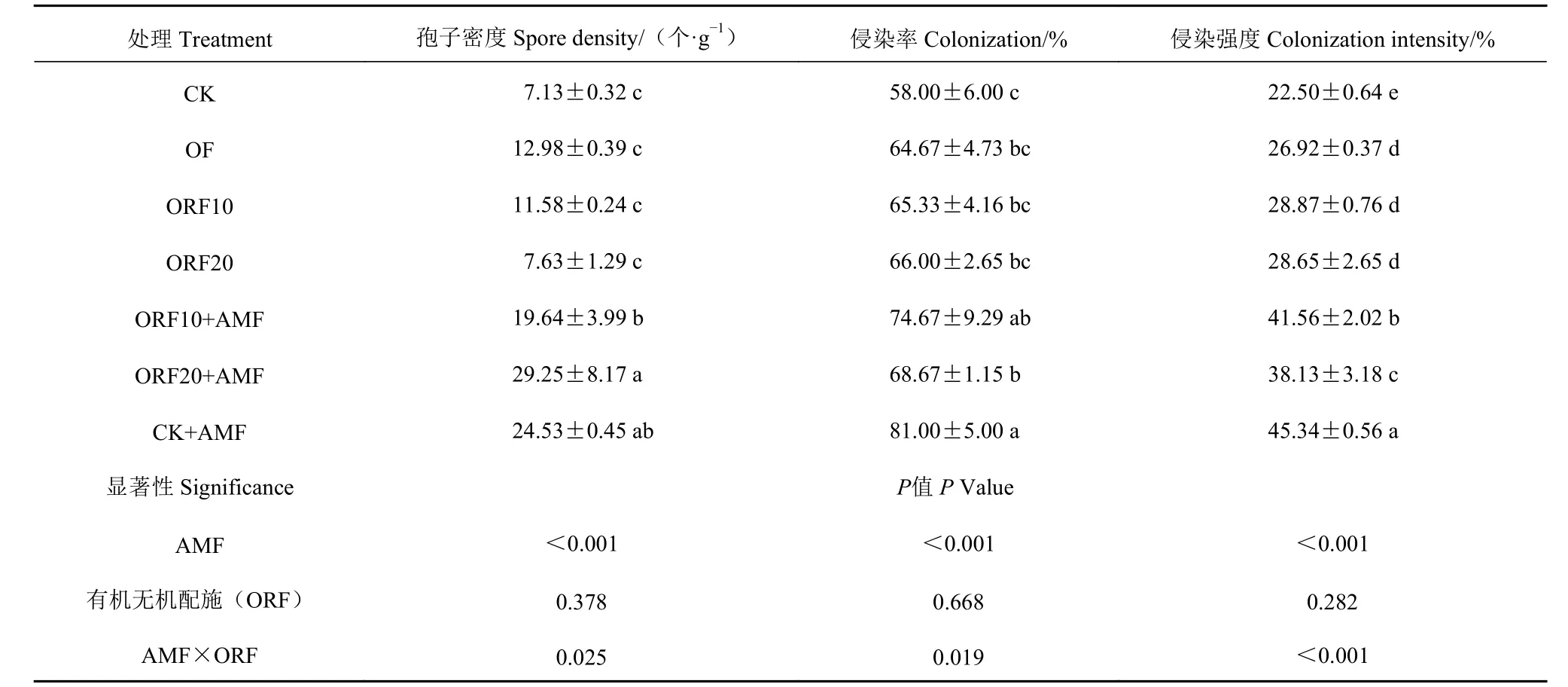
表 2 不同施肥处理对甜玉米孢子量、侵染率、侵染密度的影响Table 2 Spore quantity, infection rate, and infection density on sweet corn under treatments
2.1.3 不同处理对甜玉米根际酶活性影响分析 各处理与氮代谢相关的甜玉米根系硝酸还原酶(NR)、谷氨酸合酶(GOGAT)和谷氨酰胺合成酶(GS)活性存在差异(表4)。变形球囊霉配施有机肥处理组(ORF10+AMF、ORF20+AMF)硝酸还原酶(NR)活性显著低于对应未施菌处理组(ORF10、ORF20)。谷氨酸合酶(GOGAT)活性表现为变形球囊霉配施有机肥处理(ORF20+AMF、CK+AMF)显著低于未施菌处理组(ORF20、CK)处理。变形球囊霉配施有机肥ORF20+AMF 处理的谷氨酰胺合成酶(GS)显著高于未施菌处理ORF20。有机无机配施以及增施AMF 之间的交互作用表明,有机无机配施与增施变形球囊霉对硝酸还原酶(NR)、谷氨酸合酶(GOGAT)和谷氨酰胺合成酶(GS)活性具有极显著的正交互效应(P<0.001)。

表 3 不同施肥处理甜玉米氮利用率Table 3 Nutilization efficiency of sweet corn under treatments

表 4 不同施肥处理甜玉米根内系氮代谢相关的酶活性Table 4 Activity of microbial N metabolizing enzymes in sweet corn under treatments
2.2 根际土微生物氮循环功能基因群落差异
7 个处理21 个样品中共检测到7 个氮循环功能亚类,包括固氮、氨化、硝化、反硝化、同化氮还原、异化氮还原和氨氧化作用(图1)。7 个功能亚类强度表现为:反硝化>同化氮还原>氨化>固氮>异化氮还原>硝化>氨氧化。除氨氧化作用各处理间差异不显著外,其他6 个氮循环亚类功能强度在各处理间均有显著差异(P<0.05)。变形球囊霉配施有机肥处理组(CK+AMF、ORF10+AMF、ORF20+AMF)的6 个氮循环亚类功能强度整体上显著高于对应不施菌处理(CK、ORF10、ORF20),并且ORF20+AMF 处理的7 个氮循环亚类功能强度均为最高。
除趋势图对应分析(DCA)结果表明(图2),7 个处理甜玉米根际土壤氮循环微生物功能基因组间差异大于组内差异。DCA 排序图中,7 个处理分异为3 个集中区域,各区域内的各处理在微生物氮循环功能上较为相近。其中,CK、OF 和ORF10 处理分异到一个区域,ORF20 和ORF10+AMF 处理分异为一个区域,CK+AMF 和ORF20+AMF 处理分异为一个区域。进一步进行各处理间微生物氮循环功能基因不相似性检验,结果表明微生物功能基因结构在各处理间具有极显著的差异性(P<0.001,表5)。
2.3 根际土微生物氮循环关键功能基因特征
所有样品共检测到2 752 个氮循环基因,其中固氮基因439 个、氨化基因426 个、硝化基因82 个、反硝化基因1 263 个、同化氮还原285 个、异化氮还原189 个、氨氧化基因9 个。
各处理中土壤微生物氮循环基因强度组间差异如图3 所示。同化氮还原基因共有NiR、narB、nasA、nirA和nirB等5 个基因,其中相对强度最大的是nasA;异化氮还原基因共有napA、nrfA两个基因;固氮基因中有编码固氮酶的nifH基因;氨化作用中的ureC、glnA、gdh基因,相对强度表现为glnA<gdh<ureC;硝化作用中的氨单加氧酶amoA、hao编码基因;反硝化作用中的conrB、narG、nirK、norB、nosZ、nirS等7 个基因,其中narG相对强度最高,其次是nosZ、nirS、nirK,conrB相对强度最低。变形球囊霉配施有机肥处理ORF10+AMF 的反硝化基因cnorB、norB、nirS与对应不施菌处理间差异不显著。但变形球囊霉配施有机肥处理组(CK+AMF、ORF20+AMF)处理的反硝化基因cnorB、norB、nirS显著高于对应不施菌处理组(CK、ORF20);变形球囊霉配施有机肥处理组(ORF10+AMF、ORF20+AMF)硝化hao基因相对强度与对应未施菌处理组(ORF10、ORF20)之间差异不显著,其他同化氮还原基因、异化氮还原基因、氮固定基因、氨化作用基因均表现为变形球囊霉配施有机肥处理组(CK+AMF、ORF10+AMF、ORF20+AMF)显著高于对应未施菌处理组(CK、ORF10、ORF20)。

图 1 不同处理亚类氮循环功能基因标准化相对信号强度Fig. 1 Relative signal intensity of normalized microbial N-cycling genes under treatments

图 2 各处理土壤微生物氮代谢功能基因结构DCA 排序图Fig. 2 DCA ordination diagram on structure of N-metabolizinggenes in soil microorganisms

表 5 不同施肥处理间微生物氮循环功能基因不相似性检验Table 5 Dissimilarity test on microbial N-cycling genes under treatments
2.4 不同处理间氮代谢功能基因丰度的变化
各处理中7 个亚类功能中的具体基因在整个氮循环中的变化如图4 所示,CK+AMF 处理除同化氮还原基因nasA、NiR丰度比CK 处理呈显著增加,其余各个亚类功能基因均是极显著增加(P<0.01)。较ORF10 处理,变形球囊霉配施有机肥处理ORF10+AMF 的氨化基因gdh、同化氮还原基因nasA、nirK、nirS、显著降低了14.12%、12.50%、11.61%和7.14%。较ORF20 处理,变形球囊霉配施有机肥处理ORF20+AMF 除氨氧化基因hzo 相对丰度降低了20%,其他亚类功能均有上升或显著上升。变形球囊霉配施有机肥处理ORF20+AMF 处理反硝化基因nirK、nirS,以及异化氮还原基因napA基因显著高于OF 处理。
2.5 环境因子对微生物群落氮循环基因的影响
利用氮循环功能基因强度与土壤环境因子数据矩阵进行冗余分析(RDA)。本文共挑选了11 个环境因子,分别是NR、NiR、GS、GOGAT、pH、硝态氮(NN)、全氮(TN)、碱解氮(AN)、土壤呼吸(Soil R)、孢子量(MC)和有机碳(OM)。结果如图5 所示,图5(a)第一排序轴和第二排序轴分别解释了74.1%和11.9%,图5(b)第一排序轴和第二排序轴分别解释了74.1%和11.9%,都较好地解释了氮转化基因强度与根际土壤理化性质间的关系。不同环境因子对基因标准化强度的影响不同。在不施变形球囊霉处理中,固氮基因nifH、氨化作用基因(gdh、glnA、ureC)、硝化作用基因(amoA)、反硝化作用基因(narG、nirk、norB、nosZ、nirS)、同化氮还原基因(NiR、narB、nasA、nirA、nirB)、异化氮还原基因(napA、nrfA)、氨氧化基因(hzo)与 土 壤 呼 吸 强 度(Soil R)、 GS 活 性、NiR 活性、pH 呈正相关关系,与其他理化指标呈负相关关系;硝化作用基因(hao)、反硝化作用基因(cnorB)与 土 壤 呼 吸 强 度(Soil R)、GS 活 性、NiR 活性、pH、孢子量(MC)、速效氮含量(AN)呈正相关关系,与其他理化指标呈负相关关系。在增施变形球囊霉处理中,固氮基因(nifH)、氨化作用 基 因(gdh、ureC)、 硝 化 作 用 基 因(amoA、hao)、反硝化作用基因(cnorB、narG、nirk、norB、nosZ、nirS)、同化氮还原基因(narB、nasA、nirA、nirB)、异化氮还原基因(napA、nrfA)、氨氧化基因(hzo)与NiR 活性、侵染强度(IM)呈正相关关系,与其他理化指标呈负相关关系;氨化作用基因(glnA)、同化氮还原基因(NiR)与GS 活性、全氮含量(TN)、碱解氮(AN)、亚硝酸还原酶(NiR)活性、侵染强度(IM)呈正相关关系,与其他理化指标呈负相关关系。

图 3 不同施肥处理氮循环关键功能基因标准化相对信号强度Fig. 3 Normalized relative signal intensity of microbial N-cycling genes under treatments

注:括号中指示了根际土N 基因丰度的百分比变化。以增施变形球囊霉处理为准,基因丰度增加用红色标记;降低用绿色标记;其中*、**、***分别表示在0.05、0.01、0.001 水平存在显著差异。此处使用的GeoChip 版本没有将灰色基因作为目标,也没有发现或未发现未注释。Note: The percentage change of N gene abundance in rhizosphere soil is indicated in parentheses. Subject to the treatment of increased application of cystic mildew, the gene abundance is increased and marked with red; the decrease is marked with green; the *, **, *** indicate significant difference at 0.05, 0.01, 0.001 level, respectively. The version of GeoChip used here does not target the gray gene, nor has it been found or found. Found uncommented.

图 5 氮循环功能基因和理化性质的RDA 分析Fig. 5 RDA analysis on microbial N-cycling genes and physicochemical properties of soil under treatments
3 讨论
3.1 增施AMF 对根际土壤氮素转化的影响
近年来,由于不合理施用氮肥使得农业生产中投入成本增加的同时肥料利用率降低。梁元振等[29]研究发现有机无机肥配施能有效提高氮素利用率,并且氮素利用率达40%以上。薛玉晨等[7]研究发现有机肥的施入能有效促进煤矿复垦土壤氮素矿化,从而提高土壤氮素有效性。何跃军等[30]研究表明接种AMF 能显著提高宿主根系侵染率,使光皮树的大田侵染率达61%。裘浪等[31]的研究表明接菌后的玉米大田侵染强度可达30%。上述研究与本研究结果一致,增施变形球囊霉处理组的侵染率、侵染强度以及孢子量均显著大于未加菌处理组(P<0.05)。赵乾旭等[16]和刘铭铭等[17]通过盆栽试验分别得出增施AMF能提高27.8%和19.51%的氮肥吸收利用率。本研究与上述学者具有相似性,增施变形球囊霉处理ORF10+AMF 和ORF20+AMF 处理氮肥吸收利用率分别达36.39%和44.75%,比对应不加菌处理分别提高5.17%和11.11%,说明增施变形球囊霉配施有机肥能有效提高氮素利用率。氮素利用率总体规律为:增施变形球囊霉处理组(ORF10+AMF、ORF20+AMF)>OF处理>有机无机配施处理组(ORF10、ORF20)。其 中ORF20+AMF 效 果 最佳,相对于优化施肥OF 处理,氮肥农学效率、氮肥偏生产力、氮肥吸收利用率和氮肥生理利用率分别提高了31.15%、28.08%、6.95%和10.41%。
硝酸还原酶(NR)是植株硝态氮还原同化中的第一个酶,也称为限速酶,其活性大小直接影响植株氮代谢强弱,而谷氨酰胺合成酶(GS)处于氮代谢的中心位置,是将无机氮转化成有机氮过程中的关键酶。Tian 等[32]研究表明AMF 是一种丛枝菌根真菌体内存在和植物一样的GS-GOGAT 氮代谢途径,在这一代谢过程中能促进宿主对土壤氮素的吸收利用。并且Hawkins 等[33]研究表明增施AMF 能提高宿主植物硝酸还原酶活性,这与本研究结果一致。本研究发现增施变形球囊霉能显著增加甜玉米根系谷氨酰胺合成酶(GS)和硝酸还原酶(NR)活性,从而增强甜玉米对氮素的吸收利用和转化,且增施变形球囊霉与有机无机配施具有极显著的正交互效应(P<0.01)。Hodge 等[34]研究也表明增施AMF 能提高宿主氮同化能力,通过大量根外菌丝网络,扩大根系吸收营养面积,尤其是增强土壤N 库吸收能力,在满足自身生长后能更高效帮助宿主吸收转运氮素。徐明岗等[35]和张丽等[36]通过对有机无机肥配施下的稻田氮素转化与利用表明有机无机配施含有一定的碳源,并且供氮缓慢而持久。在本试验中有机无机配施能为共生体提供一定的碳源以及有机无机氮。共生体在满足自身生长后成为宿主的一个重要N 库,能增强宿主氮同化能力,提高宿主氮素利用率,最终达到减肥增效的作用。
3.2 增施AMF 对根际土壤氮循环微生物功能基因的影响
微生物功能基因芯片主要获取大量微生物群落参与生物化学过程的可靠信息,尤其是微生物功能基因。从DCA 排序图以及不相似检验可知,增施变形球囊霉能影响氮循环代谢功能基因。本研究中7 个处理21 个样品共检测出7 个亚类氮循环功能,分别是反硝化、同化氮还原、氨化、固氮、异化氮还原、硝化、氨氧化作用。7 个功能亚类功能强度表现为:反硝化>同化氮还原>氨化>固氮>异化氮还原>硝化>氨氧化。除氨氧化作用组间差异不显著外,增施变形球囊霉配施有机肥处理在其他6 个氮循环亚类功能强度表现为ORF10≈ORF20<ORF10+AMF<ORF20+AMF 处理。储成等[37]通过32 年定点试验指出有机质的提升能够显著增加amoA基因以及反硝化基因narG和nirS,提升氨化作用基因glnA和gdh丰度。但在本研究中有机肥替代无机肥后并没有显著上升,其原因可能是由于施肥年限较短所致。但增施变形球囊霉有助于提高除氨氧化功能的其他6 个氮循环亚类功能强度。氮代谢基因的强度和丰度在一定程度上能够表明氮代谢的强度,微生物群落的丰度在一定程度上也能反映基因的表达强度。Veresoglou 等[38]指出丛枝菌根真菌能够显著改变菌根固氮基因群落丰度,从而改变固氮生物过程。即丛枝菌根真菌具有固氮作用以及有助于宿主吸收营养元素。本研究结果从基因层面直接印证了这一点,施菌有助于增加氮循环亚类功能的整体强度。增施变形球囊霉对于氨氧化基因的影响与Cavagnaro等[39]在田间原位条件下番茄进行AMF 接种研究结果一致,接种AMF 不会显著改变土壤中氨氧化细菌的丰度。而Veresoglou 等[38]通过盆栽研究结果指出,增施AMF 易造成土壤中氨氧化菌丰度降低。Veresoglou等[13,38]通过对玉米接种AMF 结果表明反硝化细菌群落降低,显著降低了土壤中AOA和AOB 的丰度,容易造成硝化速率下滑,并且还指出接种AMF 容易改变根际土壤nirK基因的群落组成,从而影响反硝化过程。变形球囊霉配施有机肥处理组(ORF10+AMF、ORF20+AMF)硝化作用hao基因相对强度与对应未施菌处理组(ORF10、ORF20)之间差异不显著,但增施变形球囊霉能显著增加反硝化narG、nirK、nosZ、nirS的相对丰度。因此接种AMF 对于土壤氨氧化微生物功能基因并没有得出一致结论,造成结果差异的原因可能是大田中微生物系统比盆栽更复杂,盆栽试验易于控制变量。
从整体氮循环关键功能基因丰度变化可知,CK+AMF 处理相较于CK 处理各个氮循环关键功能基因均有显著或极显著提升,增加幅度最大。这主要是由于AMF 在土壤氮素匮乏时更易与宿主形成菌根共生体,对周围氮素的吸收转运作用更强[40−41],AMF 会吸收有机氮以提供自身和宿主生长[42],增施变形球囊霉(摩西管柄囊霉)能改善植物的氮素营养状况[43]。但从实际应用考虑,ORF20+AMF 处理是适于推广的化学氮肥减施增效方案,其氮循环功能基因丰度显著高于ORF10+AMF。这可能是由于AMF自身生长需要一定的营养物质,而接种后根系分泌的有机碳化合物降低使得反硝化细菌数量降低[44]。ORF20+AMF 处理较高的有机质含量能同时满足反硝化细菌以及丛枝菌根自身生长。并且在ORF20+AMF处理中氮循环功能基因丰度高于OF 处理。所以,就甜玉米生产而言,ORF20+AMF 施肥处理比OF 处理能更好地驱动土壤氮素循环向植物氮高效利用的途径转化。
增施变形球囊霉和未施菌处理均解释了86.0%的氮循环功能基因与环境因子的关系,说明所选的环境因子能较好解释氮循环功能基因与环境因子间的相关程度。增施变形球囊霉处理组的氮循环功能基因主要受NiR 活性、侵染强度(IM)、GS 活性、全氮含量(TN)、碱解氮(AN)的影响;而未施菌处理组主要受土壤呼吸强度(Soil R)、GS 活性、NiR 活性、pH 的影响。储成等[37]学者提出pH 和有机碳是影响氮循环功能基因的最主要因子。罗蓉[45]研究也表明有机碳能显著影响氮循环细菌群落结构。本研究表明增施变形球囊霉能够提高甜玉米的侵染强度,并且能显著影响氮循环功能基因。
4 结论
(1)变形球囊霉能有效提高甜玉米根际侵染率,增施变形球囊霉处理组的侵染率强度以及孢子量均显著大于未加菌处理组。
(2)变形球囊霉配施有机肥可显著调增同化氮还原基因(NiR、narB、nasA、nirA、nirB)、异化氮还原基因(napA、nrfA)、氮固定基因(nifH)、氨化作用基因(ureC)相对强度,驱动土壤氮素循环向植物氮高效利用的途径转化,提高氮肥农学效率(NAE)、氮肥偏生产力(PFP)、氮肥吸收利用率(NRE),变形球囊霉配施有机肥可作为一种化学氮肥减施增效的模式进行推广应用。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
今日农业(2021年12期)2021-11-28
中国瓜菜(2020年8期)2020-09-26
农民致富之友(2017年23期)2018-01-02
中国果菜(2016年9期)2016-03-01
中国烟草学报(2012年5期)2012-04-12
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年3期)2011-10-24
