
抗鲤疱疹Ⅱ型病毒卵黄抗体制备及其功能鉴定
2021-02-01 02:51寇海燕郭培红田丹阳张婷婷贡成良孙丙耀宋学宏
淡水渔业 2021年1期
寇海燕,郭培红,田丹阳,何 稳,李 坤,张婷婷,贡成良,孙丙耀,宋学宏
(1.苏州大学医学部基础医学与生物科学学院,江苏苏州 215123; 2.昆山贝瑞康生物科技有限公司,江苏昆山 215345)
异育银鲫(Carassiusauratusgibelio)是我国广泛养殖的优质杂交鲫,具有抗逆性强、肉质肥美、繁殖速度快等优点。但是,由于受鲤疱疹病毒Ⅱ型(cyprinid herpesvirus 2,CyHV-2)感染的影响,养殖异育银鲫爆发鳃出血病,病鱼的鳃、体表多处出血,尾鳍末端发黑,脾脏、肾脏肿大等症状,造成大量死亡[1],给鲫鱼养殖业带来了严重的经济损失。为应对CyHV-2带来的危害,研究者采取了多种方式来控制疾病的爆发。在饲料中添加免疫活性物质增强鱼的体质[2]、改善养殖环境[3]、制备相关疫苗[5-6]并通过浸泡免疫方式提高异育银鲫的免疫力,以及药物防控[7]等措施降低发病率。但是,这些措施防治效果并不理想。
卵黄抗体(IgY)被认为是IgG、IgE的祖先,由禽类、两栖类动物通过血液循环将血清中抗体转移到卵黄中产生[8]。IgY能够抵抗-30 ℃低温,65 ℃巴氏消毒30 min也不会失活,4 ℃下可以保存5年,室温下可以存放6个月[9]。其耐受pH为4~10,当pH为4时,经过胃蛋白酶酶解后的Fab’片段能够保持70% IgY活性[10]。IgY经过海藻糖[11]、壳聚糖、婴儿奶粉[12,13]等物质微包膜处理后能够提高其在胃液、肠道中作用时间。与其他抗体相比,它具有成本低、制备过程简单且无创伤、特异性高、产量大、活性稳定等优点。近年来,IgY在水生动物疾病防治中的应用日益增多。雷勇等[14]发现,饲喂0.2%白斑综合症病毒(WSSV) IgY的鳌虾免疫保护率达到70%,而对照组死亡率达到100%。Lee等[15]的研究显示,IgY显著降低虹鳟(Oncorhynchusmykiss)肠道和肾脏中鲁氏耶尔森菌(Yersiniaruckeri)载量。另外,IgY还对遭受嗜水气单胞菌(Aeromonashydrophila)[16]、鲁氏耶尔森菌[15]、鳗弧菌(Vibrioanguillarum)[17]、鮰爱德华氏菌(Edwardsiellaictaluri)[18]、WSSV[19]、锦鲤疱疹病毒3(CyHV-3)[20]危害的水生动物有保护作用,一定程度上可抑制病原生物的生长[21]。因此,开发安全有效的IgY以预防异育银鲫感染CyHV-2具有重要的实践意义。本研究通过原核表达CyHV-2的ORF72衣壳蛋白免疫蛋鸡制备卵黄抗体,并初步加工为微包膜的全蛋粉及蛋黄粉;进行体外中和实验及生产性治疗试验,验证了其抗病毒特性,为下一步对鲫攻毒保护研究及用作养殖鱼饲料添加剂奠定了基础。
1 材料与方法
1.1 材料
原核表达载体pET-28a(+)、大肠杆菌BL21(DE3)表达菌株由本实验室保存;鲤鱼上皮瘤(epithelioma papulosum cyprinid,EPC)细胞由苏州大学生化与分子生物实验室馈赠;异育银鲫鳃出血病的典型病料由江苏省盐城市大丰区水产技术推广站提供;110~120日龄的健康蛋鸡购自苏州市吴江区汾湖镇红兴禽蛋经营部。HRP-His Tag、HRP标记羊抗鸡IgY抗体购自Sigma公司,鼠抗ORF72血清由实验室前期制备保存[6]。
1.2 方法
1.2.1 CyHV-2-ORF72重组蛋白的制备与验证
根据NCBI中CyHV-2基因组(GenBank登录号KT387800)中ORF72的序列设计1对引物,即5′-GCTGGATCCATGTACGGCTTAAACAACGCG-3′和5′-ACAGAATTCAGAGGCCGTTGAATGAATGTATG-3′,以CyHV-2基因组为模板进行PCR扩增。回收PCR产物,对其进行BamH I/EcoR I双酶切,插入pET-28a(+)载体构建重组质粒。测序验证后转入大肠杆菌BL21(DE3),在含氨苄青霉素的LB液体培养基中振荡过夜培养,取培养物按1 ∶100的比例接种到2YT诱导培养基中,优化IPTG诱导条件后,确定在37 ℃条件下,220 r/min振荡培养2 h后,在培养物中加入IPTG至终浓度1 mmol/L,27 ℃,100 r/min培养8 h诱导表达。12 000 r/min离心10 min收集菌体,超声破碎后用SDS-PAGE电泳鉴定诱导表达效果。将IPTG诱导后的总细菌裂解物进行SDS-PAGE电泳,然后转移至NC膜上,37 ℃孵育Anti-His Tag (HRP)抗体(1 ∶5 000稀释)2 h;PBST洗涤后加入ECL显色液用多功能荧光化学发光成像系统(ChemScope6300型,上海寰熙医疗器械有限公司)进行显影检测,验证原核表达的CyHV-2-ORF72重组蛋白(rCyHV-2-ORF72)的正确性。
1.2.2 免疫与免疫蛋的收集
将验证后的表达菌进行大量诱导,收集诱导出的含rCyHV-2-ORF72的菌体沉淀物,加入超声破碎液(50 mmol/L NaH2PO4,300 mmol/L NaCl,pH 8.0)于高压细胞破碎仪中破碎处理20 min,4 ℃条件下18 000 r/min离心20 min。将沉淀重悬于变性裂解液中(10 mmol/L NaH2PO4、10 mmol/L Tris-HCl和8 mol/L尿素),室温放置60 min后,于4 ℃条件下18 000 r/min离心15 min,收集上清。利用Ni-NTA镍柱纯化重组蛋白,收集蛋白于透析袋中,再先后放置预冷的含6、4、2、1、0.5、0 mol/L尿素复性液中分别透析5 h,再用PBS溶液(0.01 mol/L,pH 7.4)透析12 h,取出透析袋中蛋白,利用BCA蛋白浓度试剂盒测定蛋白浓度,并用SDS-PAGE电泳检测纯化效果。
将镍柱纯化后的rCyHV-2-ORF72加上其体积4%的吐温-80,并与含92%白油、6%司班-80、2%硬脂酸铝的复合佐剂以1 ∶2的比例,7 000 r/min剪切乳化10 min以上,充分乳化后的重组蛋白终浓度为0.5 mg/mL。免疫剂量为每羽蛋鸡0.5 mg,初次免疫10 d后同样剂量进行二次免疫,二免7 d后收集鸡蛋,用间接ELISA进行抽样检测,当效价达到1 ∶8 000时正式收集鸡蛋。5 d抽检一次,当效价低于1 ∶8 000时,以相同剂量加强免疫。
1.2.3 卵黄抗体的冷冻干燥加工
收集效价1 ∶8 000以上的免疫蛋,清洗消毒后分成全蛋组、蛋黄组,称重待用。蛋液中加入质量百分比分别为1%、2%、3%的海藻酸钠、β-环状糊精、羧丙基甲基纤维素,均质乳化10 min。乳化后转移至巴氏消毒罐,60 ℃消毒30 min。将消毒后的物料倒入冷冻托盘中,厚度0.5 cm,于-80 ℃冻成固体;再放入真空冷冻干燥机(Lyoquest-55plus,CHRIST公司)中,在真空度20 Pa条件下干燥48 h,获含特异抗体的冻干全蛋粉及蛋黄粉。
1.2.4 抗体特异性及效价检测
采用Western blot方法检测抗体的特异性。将诱导后的BL21总细菌裂解物、pET-28a(+)细菌总裂解物、pET-28a(+)-CyHV-2-ORF72细菌总裂解物进行SDS-PAGE电泳分离。经去离子水冲洗后,置于转膜装置,冰水浴110 mA湿转3 h,转移至NC膜,于Marker条带处剪开分为A、B二张膜,用PBST缓冲液洗涤3次,每次5 min;接着用2% Casein封闭过夜,弃去封闭液,PBS洗涤3次,每次5 min。洗涤后,分别将A膜、B膜转移至蛋黄粉溶解液(1 ∶500)和预结合过量rCyHV-2-ORF72的蛋黄粉溶解液(1 ∶500)中,37 ℃孵育1.5 h,PBST洗涤后于HRP-His羊抗鸡IgY(1∶500)中37 ℃孵育1 h,经3次PBST溶液洗涤后,加入ECL显色液,用核酸/蛋白自动曝光仪进行显影检测。
采用ELISA法检测抗体的效价。将镍柱纯化的rCyHV-2-ORF72溶于PBS溶液至终浓度为10 μg/mL,包被96孔板,以PBS液包被作为空白对照。包被后37 ℃温育1.5 h,4 ℃过夜;PBS洗涤3次,每次5 min;10%脱脂牛奶37 ℃封闭2 h;如上洗涤3次。将卵黄抗体冻干粉溶于TBS中,12 000 r/min离心10 min。取上清作为一抗,按照1∶2 000、1∶4 000、1∶8 000、1∶16 000、1∶32 000、1∶64 000的比例梯度稀释,每孔加入100 μL,37 ℃湿孵2 h,PBST洗涤3次,每次5 min,以免疫前蛋粉作为抗体的阴性对照,洗涤后每孔加入100 μL HRP标记羊抗鸡IgY抗体(1 ∶2 000),37 ℃湿孵1.5 h。洗涤后每孔加入100 μL ELISA显色剂(OPD显色底物),避光反应3~5 min后加入50 μL 2 mol/L H2SO4终止反应,用酶标仪测定样品OD490值,以P/N>2 ∶1的最高稀释度作为抗体效价的评判依据,P/N=(待测抗体的OD值-空白对照OD值)/(阴性对照OD值-空白对照OD值)。
1.2.5 抗体中和实验
将0.2 g感染CyHV-2发病的异育银鲫体肾组织加入5 mL PBS缓冲液(0.01 mol/L,pH 7.4),冰浴研磨,4 ℃下8 000 r/min离心15 min,收集上清后经0.22 μm滤膜过滤除菌获病毒粗提液,接种于单层EPC细胞上,待细胞出现病变现象后收集上清,再次感染单层EPC细胞获实验用病毒液。
用M199培养基将鼠正常血清、鼠抗ORF72血清稀释100倍,普通蛋粉、含ORF72卵黄抗体的蛋粉稀释10倍涡旋混匀后用0.22 μm滤膜过滤除菌,获抗体稀释液。
各取上述抗体稀释液、CyHV-2病毒滤液200 μL于1.5 mL EP管中,M199培养基为空白对照,26 ℃孵育1 h,期间颠倒混匀4~6次。将混合液加入到六孔板内约80%汇合度的单层EPC细胞中,每组设置3个重复孔,26 ℃孵育4 h后吸出混合液,经M199培养基洗涤3次后,加入含2%~3% FBS的M199维持培养基中继续培养。4 d后收集细胞,提取细胞基因组DNA,以EPC细胞的β-actin作为内参基因(引物序列为5′-GGAGCCGTTATCAACGACCA-3′和5′-GCTTGGTTTCATCCCTGTGC-3′),进行实时定量PCR分析,以检测CyHV-2 Hel基因(引物序列为5′-GGGTGAGGACTTGCGAAGAG-3′和5′-CGCTCGTCCGGGTTCTGCACG-3′)的相对表达水平,以此指示病毒量。PCR反应体系扩增条件为:95 ℃预变性3 min;随后39个循环:95 ℃ 10 s,60 ℃ 1 min。基因表达水平以2-ΔΔCT法[22]计算。
1.2.6 卵黄抗体生产性应用试验
盐城市大丰区某养殖户的4个养殖池,1、2、3、4号池的面积分别为30 015、40 020、23 345、53 360 m2。3月下旬放养异育银鲫3尾/m2,平均规格150 g/尾。5月8日发现各池均出现鲫鱼死亡,病鱼体重约250 g/尾,具明显的鳃出血症状,从5月8-15日,各池鱼的死亡量从数尾快速上升为数百尾。发病池的水质良好,溶氧在6.2~8.5 mg/L,pH值7.5~8.0,透明度达到35 cm左右。当地水产技术推广部门进行PCR检测分析证实鲫鱼罹患鳃出血病。选择1、2号池为对照池,采用口服维生素C,晚上9点开始增氧至第2天清晨;定期泼洒微生态制剂调节水质等常规处理方式提高病鱼的抗应激能力,减少死亡率。3、4号池作为试验池,在常规处理方式的基础上全池泼洒和口服效价为1 ∶16 000的抗CyHV-2病毒的卵黄抗体。其中3号池在第1、2天每天全池泼洒,剂量为1 g/m3;第3天开始投喂卵黄抗体,按饲料重量0.15%的剂量拌入鲫鱼商品饲料中制成功能性饲料,按1%鱼体重的投喂量投喂。4号池将饲料中卵黄抗体添加量增加到0.30%,其他治疗方法与3号池相同。在病情好转后,饲料投喂量增加到2%,早晚各投喂一次。试验从5月13日开始,7月中旬鲫吃食、生长完全正常,水温稳定在28 ℃后改投普通商品饲料。试验过程中每天记录死鱼情况、鱼的摄食状态及用药情况,9月底捕捞上市,统计成活率、规格与产量。
1.3 统计方法
使用SPSS统计软件19.0进行数据统计分析,所有定量数据均表示为平均值±SEM;差异显著为P<0.05,差异极显著为P<0.01。以GraphPad Prism 5.0绘图。
2 结果
2.1 CyHV-2-ORF72重组蛋白原核表达及纯化
CyHV-2-ORF72重组蛋白原核表达产物大小为43 kDa左右,与预测大小吻合,推测诱导出了rCyHV-2-ORF72蛋白。表达产物经破碎、离心、镍柱纯化后获较纯的rCyHV-2-ORF72(图1)。图1显示,rCyHV-2-ORF72主要表达在包涵体中,少量为融合蛋白;超声与高压破碎、镍柱纯化后获较纯的重组蛋白。经镍柱纯化后的重组蛋白纯度在80%以上。镍柱纯化的蛋白经复性、透析后测得蛋白浓度为1.2 mg/mL。用Anti-His Tag (HRP)抗体进行Western blot鉴定结果显示,原核表达产物在43 kDa附近出现褐色条带,空载体pET-28a(+)总细菌裂解物未出现条带,进一步证明诱导出的重组蛋白特异性强(如图2)。
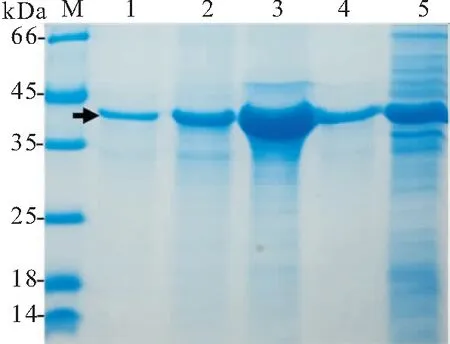
图1 CyHV-2-ORF72重组蛋白诱导表达的SDS-PAGE鉴定Fig.1 SDS-PAGE analysis of recombinant CyHV-2-ORF72 capsid protein
M,蛋白质分子量标准;泳道1,镍柱纯化后的CyHV-2-ORF72(剪头所指);泳道2,高压破碎后的离心沉淀;泳道3,诱导细菌超声破碎后离心沉淀;泳道4,诱导细菌破碎后离心上清;泳道5,诱导后pET-28a(+)-CyHV-2-ORF72/BL21总细菌裂解物M,蛋白质分子量标准;泳道1,pET-28a(+)-CyHV-2-ORF72/BL21总细菌裂解物,剪头所指为CyHV-2-ORF72;泳道2,pET-28a(+)/BL21空载总细菌裂解物。

图2 CyHV-2-ORF72重组蛋白的Western blot验证Fig.2 Western blot detection of recombinant CyHV-2-ORF72 capsid protein
2.2 冻干全蛋粉、蛋黄粉中ORF72抗体特异性及效价检测
全蛋粉、蛋黄粉中ORF72抗体特异性的结果如图3。图3A显示,孵育CyHV-2-ORF72抗体后,在43 kDa大小附近出现一条褐色目标条带,而阴性对照组未出现条带,表明卵黄抗体中产生了CyHV-2-ORF72重组蛋白的特异性抗体;进一步用过量的rCyHV-2-ORF72预结合卵黄抗体(即封闭抗体后再进行Western blot检测),在43 kDa处的褐色条带消失(图3B),证明了卵黄抗体的特异性。

图3 蛋粉中抗ORF72衣壳蛋白抗体的特异性检测Fig.3 The specificity of anti-ORF72 capsid protein antibody in egg yolk and whole egg powder
M,蛋白质分子量标准;泳道1,BL21空菌总细菌裂解物;泳道2,pET-28a(+)/BL21空载总细菌裂解物;泳道3,pET-28a(+)-CyHV-2-ORF72/BL21总细菌裂解物。剪头所指为CyHV-2-ORF72。A,蛋粉稀释液;B,以过量的CyHV-2-ORF72蛋白预结合后的蛋粉稀释液利用间接ELISA检测冻干全蛋粉与蛋黄粉中抗体的效价,根据P/N>2.1计算,结果显示,全蛋粉中的CyHV-2-ORF72抗体效价为1 ∶8 000;蛋黄中的效价达到1 ∶16 000。全蛋粉中的抗体效价略低于蛋黄。
2.3 卵黄抗体与CyHV-2的亲和力
对EPC细胞中CyHV-2病毒解螺旋酶(Hel)基因进行实时定量PCR分析,并以空白对照中Hel基因的Ct值为基线1,计算其它各组Hel基因的表达量,结果如图4所示。rCyHV-2-ORF72免疫小鼠的抗血清、抗CyHV-2-ORF72卵黄抗体孵育CyHV-2病毒后,Hel基因表达水平分别下调28%倍和60%,与空白对照组和阴性对照组差异极显著,说明鼠IgG、鸡蛋IgY均降低了病毒量,且鸡蛋IgY比鼠IgG对CyHV-2的亲和力更强。而孵育普通卵黄抗体后,反而促进病毒的繁殖,病毒的相对表达量上升了0.25倍,与空白对照组差异显著。结果表明制备的CyHV-2-ORF72卵黄抗体具有较好的病毒中和作用。
2.4 抗CyHV-2卵黄抗体对患鳃出血病异育银鲫的免疫保护作用
于5月16日、17日对23 345 m2与53 360 m2试验池连续两次泼洒CyHV-2-ORF72卵黄抗体后,病鱼死亡数量不再上升。两试验池分别连续投喂0.15%和0.30%的抗病毒功能性饲料7 d后,鱼的死亡量下降至100尾以下。到7月中旬,水温稳定在28 ℃以上时,鱼摄食、生长恢复正常。至9月底10月初,两个试验池鱼的平均规格分别达420、470 g/尾。而两个对照池鱼死亡率仍快速上升,5月26日达到1.5尾/m2,成活率仅有31.5%和28.3%。试验表明,卵黄抗体对患病异育银鲫具有较好的免疫保护作用,投喂0.30%卵黄抗体的池塘中,鱼的成活率高于投喂0.15%的池塘,具体见表1。
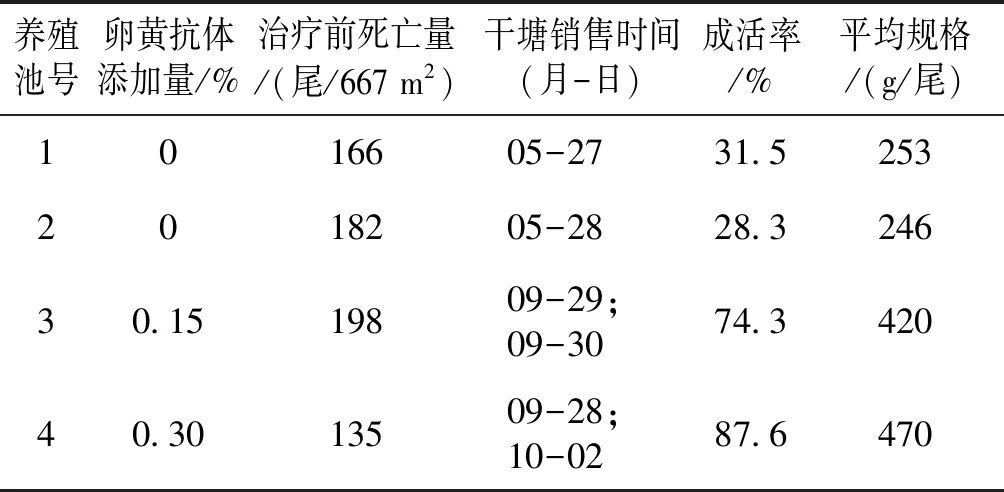
表1 抗CyHV-2卵黄抗体对异育银鲫鳃出血病的治疗效果Tab.1 Therapeutic effect of anti-CyHV-2 yolk antibody in allogynogenetic crucian carp with haemorrhagic gill disease
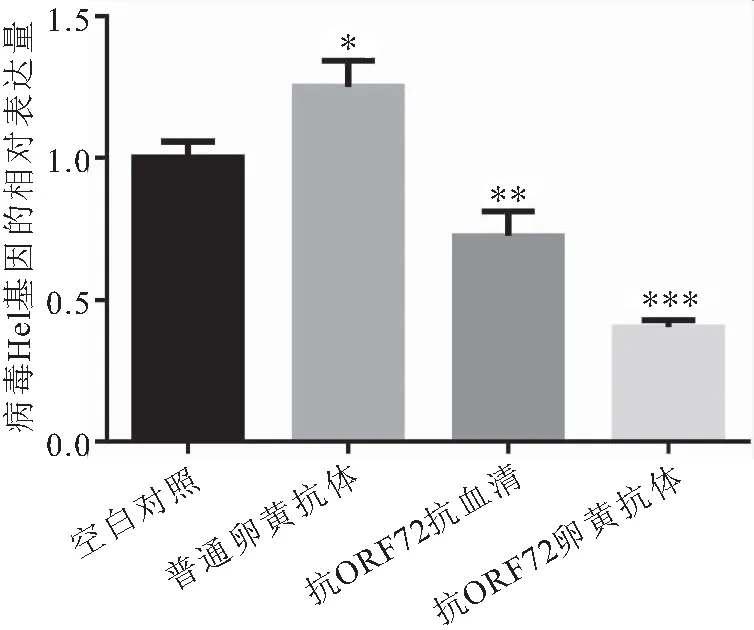
图4 CyHV-2-ORF72抗体的病毒中和活性Fig.4 The virus-neutralizing activity of anti-CyHV-2-ORF72 antibody *、**和***分别表示与对照组差异显著水平为 P<0.05、P<0.01和P<0.001。
3 讨论
研究显示,CyHV-2基因组DNA上存在150个保守的开放阅读框,已证实可编码74种蛋白,包括3种衣壳蛋白、18种膜蛋白和53种其它蛋白,其中包括衣壳蛋白ORF72在内的8种蛋白被发现存在免疫原性[23]。孔善云等[24]证实,CyHV-2的衣壳蛋白72具有较好的免疫原性,鼠抗重组ORF72抗体可以识别CyHV-2病毒粒子的衣壳蛋白72。此外,有研究也显示,单克隆抗体可特异性识别ORF72蛋白[25]。因此,本研究选取编码蛋白衣壳的基因ORF72,进行原核表达制备卵黄抗体,Western blot与ELISA检测分析显示,该抗体具有特异性强、效价高(1 ∶16 000)等特点。体外病毒中和实验则表明,抗CyHV-2-ORF72的鸡蛋IgY与CyHV-2病毒的亲和力显著强于鼠IgG。生产性临床试验结果表明,投喂0.15%至0.30%抗CyHV-2卵黄抗体的全蛋粉,对异育银鲫具有显著的免疫保护作用。充分显示了抗CyHV-2-ORF72的IgY作为生物渔药防控异育银鲫鳃出血病的潜力。
卵黄抗体与制备其他动物的血清抗体相比,具有成本低、无创伤、产量大等优点,一只鸡单月可产生相当于500 mL血清抗体的抗体,含有2%~10%特异性IgY[26],一年约产生40 g IgY抗体,而一只兔2~3周采一次血,一年约产生1.3 g抗体,使得大规模制备卵黄抗体成为可能。利用Western blot进行检测发现,我们制备的卵黄抗体,稀释500倍后能够结合ORF72重组蛋白,而且将ORF72蛋白与卵黄抗体预先结合(封闭抗原表位)后作为一抗孵育,未能够检测到目的蛋白条带,证明了卵黄抗体的特异性。这一结果也为生产应用抗CyHV-2预防和治疗异育银鲫鳃出血病提供了理论依据。
IgY抗体通常对消化酶胰蛋白酶和胰凝乳蛋白酶非常稳定,尽管在低pH值下胃蛋白酶会造成很高的活性损失[27]。研究表明,在pH值大于3时,IgY对胃蛋白酶具有相对抗性,并保持一定的抗原亲和活性。然而,在pH 2下,由于消化,IgY的活性降低[28]。利用β-环糊精等包膜剂包膜IgY,已被证实在南美白对虾的肠道中保持了IgY的活性[29]。因此,在制备抗CyHV-2的IgY时,我们采用海藻酸钠、β-环状糊精、羧丙基甲基纤维素等优质包膜剂包膜,以保护IgY活性;同时,我们发现全蛋粉与蛋黄粉中卵黄抗体的效价差异不大,如作为饲料添加剂,蛋清还可补充营养元素。因此,采用简单的加工方式生产全蛋粉,可实现卵黄抗体的规模化生产。所制备的IgY可作为饲料添加剂口服、或采用浸泡等方式群体处理养殖动物,实现生产性规模化防病抗病目标。
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
健康体检与管理(2022年4期)2022-05-13
心理学报(2022年2期)2022-02-17
现代临床医学(2021年5期)2021-11-02
延安大学学报(医学科学版)(2021年3期)2021-10-25
现代检验医学杂志(2021年4期)2021-08-10
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年2期)2021-03-29
天津医科大学学报(2021年2期)2021-03-29
分析化学(2018年12期)2018-01-22
