
降水变化和氮沉降影响森林叶根凋落物分解研究进展
2021-02-06 05:55谭向平申卫军
生态学报 2021年2期
谭向平,申卫军
中国科学院华南植物园退化生态系统恢复与管理重点实验室,广州 510650
自工业革命以来,化石燃料的大量燃烧和化学肥料的过度使用等人类活动导致了明显的全球环境变化。IPCC的报告指出全球降水格局已经发生明显变化,未来极端降水事件(如干旱和强降水)趋于频繁,将会加剧部分地区的干旱程度或引发洪涝灾害[1]。同时,大气氮(N)沉降量的急剧增加也已导致了一系列的环境问题,如加速土壤酸化和森林衰退、减少生物多样性等[2]。预计到2050年,全球氮沉降量将增加至200 Tg N/a,是20世纪90年代的两倍[2]。诸多研究表明全球环境变化会直接和间接地影响陆地生态系统的结构和功能,从而对水资源、生态系统服务功能和社会经济发展等产生深刻的影响[3]。因此,深入理解森林生态系统的关键生态过程对全球环境变化的响应和适应机制,成为当前最为关注的前沿性重大科学问题之一[4]。
在森林生态系统中,90%以上的植物净初级生产量通过凋落物(枯枝落叶、繁殖器官、枯死的树根等)的形式返回地表,其在维持森林生态系统初级生产力、净碳储量、物质循环等方面起着十分重要的作用[5]。凋落物分解是森林生态系统中极为关键的生态过程之一,一方面凋落物分解释放的CO2是全球碳循环的重要组成部分[6];另一方面凋落物分解产物也是土壤有机质的主要来源[7]。因此,研究凋落物分解及其对环境变化的响应对了解森林生态系统碳循环具有重要的现实意义。在不同类型森林生态系统中,针对控制凋落物分解的影响因子以及凋落物分解对环境变化的响应已开展了大量的研究,发现并获得了一些重要的科学现象和规律[8-9],特别是在持续高氮沉降对凋落物分解的作用机理方面有了比较清楚的认识[10-11]。然而,对环境变化调控森林凋落物分解的了解仍有很大的不确定性,究其原因主要是:(1)不同类型凋落物(例如叶和细根凋落物)的分解过程受自身化学性质的影响[12],导致同一种植物不同部位的凋落物对环境因子的变化存在不同的响应行为[13-14],但其潜在的生物学机制尚不清楚。这极大地降低了以叶凋落物分解特征为基础建立的森林物质能量循环模型的正确性和准确性。(2)环境因子之间通常存在着复杂的相互作用模式,单因素控制实验往往不能完全模拟多环境因子对陆地生态系统的共同作用[15]。尽管凋落物分解对单一气候或环境因子变化的响应已有大量的研究,发现存在促进、抑制和无影响3种方式[16-17],但对于多环境因子耦合如何影响凋落物分解(协同、拮抗、或无交互作用)还缺乏深入、系统的研究[18-19],这样就难以支撑陆地生态系统固碳速率和潜力的准确评估[20]。(3)当前微生物生态学研究的关键挑战是量化微生物类群在自然界中的功能[21-22]以及其对多环境因子的响应机制[23]。微生物群落可能会通过改变某个微生物物种或功能群的资源利用特征,导致在凋落物分解过程中每个微生物所起的具体作用难以确定[24],加之环境因子对微生物群落的复杂影响[23],因此阐明微生物群落结构和功能的变化仍然是解释森林凋落物分解如何响应环境变化的关键途径。
本文在系统梳理国内外叶根凋落物分解研究的基础上,聚焦于微生物群落功能研究的瓶颈,结合全球变化的大背景,尝试从凋落物质量、微生物群落结构和功能的角度,深入探讨森林地上和地下凋落物(叶和细根)分解对降水季节分配变化和高氮沉降响应的机理;侧重总结降水季节分配变化和高氮沉降如何通过影响微生物群落来调控森林凋落物的分解过程。最终,构建一个环境变化影响森林凋落物分解的概念图,并为以后凋落物分解的研究提供新的视角。
1 概述森林凋落物分解及其环境响应
所有陆地生态系统都是由地上和地下两部分组成,这些组成部分的相互作用直接影响群落和生态系统层面的生态过程和属性,因此整合地上-地下生态的研究可以加强我们对生物多样性的功能及其对全球变化反馈调节机制的理解[25]。然而,目前的研究通常基于叶凋落物的特征来预测陆地生态系统有机质的循环,这对植物性状与碳、养分循环之间反馈的理解容易产生偏差[26],因为植物叶和根系凋落物之间的可分解性可能存在显著差异[27]。在全球或区域尺度上,凋落物质量(N、C∶N比、木质素:N比)和气候条件(温度、降水量、蒸散量)被认为是控制凋落物分解的主要因素[28-29]。基于叶凋落物分解实验,研究者利用凋落物氮含量、C∶N比、木质素:N比和年平均气温构建了分解模型去预测全球森林凋落物的分解速率[29]。相比地上凋落物,细根暴露于完全不同的分解环境,因此控制细根分解速率的因素随外界环境和植物类型的不同而发生变化[30]。整合分析(Meta-analysis)表明细根C∶N比和钙(Ca)含量是影响全球森林生态系统根系分解速率的主控因子[30],但最新的整合分析发现N、磷(P)和木质素含量才是控制细根分解的主要因素[31]。也有研究观察到细根分解速率与初始C∶N比和Ca含量均无关[32-33],而取决于非结构性碳水化合物、总酚和单宁的含量[12]。由此可见,控制植物地上和地下凋落物分解的初始理化性质可能存在差异,其潜在导致同一物种叶和根系凋落物分解对环境因子变化的响应不一致。
近年来,全球环境变化影响相同植株地上和地下凋落物分解的研究仅有少量报道。García-Palacios等[34]的研究表明降雨量减少通过降低土壤微生物生物量以及改变其生理特征抑制了地中海麻栎(QuercusilexL.)细根的分解,而对其叶凋落物分解没有影响。氮添加会对马尾松(Pinusmassoniana)、荷木(Schimasuperba)、糖枫(Acersaccharum)、栎树(Quercusellipsoidalis)吸收根(1—2级细根)的分解产生负面影响,而对叶凋落物分解没有影响或促进其分解[13-14,35]。另有研究发现氮添加均会抑制湿地松(Pinuselliottii)叶和吸收根的分解,但具有不同的抑制机理[36]。外源氮通过与酸不溶组分形成难分解物质来阻碍吸收根的分解,而叶凋落物分解被抑制是因为胞外酶活性降低的缘故[36]。也有学者提出了不同的作用机理,认为氮添加对木质素、单宁和蛋白质分解酶活性的抑制作用可能是影响细根分解的重要机制[14,37- 39]。外源磷添加可增加叶凋落物分解而不影响根凋落物分解,但同时添加氮磷对叶根凋落物分解没有影响[13]。综上表明,森林地上和地下凋落物分解对环境条件变化的响应机理并不一致,尤其是多环境因子对同一植株叶根凋落物分解的影响尚不清楚。
尽管比较森林叶和细根凋落物分解过程的研究逐渐增加,但这些研究的关注点大多聚焦在凋落物质量和形态特征,对参与地上和地下凋落物分解的微生物群落与凋落物化学组成之间关系的认识仍然匮乏。例如,叶和细根凋落物分解过程中养分的释放模式不尽相同[40]。主要是因为我们对凋落物分解过程中不同微生物功能群的动态及其与底物的相互作用还知之甚少[24],特别是胞外酶的生产和周转与微生物资源需求和底物质量间的关系仍然不清楚[41]。虽然有研究测量了土壤动物对森林凋落物分解速率的贡献,并发现叶和细根凋落物分解过程中土壤动物群落的演替存在明显的差别[42-43],但对参与细根分解的微生物群落结构和功能尚不清楚[44- 46]。
2 降水变化和氮沉降对叶根凋落物分解的影响
凋落物分解取决于多种生物和非生物因子在空间和时间尺度上的相互作用(图1)。环境变化通过改变分解过程中的生物化学反应速率和微生物生理直接影响凋落物分解,同时也可以诱导植物的形态和生理变化,特别是改变植物组织的化学成分,从而间接影响凋落物分解[5,47-48]。目前,对全球变化影响森林凋落物分解的研究大多集中在单一环境因子改变对叶凋落物分解的直接影响,特别是模拟氮沉降增加效应的研究。全球范围内的研究结果表明外源氮输入对森林叶和细根凋落物分解的影响有着促进、延缓、无作用3种不同的表现[12,49- 52]。由于已有大量综述和整合分析文章报道加氮处理对凋落物分解的直接影响[5,49,51,53- 56],因此本文不再赘述,而着重阐述降水变化及其与氮沉降的交互作用对凋落物分解的影响。
2.1 降水变化和氮沉降对凋落物分解的间接影响
全球变化引起土壤水分和氮素的变化会直接或间接地影响植物生长、碳吸收及光合产物的分配,从而改变叶和细根的产量以及化学组成,间接调控凋落物的分解[47,57]。一项长达13年的降水控制实验表明,年降雨量减少29%显著降低地中海常绿森林叶凋落物(C∶N≈61)总酚和缩合单宁的含量,但对细根凋落物质量(C∶N≈78)没有影响[34]。叶根响应行为的不同致使影响各自凋落物分解的因子存在明显的差别,减少降雨处理主要是通过改变凋落物质量影响叶凋落物分解,而细根凋落物分解主要受到土壤微生物群落变化的影响[34]。同样在地中海灌木林的研究发现,考虑到凋落物初始质量的改变,减少降水处理对凋落物分解的抑制率从18%增加到26%[48]。对长白山阔叶红松林的研究表明,降水减少诱导黄柏(Phellodendronamurense)和紫椴(Tiliaamurensis)的细根直径减小,但其具有更强的吸收能力;水曲柳(Fraxinusmandshurica)的细根变粗但吸收能力减弱[58]。然而,细根形态的变化是否会影响细根凋落物分解对降水变化的响应尚不清楚。
整合分析显示氮添加引起森林叶凋落物中铝、硼、钙、镁、锰、磷、锌含量降低,提高了叶凋落物的氮含量并降低C∶ N比[59-60],也有研究发现氮添加会提高叶凋落物非结构性碳水化合物含量,而增加细根凋落物木质素的含量[39,61]。初始质量的改变对凋落物分解速率影响迥异,其可能促进叶凋落物初期的分解速率[62]或抑制叶根凋落物后期的分解速率[60,63]。仅有的研究发现氮沉降增加与降水减少交互作用对细根形态的影响取决于树种,可能存在拮抗或协同作用,例如其交互作用对细根生物量的影响大于单一因素[58]。可见,降水格局变化和氮沉降增加背景下,森林凋落物初始质量和数量的改变对其分解的影响可能因植物功能器官、种类、生态系统的不同而不同。同时,也表明仅采用元素化学计量比很难预测全球变化对凋落物分解速率的影响[64],因为化学计量难以反映凋落物中杂聚物和酚类化合物含量及结构的改变[47]。例如,在季节性干旱的热带森林生态系统中,年降水量低的地区树木具有较高的角质和木栓质[65]。地中海地区富含酚类化合物的植物会通过增加木质素含量和C∶ N比来应对极端干旱[59]。因此,探讨未来环境变化对植物凋落物分解的影响应当考虑植物各器官凋落物化学组成和结构的改变。
2.2 降水变化和氮沉降对凋落物分解的直接影响
在农田和草地生态系统,降水减少显著降低了凋落物的分解速率[66]。在森林生态系统中,降雨控制实验发现降水减少对叶凋落物分解的影响存在无效应或减缓的情况[34,67]。例如,降水减少20%—50%显著降低了亚热带常绿阔叶林和地中海橡树林叶凋落物分解速率,主要是因为土壤含水量急剧降低[67- 69]和微生物及土壤动物活性的减弱[70]。但也有研究表明降雨量的减少对叶凋落物的分解没有影响,而土壤微生物生物量的减少和生理特征的改变抑制了细根的分解[34]。相对减少降雨,降水(≥10 mm)频率与森林叶凋落物分解速率呈正相关[71]。在热带森林的研究表明,模拟降水增加可能促进或抑制凋落物的分解,产生积极的影响是因为降水增加可以促进凋落物可溶组分的淋失以及微生物活性;当水分过饱和时,会导致溶解氧含量降低进而抑制微生物活性[72- 74]。这些研究表明,降水变化模式的不同(减少、增加、季节分配、降雨量大小和频次)也是导致凋落物分解响应差异的因素之一[75]。
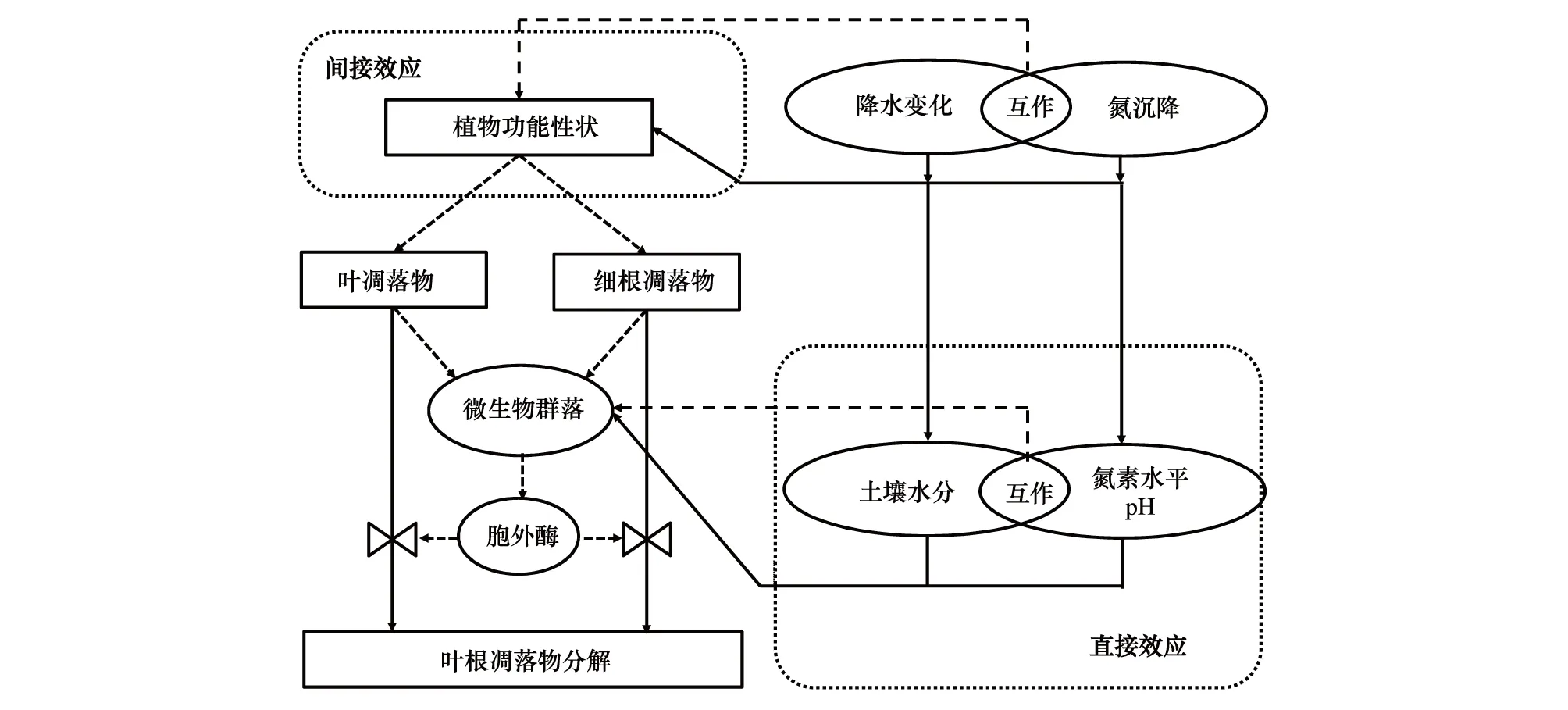
图1 降水变化和大气氮沉降影响森林凋落物分解的主要途径Fig.1 Direct and indirect effects of precipitation variability and atmospheric nitrogen deposition on the forest litter decomposition环境变化主要通过直接效应和间接效应影响凋落物的分解;实线路径表示以往研究对该过程有一定的认识、虚线路径表示对该过程的认识仍非常匮乏;本概念模型基于Suseela and Tharayil [47]提出的全球变化影响凋落物分解示意图演变而来
迄今为止,关于降水格局变化和氮添加交互影响凋落物分解的野外实验主要建立在草原[76-77]和荒漠生态系统[78]。模拟降水增加和氮沉降的双因子实验表明,尽管处理增加了半湿润半干旱草原和荒漠地表的水分和氮素,但其对叶凋落物分解的影响很小[78-79]。在半干旱草原生态系统中,单独施氮和加氮水处理显著抑制叶根凋落物的分解速率,但单独加水处理对叶根凋落物的分解几乎没有影响[76]。也有研究发现降雨量变化和增加氮素供应可以促进高草草原叶凋落物的分解,但两者的交互作用随凋落物种类而改变[77]。在森林生态系统中,增加桉树人工林灌溉水和氮肥的投入对凋落物早期分解(60 d)的影响较小,且不存在交互作用[79]。近年来我国学者相继在温带森林和热带森林建立降雨格局变化(增加或减少)和氮沉降增加双因子野外实验平台[80- 82]。但目前仅有一项研究关注凋落物分解,其结果表明减少降雨和氮添加处理抑制了阔叶红松林叶凋落物的分解速率,但两者的交互作用很弱,然而对凋落物分解过程中氮的固定具有明显的交互作用[83]。已有的研究结果表明两者对凋落物分解的作用呈现出拮抗、协同或无交互作用的特征,因此当前的研究仍需深入阐明降水格局变化和氮沉降交互作用影响森林凋落物分解的特征,特别是要加强在热带、亚热带森林生态系统的研究。
2.3 降水变化和氮沉降影响凋落物分解的微生物机理
环境因子和凋落物质量对分解过程的影响大部分是通过土壤微生物的活动来实现的(图1)。当前已有少量的研究发现,降水减少通过影响土壤微生物多样性[69],降低微生物生物量[34]和胞外酶活性[84]来抑制凋落物的分解。Allison和Goulden[85]采用DEMENT模型模拟干旱对草原凋落物分解的影响发现,尽管干旱可以增加耐旱微生物的丰富度,但微生物之间的互作减少了胞外酶的分泌以及维持碳利用效率成本的增加,导致凋落物的分解速率降低。同时,还发现降水季节变化比单纯降水量减少的处理对草原土壤微生物群落的影响更大[85]。然而,关于降水季节分配变化对森林凋落物分解的影响及其微生物学机理尚缺乏相应的实证研究[85-86]。
大量的研究发现,氮沉降持续增加背景下参与凋落物分解的微生物群落结构和功能(例如胞外酶活性)的改变是调控分解的核心机制[24],并形成了三种理论假说:(1)计量比假说:凋落物分解是通过底物的化学计量比(例如:C∶N比等)和微生物对资源的需求来控制[87]。在高C∶N比凋落物分解过程中,添加氮缓解了微生物对氮的需求,从而促进凋落物中结构性糖的分解来满足能量的需求[62]。(2)微生物氮开采假说:微生物通过分泌木质素酶(氧化酶)分解木质素以获取被保护的氮[88]。当外界条件满足微生物对氮的需求时,微生物可能会减少木质素酶的分泌,致使凋落物分解受到抑制。(3)褐变效应:外源氮能够引起多酚和氨基化合物之间的随机反应,合成难分解的芳香化合物来抑制微生物和胞外酶活性,导致凋落物的分解速率降低[89]。然而,这些假设不能完全解释外源氮添加对凋落物分解的抑制作用[90-91]。例如,有证据表明氮添加可以通过减少木质素分解酶的活性来抑制分解[92],可能是因为参与木质素分解真菌相对丰度的减少[11],微生物群落组成的改变[93]或功能基因表达的下调[94]。但也有研究发现氮添加降低凋落物的分解速率与木质素降解酶活性和微生物群落的变化无关[95-96]。我们前期的研究发现外源氮添加只有同时降低氧化酶和水解酶的活性才能抑制凋落物的分解[41]。
综上所述,环境变化影响凋落物分解的微生物学机理已有初步的了解,然而微生物群落与凋落物分解速率之间的关系及其对环境变化的响应规律仍缺乏统一的认识。在全球范围内分解者功能多样性的丧失会减少凋落物C、N的损失[97];但有研究表明细菌群落结构决定凋落物分解速率,而真菌群落结构的变化对凋落物分解速率没有影响[18,98];也有研究发现在短时间内(1年)微生物群落结构和功能(以凋落物分解率来衡量)对全球变化的响应并不同步[99]。出现对立的观点和结论主要是因为缺乏对不同微生物种类或群体如何控制凋落物分解以及不同类群间互作的了解[24]。因此,阐明参与凋落物分解的微生物群落结构和功能之间的关系,特别是胞外酶的生产和周转与微生物资源需求和底物质量间的关系仍是当前研究的重点和发展方向,同时也可以加深对全球变化引发的效应机制和长期作用后果的认识[18]。例如,通过控制初始微生物群落结构的实验表明,初始微生物群落结构的差异以及不同类群对环境因子的抵抗力和恢复力,决定着凋落物分解对温度和降水变化的响应程度[18,99]。另外,单一环境因子的研究还不能完全说明在其他环境变化条件下,微生物群落变化影响凋落物分解的普遍规律,因为土壤中存在着多种重要的微生物并且它们自身及与土壤性质之间存在错综复杂的关系[18]。例如,增雨和加氮对草原土壤微生物群落和功能的影响存在极为复杂的交互作用[100-101],而在荒漠生态系统的研究发现两者不存在任何的交互作用[102]。
3 新技术方法在凋落物分解机制研究的应用
森林凋落物分解的研究主要采用野外分解袋法、实验室模拟分解法(例如测定分解产生的CO2)、同位素示踪法(例如14C)、现存量估算法(例如积累平衡模型)、生态模型(例如DayCent)模拟[103- 105]。想要准确量化凋落物分解速率,所采用的方法不能排除凋落物分解过程中的任一要素即浸出、物理破碎和生物分解,然而现有方法均具有不同的优缺点[55,103-105]。尽管野外分解袋法会排除物理破碎作用,但其仍然是应用最多、且被推荐用来研究凋落物分解速率对环境变化响应的重要方法[103]。凋落物分解的研究不仅是要精确测定凋落物的分解速率及其影响因素,而且还要解析凋落物分解过程中元素的去向,比如C的分配[7,106]。Cotrufo等[7]研究发现在分解早期,凋落物中大多数非结构性化合物经过微生物的新陈代谢作用,以微生物残体等形式进入土壤形成矿物复合态有机质(被称之为有机质-微生物途径);然而在分解后期凋落物碎片通过物理迁移路径进入土壤形成颗粒态有机质。上述研究表明微生物群落既控制凋落物的分解进程,又是土壤有机质形成的关键环节。随着分子生物学和同位素示踪技术的发展,稳定同位素探针(SIP)等方法为追踪凋落物分解过程中元素的分配提供了新手段(例如SIP-PLFA、SIP-DNA),而且还有利于解析凋落物分解的微生物学和酶学机制[103,107]。
从生态学的角度来说,对微生物群落结构认识的最终目的是为了理解其功能。但由于缺乏合适的方法,对凋落物分解过程中不同微生物的种群和功能的动态以及与底物质量相互间的作用还知之甚少[98],特别是对全球变化的响应仍缺乏相应的研究[62]。目前我们对微生物功能群的认识仍然局限于磷脂脂肪酸(PLFA)以及分子生物学技术,虽然这些技术能够鉴定出参与特定代谢途径的微生物功能基因数量和种类,但是不能真实地反映微生物的代谢途径[108]。这极大的阻碍了我们对微生物群落结构和功能的理解[109]。相比DNA和RNA,大多数蛋白质都具有固定的代谢功能,可用于了解生物群落中特定微生物的活性。因此,对给定生境的微生物蛋白质的识别可以分析其系统起源及时间分布,为解析生物地球化学过程中微生物多样性提供新的手段。
宏蛋白质组学用于描述土壤[110]、水下沉积物[111]、叶际[112]、人体肠道[109]中微生物的功能取得良好的成果。Schneider等[108]利用宏蛋白组学对森林凋落物层研究表明微生物丰富度和胞外酶活性直接决定微生物群落的分解能力。胞外酶被认为是外界营养条件调控下细胞代谢的产物,是微生物功能多样性的体现[113]。通过宏蛋白组对它们的鉴定有助于我们建立特定物种或微生物的出现与土壤的生物地球化学关系,进而帮助我们理解森林凋落物分解的动态变化以及环境变化影响其分解速率的机理[110]。因此,未来的研究工作需要通过多组学的研究将胞外酶与微生物功能群整合起来,加深我们对环境改变影响微生物群落结构和功能的了解[114],为进一步解释全球变化如何影响森林凋落物分解提供新的视角[110]。
另一方面,元素和化学组分的含量及比值是衡量凋落物质量最常用的化学指标,但这些指标并不能完全揭示凋落物各化学组分的分解机制。有研究者采用更精细的化学分析手段(例如,核磁共振光谱和傅里叶变换红外光谱)来研究凋落物分解过程中主要化学组分的变化,发现叶凋落物分解过程中分子结构的变化与微生物群落的演替密切相关[39,115]。因此,结合有机化学和分子生物学的手段可以进一步探明凋落物化学组分变化与微生物群落动态之间的联系,并阐明地上和地下凋落物分解的关系,从而对森林地上和地下凋落物分解对环境变化的响应及潜在的生物化学机理形成全面的认识,进而在物种水平上理解森林生态系统凋落物分解规律及其对环境变化的响应方式。
4 总结与展望
目前,在全球范围内降水控制实验主要集中在中纬度地区的草原、荒漠以及森林生态系统,针对低纬度地区森林生态系统的研究相对较少,并且大多降水控制实验主要是探索增加或者减少降雨对生态系统的影响。但关于降水季节分配变化的研究还十分有限,这极大地影响了预测凋落物分解对全球变化的响应[116-117]。例如,近年来我国东南部地区降水格局发生明显的变化:总降水量不变,但干季降水次数减少、雨季强降水事件增加,预测未来该地区的气候可能呈现出明显的干季更干、湿季更湿的特征[118]。特别是该区域还面临着严重的大气氮沉降(35 kg N hm-2a-1),且呈继续增加的态势[119]。相比温带和寒带地区,热带地区的植物通常利用增加杂聚物和酚类化合物来抵御高温和高紫外线辐射的胁迫,使得它们面临极端干旱或降水格局变化时更容易改变组织的化学组成[47]。然而,在凋落物化学组成如何响应未来环境变化以及其对分解的影响尚缺乏清晰的认识。
氮沉降增加对生态系统的影响在很大程度上依赖于自然降水,例如降水季节分配变化会导致南亚热带森林雨季和旱季期间水分和氮素供应状况的改变,进而影响氮素生态效应的发挥,可见两者的交互作用能够对森林生态系统的各关键生态过程产生更为复杂的影响[20]。然而,当前的野外控制实验多局限于模拟单一环境因子的作用,其结果并不能解析多因素对森林生态系统的影响。因此,亟需探明降水季节分配变化和氮沉降增加之间的交互作用对森林生态系统物质和能量循环的影响,为模拟和预测陆地生态系统对未来全球变化的响应提供理论基础和数据支持。考虑到控制地上和地下凋落物分解因素的差异[12],以及微生物群落对土壤水分变化、外源氮添加的响应方式和机理存在差异[86,120-122],如何准确回答全球变化对森林凋落物分解的影响及其微生物学机理显得尤为迫切。综上分析,目前我们认为在以下三个方面的研究亟待加强:(1)凋落物质量是如何通过微生物群落调控地上和地下凋落物的分解?过去研究凋落物质量对分解的影响仅关注氮和大分子物质(半纤维素、纤维素、木质素等)的含量,以后的研究应注重通过测定各化学组分的基本结构来探讨凋落物中大分子物质的分解动态,并结合微生物群落结构和功能特征,从而更为准确地解析分解过程中凋落物质量变化与微生物群落之间的关系;(2)降水格局变化和氮沉降增加对森林凋落物质量的影响以及质量改变对凋落物分解的影响程度,尤其是加强对细根凋落物质量和分解的研究;(3)降水格局变化和氮沉降增加对参与凋落物分解的微生物功能群的影响及其对凋落物分解的贡献。
致谢:华南师范大学胡中民教授、中国科学院西双版纳热带植物园刘胜杰副研究员帮助写作,特此致谢。
猜你喜欢
当代水产(2022年8期)2022-09-20
草业学报(2022年9期)2022-09-16
东北林业大学学报(2022年6期)2022-07-25
西北农业学报(2022年4期)2022-05-19
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
绿色科技(2021年5期)2021-04-09
人民黄河(2020年12期)2020-12-30
山西农业科学(2020年9期)2020-09-14
河南科学(2020年3期)2020-06-02
