
起测径对南盘江流域次生林中枯立木空间结构的影响
2021-05-25 02:01何吉安李惺颖喻素芳李远干李远发
西北植物学报 2021年4期
何吉安,刘 通,李惺颖,喻素芳,李远干,李远发*
(1 广西大学 林学院,广西森林生态与保育重点实验室,南宁 530004;2 北京林业大学 园林学院,北京 100083;3 广西壮族自治区森林资源与生态环境监测中心,南宁 530028;4 天峨县林朵林场,广西河池 547300)
次生林在演替过程中常常形成一些已经死亡,但依然站立的树木即枯立木 (snag,standing dead tree)。它们属于森林生态系统的重要组成部分,显著地影响森林结构和功能[1-2]。枯立木的形成直接释放了林地空间,改变保留木的生长节律、林下更新、萌蘖、物种丰富度、多度和重要值[3]。枯立木是森林碳汇、土壤养分以及许多腐生生物、菌类的食物来源[2,4],还是一些小型野生动物造穴、栖居、觅食、避难和繁殖后代的重要场所[5-8],甚至被用于研究气候变化[9]。一定程度上,枯立木的大小和蓄积可被视为生物多样性指标和森林资源监测与评价的重要内容[10-11]。尤其在非木材生产的保护区域,枯立木在生物多样性保护方面的作用已经被广泛接受[12]。
在过去,许多研究将枯立木视为粗木质残体的一部分[13-14],重点关注其组分和数量特征[15-19]。一些研究涉及枯立木的空间结构,但并不关心枯立木本身,而是期望通过其特征获得群落的结构动态[20-22]。还有少数研究试图利用大样地数据来解析枯立木的空间结构特征[4,23-25]。但在林分水平上,次生林中枯立木的空间结构分析甚少。除了生态价值之外,它还具有重要的实践价值[26-27]。例如,指导异龄林经营中采伐木的选择。同时,当前研究主要集中在温带地区[10,13,17-18,20]以及少部分的热带地区[26,28-29],而在亚热带地区的探索很少[4,24],凸显了区域研究的不平衡。
除了树木坐标和树种之外,有关林分空间结构分析的另一个重要因素就是树木大小[29-31]。林分结构与功能 (如,垂直分层、水土保持)、生长与收获 (如,生长量、采伐量、经济收入)、物种多样性 (如,丰富度和多样性) 以及一些重要生态过程 (如,负密度依赖、非对称性竞争) 都与树木大小密切相关[32-35]。大量研究采用时空置换的方式探讨树木大小或分层与空间结构的关系[36-39],即将直径分布划分为几个跨度较大的径阶,每个径阶代表不同的生活史阶段。径阶划分的位置和数量通常带有很大主观性,其结果可能会漏掉一些很重要的信息[32]。同时,这些研究常常使用不同大小的直径,进一步降低结果的可对比性[8]。这一缺陷可能在多树种构成的天然异龄林中更加突出。另一种探讨树木大小和空间结构的方法是测量林分内包括了小树 [如,胸径 (diameter at breast height,DBH),DBH = 1 cm] 在内的所有个体,然后再分析它们取不同起测径 (即每木检尺的最小直径,minimum measured diameter,mmd) 时的空间结构变化[40]。最近一项研究表明活立木的起测径和空间结构是一种线性模式[31]。到目前为止,天然林中枯立木的起测径与空间结构的关系尚不清楚。
细叶云南松 (Pinusyunnanensisvar.tenuifoliaCheng et Law) 和数种栎类 (Quercus) 共生形成的松栎混交林是中国西南地区南盘江流域的地带性植被,有非常明确的起源、发展历程和经营历史记录[31,34,41-42]。本研究中,我们分析其枯立木的空间结构特征,目的是揭示林分水平上枯立木的大小和空间结构之间的关联。特别地,我们关注 (1) 枯立木的起测径是否与其空间结构有关;(2) 枯立木的空间结构特征;(3) 同类型林分枯立木空间结构的差异状况。
1 材料和方法
1.1 试验地概况及样地建立
试验地位于广西壮族自治区百色市乐业县广西雅长兰科植物国家级自然保护区一沟保护站,毗邻南盘江 (图1,见区位图部分)。该区域属于南亚热带气候,季节性干旱严重,春秋少雨,夏季漫长、多雨且炎热,冬无霜降及寒雪,年均降雨量约为800 mm,7月份平均气温为28.4 ℃,极端高温达42.5 ℃。区域内气温、降雨量随海拔升高和地貌改变而显著变化[43]。土壤类型由河谷地段的红褐土逐渐过渡到山原的褐红壤及高山的黄壤,整体上土壤贫瘠且石砾含量很高[31,34,41-42]。
该区域绝大多数细叶云南松原始林在1958-1960被皆伐,一些伐后迹地在无人干扰的条件下自我更新并发育形成了松栎混交林。林分垂直分层十分明显,常见的乔木层优势树种包括细叶云南松、栓皮栎 (QuercusvariabilisBl.)、青冈 [Cyclobalanopsisglauca(Thunb.) Oerst.] 和山合欢 [Albizziakalkora(Roxb.) Prain],常见的灌木层优势种主要有南烛 (VacciniumbracteatumThunb.)、金叶子 [Craibiodendronstellatum(Pierre) W.W.Smith]、余甘子 (PhyllanthusemblicaL.)、水锦树 (WendlandiauvariifoliaHance)、毛桐 [Mallotusjaponicus(Thunb.) Muell.Arg.var.ochraceo-albidus(Muell.Arg.) S.M.Hwang] 和红毛椿 (ToonaciliataM.Roem.)。林中偶见数种兰科植物以及粽叶芦 [Thysanolaenamaxima(Roxb.) Kuntze]、五节芒 [Miscanthusfloridulus(Lab.) Warb.ex Schum et Laut] 等草本植物[31,34,41]。
在2016-2018年期间,我们根据地形、植被组分以及外界干扰特征于一座大山中部和顶部建立了3块标准固定样地 (样地A,100 m × 60 m;样地B,80 m × 70 m;样地C,80 m × 80 m)。首先,在选择好目标林分之后,用全站仪 (NTS-372R10) 沿着坡面水平方向确定第一条边,再逆时针绕测出剩余3条边,并控制测量闭合差在1/400之内,获得样地外框。其次,用全站仪将样地外框划分为若干个 20 m × 20 m的小样方,在每个小样方的顶角埋入塑料管并用螺纹钢加固。最后,使用全站仪逐一定位每木的三维坐标 (x,y,z)。具体的定位对象包括活立木 (DBH ≥ 1 cm)、枯立木 (DBH ≥ 1 cm) 和枯倒木 (DBH ≥ 3 cm,长度 ≥ 3 m)。同时,我们检尺活立木树高、胸径、冠幅并对其挂牌和记录树种,也检尺了枯立木的胸径、树高,并依据根部萌蘖、树皮、干形、枯叶等特征判别其树种,还记录了枯倒木的腐烂程度。样方A中共有1 745活立木和265株枯立木,分别属于26树种。样方B中共有1 319活立木和168株枯立木,分别属27树种。样方C中共有1895株活立木和424株枯立木,分别属27树种[34]。本研究仅使用了有关枯立木的数据 (表1)。
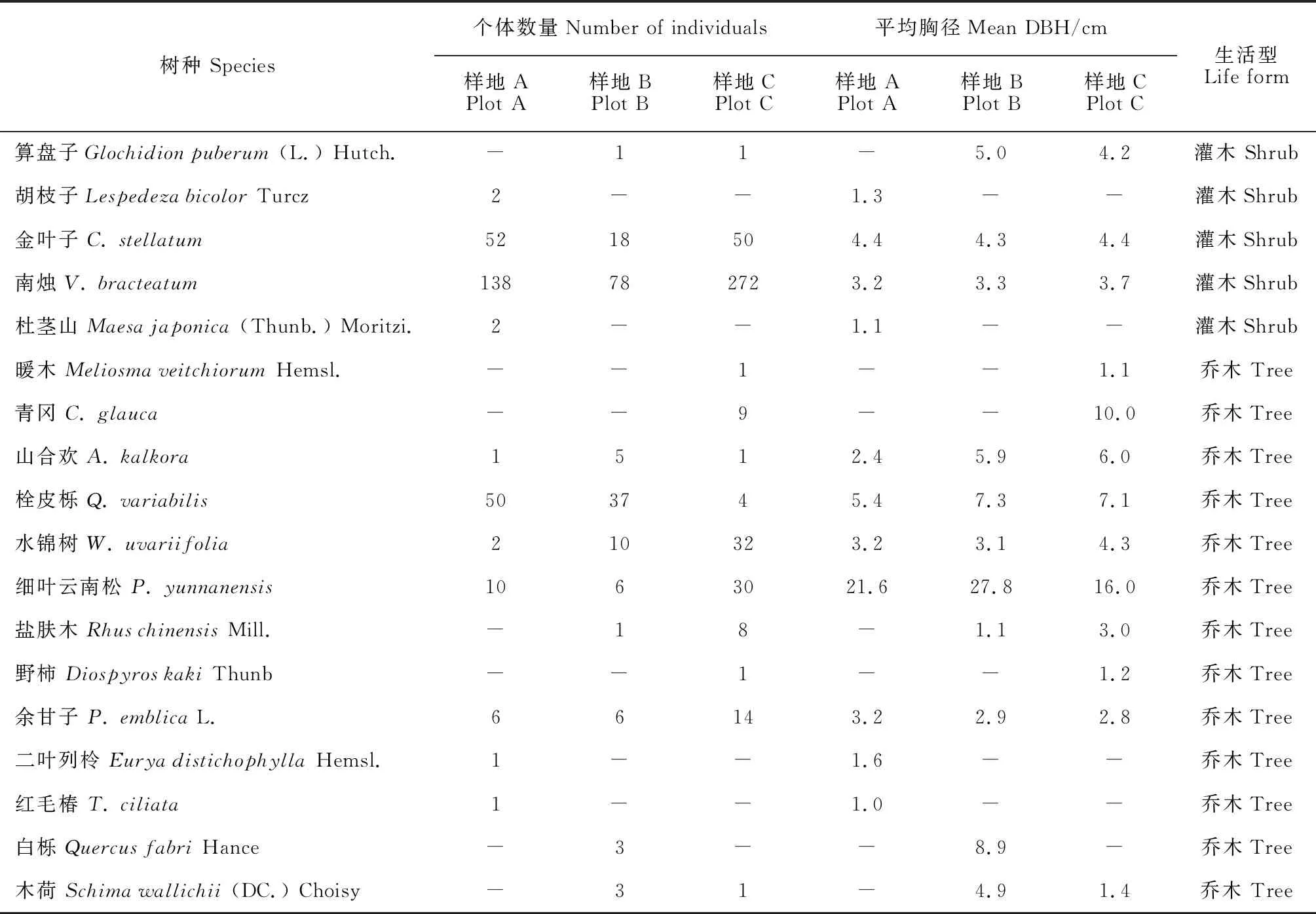
表1 南盘江流域松栎混交林中枯立木的树种信息Table 1 Species information of snags of pine-oak mixed forest stands in Nanpan River Basin
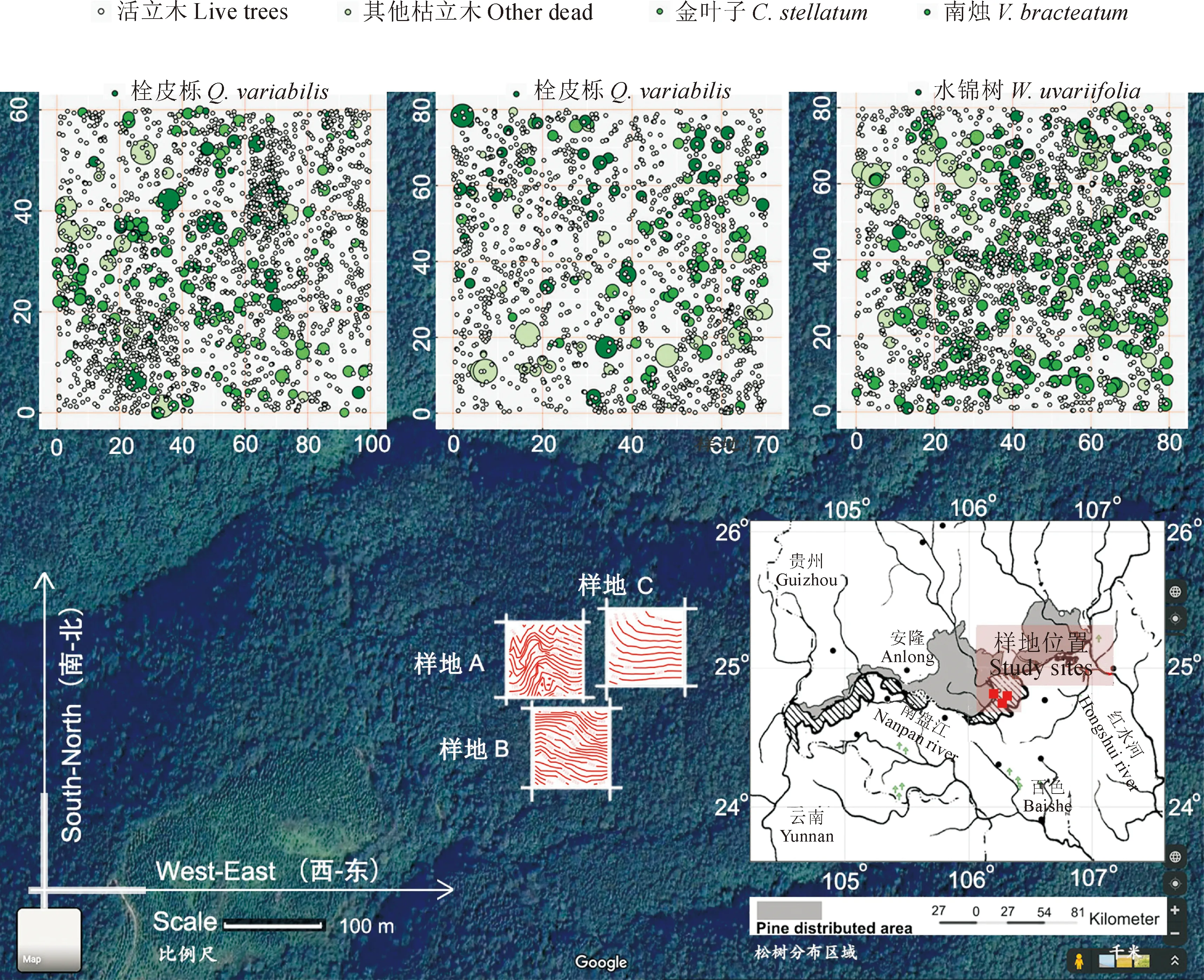
绿色点表示枯立木,点大小与胸径呈正比,灰色圆点表示活立木。图中部方形块状为样地A、B、C的具体位置,红色线条为实测等高线。右下角为区位图图1 试验地位置以及样方中树木的点分布The green and grey dots represent snags and live trees,respectively.The sizes of green dots are proportional to the diameter at breast height of snags.The squares in the middle of this graph represent the locations of quadrats and the red lines are measured contours.The location map is arranged at the lower right cornerFig.1 Study site and tree point pattern of the three quadrats
1.2 数据分析
我们将每块样方中的枯立木数据按照起测径从小到大划分为8组 (1-MaxBBHcm,2-MaxBBHcm,3-MaxBBHcm,……,8-MaxBBHcm),再分析每组数据的分布格局、标记特征以及距离分布。由于受到样方尺度的限制,我们直接假设它们不存生境异质性,也就是以完全空间随机 (complete spatial randomness,CSR) 为零模型,用双相关函数 [pair correlation function,g(r)] 分析枯立木的分布格局。我们用标记相关函数 [mark correlation function,kmm(r)] 分析标记特征“树种”的相似程度,即同种在空间上的聚集或隔离程度[20,34,44-45],用标记变差函数 [mark variogram function,γ(r)] 分析标记特征“大小”(本研究中指胸径) 的相似程度[33,46]。在上述3种分析中,考虑到林分年龄和树木大小,我们将相邻木可能发生交互作用的半径范围统一设置为r= 0-20 m,用蒙特卡洛 (Monte Carlo) 方法模拟每个尺度上的观测值999次并各取上下极值的2.5%作为包迹线,获得随机分布区间并对其拟合优度进行检验 (表2)。此外,我们还用距离累计分布函数Dk(r) 分析了枯立木的最近邻体 (k= 1,2,4,6,8,12) 在r= 0-10 m上的距离分布 (表2)。数据分析和图形制作在R语言的“spatstat”数据包[47]和“ggplot2”数据包[48]中完成。

表2 本研究中计算枯立木空间结构的公式Table 2 Formulas are used to calculate the spatial patterns of snags in this study
2 结果与分析
2.1 枯立木的分布格局
随着起测径和观察尺度的增加,3块样方的分布格局均由聚集转向并保持随机状态 (图2)。当mmd = 1~4 cm时,样方A中分布格局在多数尺度 (如,r= 0~20 m,0~9.5 m) 上表现出显著的聚集性 (图2,a-d)。但它在 mmd = 5~8 cm 的每个尺度上 (r= 0~20 m) 表现为随机分布,即落入随机分布区间 (图2,e-h)。样方B中的枯立木只在mmd = 1~2 cm时候的小尺度上 (r= 0~2 m) 呈现轻微聚集分布 (图2,i-j),而在其他情况下呈现随机分布 (图2,k-p)。样方C中枯立木分布格局的变化也只出现在小尺度上 (r= 0~1.5 m),即随着起测径取值的增加观测值由远离上包迹线逐渐靠近随机分布,而在其他尺度上保持随机状态 (图2,q-x)。
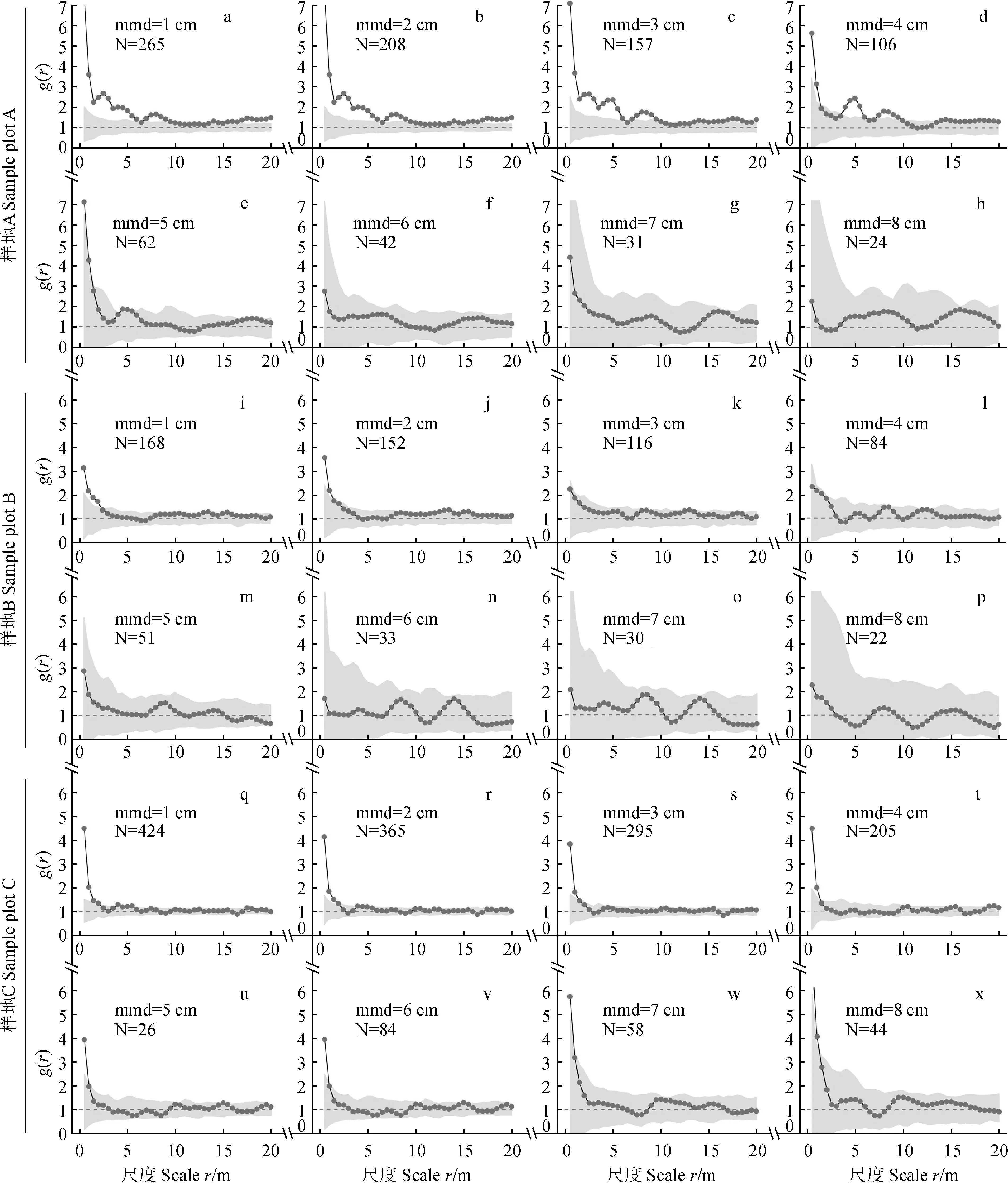
N代表株数,虚线为随机分布下的期望值,圆点黑色实线为观测值,灰色背景为95%蒙特卡洛模拟区间,也就是随机区间。当观测值落在模拟区间的上方、内部和下方时分别表示聚集分布、随机分布和规则分布。下同图2 南盘江流域松栎混交林中枯立木的分布格局N is the number of individuals.The dashed line stands for expectation under random distribution.The black solid line with dots dominates observed values and the grey area represented the 95% interval of Monte Carlo simulations (i.e.,random interval).The observed values fall above,inside and below the simulation interval,representing clustered,random and regular distribution,respectively.The same as belowFig.2 Distribution patterns of snags of pine-oak mixed forests in Nanpan River basin
2.2 枯立木的大小分布
3块样方中枯立木的大小分布也非常相似。当起测径的取值比较小时 (mmd = 1~2 cm),每块样方中枯立木的大小分布在小尺度上 (r= 0~1 m) 非常接近95%蒙特卡洛的下包迹线,即有小树聚集的趋势 (图3,a-b;图3,i-j;图3,q-r)。除了mmd = 6~8 cm,样方B中枯立木的大小分布在小尺度上 (r= 0~3 m) 也非常靠近下包迹线 (图3,n-p),其余部分的大小分布完全落入随机分布区间,也就是说枯立木的大小分布呈随机状态 (图3,c-h;图3,k-m;图3,q-x)。
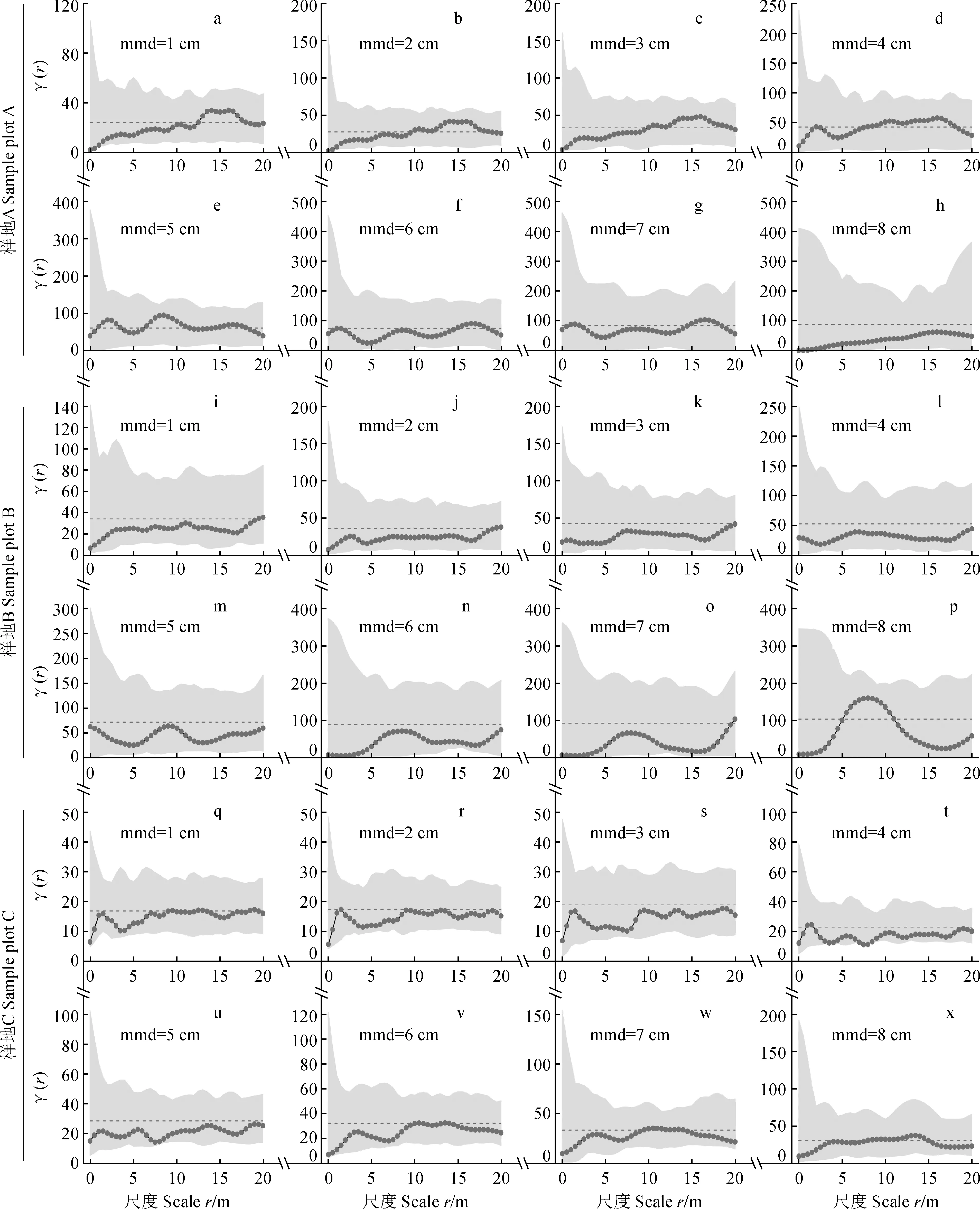
图3 南盘江流域松栎混交林中枯立木大小的空间分布Fig.3 Spatial patterns of tree size of snags of pine-oak mixed forests in Nanpan River basin
2.3 枯立木的树种分布
在样方A中,当起测径从1 cm增加到8 cm时,树种分布由聚集状态逐渐变呈随机状态 (r= 0~20 m,图4,a-h)。样方B中枯立木的树种分布也有相似特征,但它从起测径为5 cm开始,在所有尺度上 (r= 0~20 m) 都是随机分布 (图4,m-p)。当mmd = 1~3 cm时,在样方C中枯立木的树种分布在中小尺度上 (r= 0~7.5 m) 为聚集状态,而在剩余其他尺度上 (r= 8~20 m) 十分靠近随机分布 (图4,q-s)。随着起测径的进一步增加 (mmd = 4~8 cm),所有尺度上的观测值也进一步靠近并最终全部落入随机分布区间 (图4,t-x)。

图4 南盘江流域松栎混交林中枯立木树种的空间分布Fig.4 Spatial patterns of tree species of snags of pine-oak mixed forests in Nanpan River basin
2.4 枯立木的距离分布
随着起测径的增大 (mmd = 1~8 cm),3块样方 (A、B、C) 在10 m 范围内的相邻木逐渐减少。k值由12分别减少到只有2、4和6,第一株 (k= 1) 的距离分布累计概率也由1减少至0.708,0.636和0.818,而其他株 (k= 2,4,6,8,12) 的减幅更大,甚至到0。这种变化趋势在mmd = 5 cm之后尤其明显 (图5,a-x)。同时,它们的距离分布累计概率随k值的增加 (k= 1,2,4,6,8,12) 而降低 (图5,a-x)。也就是说,起测径的增加会减少相邻木的数量,同时还增加相邻木之间的距离。此外,样方中枯立木的株数也会对距离分布产生影响。当起测径取值相同时,同一尺度上株数多的样方比株数少的样方的累计概率可能更高,k值也可能更大 (图5,q-xvs图5,a-h;图5,q-xvs图5,i-p;图5,i-pvs图5,a-h)。
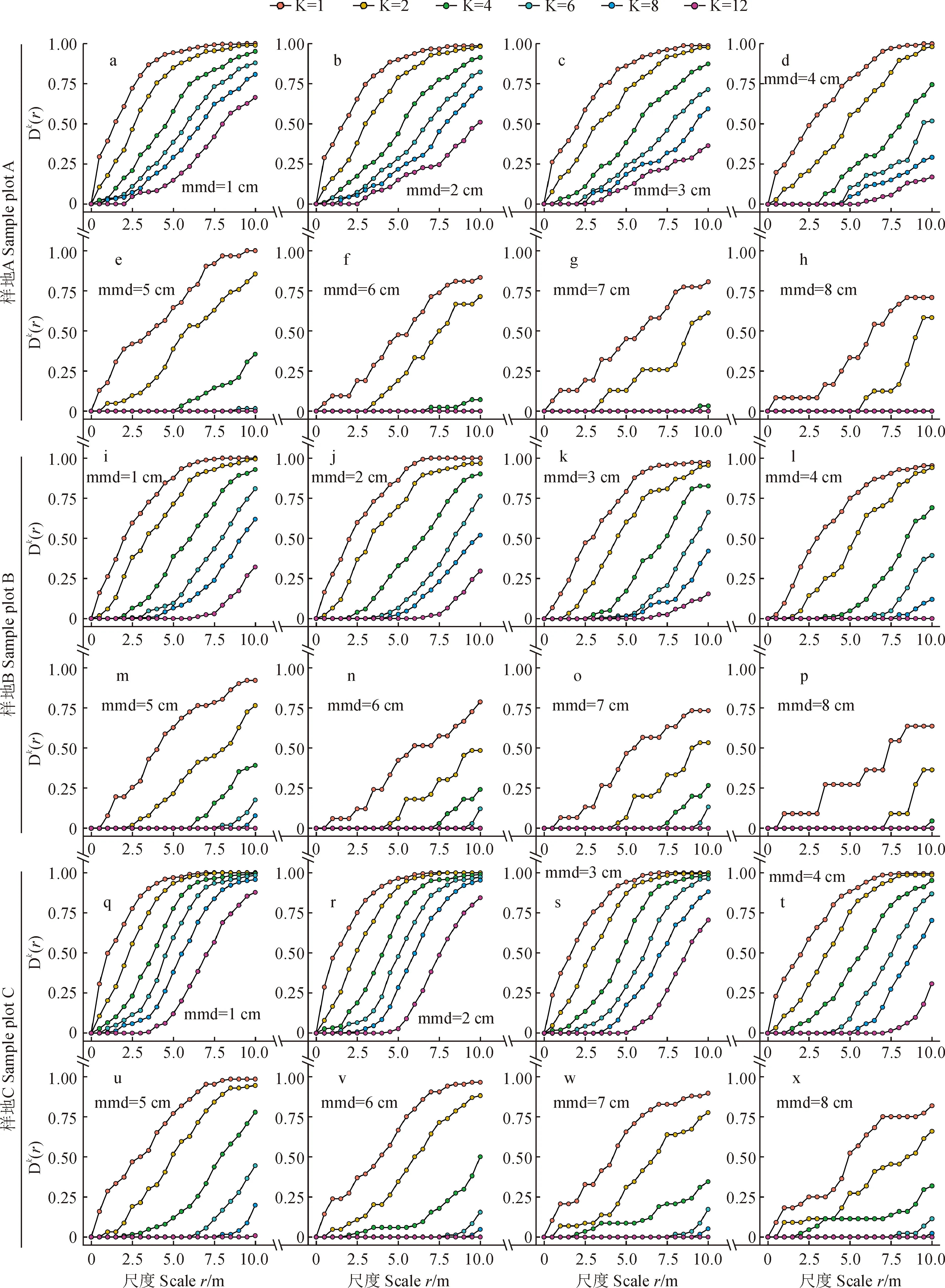
图5 南盘江流域松栎混交林中枯立木最近邻体的距离分布Fig.5 Distance distribution of the snags nearest neighbors of pine-oak mixed forests
3 讨 论
3.1 枯立木的大小与分布格局的关系
枯立木的形成是森林群落动态发生的直观体现[20],直接改变了分布格局。本研究中松栎类混交林中枯立木的大小与其分布格局密切相关,体现在起测径越小,个体数量越多,发生聚集性死亡的也越严重,说明小树是枯立木呈聚集分布的主要贡献者。同处中国亚热带地区 (广西弄岗自然保护区和湖南八大公山) 的两块大型固定样地 (15~25 hm2) 的起测径 (1 cm) 以及枯立木径阶 (分段) 和分布格局的关系与本研究相同[4,24]。在没有明显外源干扰的条件下,一些半天然或天然林中,树木的死亡率或数量随径阶增加呈负指数分布的下降趋势[3,49,21],也支持我们的结果。它常被理解为非对称性竞争的结果,树木越小承受的竞争压力越大,死亡风险越高。当起测径相对较大时 (如,5 cm),多数枯立木很可能处于林分上木层,与无梗花栎 (Quercuspetraea) 和欧洲山毛榉 (Fagussylvatica) 天然老龄林中的上木层的随机分布一致[50]。优势木的随机分布常被认为是种间、种内关系缓和的结果,是森林群落发展不可逆转的趋势并可能在将来演替中得以保持[51-52]。
在空间位置上,许多研究还认为枯立木的分布格局和数量与环境条件密切相关,尤其是地形、土壤和海拔。从沟谷到山坡,再到山脊,海拔高度增加,枯立木数量和分布格局强度亦增加[4,12,16,23]。本研究中各林分枯立木之间的数量和分布格局也与此相符。相对高处的聚集现象可从所谓的“土壤生态位”[24]、“生物组”[12]或是土壤水分干旱度[53]等角度进行解释和预测。此外,还有许多因素,如气候[54]、病虫[10]、林分年龄[55-56]、种子传播距离[28]、树种组分及其比例[12,17]等都可能影响枯立木的分布格局。进一步使用logistic函数分析单个因子的树木死亡率[10]或是建立多个因子与死亡率之间的logistic回归方程[54,57-58],可获知各因子在枯立木形成中的贡献。
3.2 枯立木的大小与标记特征的关系
枯立木形成的部分原因可能来自死前的内部竞争。在分布格局不变的前提下,可借助标记变差函数辅助判断它们死前可能受到的竞争压力。本研究中,标记变差函数展示了枯立木的大小分化并不显著,但个体越小,种内聚集程度越高。一个可能的解释是,该松栎混交林上木层是由高大的乔木树种组成,下木层由少数几个聚生的阳性灌木树种组成,灌木种间、种内的大小分化相对较弱,而上下层大小分化严重[34,41]。随着上木层郁闭度的增加,势必会引起林隙中的灌木集体死亡[34,59]。枯立木的树种组分和株数比例进一步说明了这种可能。许多研究还发现林内灌木树种的周期率 (turnover rates) 比乔木树种更短,也就是死亡较快[57-58]。这暗示植物生活型在一定程度上决定了个体的生存概率,亦支持了本研究结果。
当同时结合分布格局、大小、树种和距离分布时,我们发现松栎混交林中大量死亡的是聚生在小范围内的同种小树。这一现象除了可以被上述分层竞争解释之外,也很符合所谓的负密度依赖。即小范围内同种小树因资源竞争以及病虫侵害而导致高死亡率,从而降低同种聚集强度和促进物种共存。它被认为广泛存在热带、温带以及少数亚热带地区森林[20,60-61],尤其是林分演替早期或是树木生长的早期阶段[21,34,62]。然而,到目前为止很少有研究明确指出树木发生负密度依赖时的个体大小,我们的结果显示松栎混交林中5 cm以下的小树发生负密度依赖的概率很大。再者,负密度依赖在灌木和乔木之间的强度差异也未清楚,因此从多个角度探讨树木之间的交互作用是非常有必要的[59]。一项长达65年的固定样地监测显示,分布格局、树木大小和其他属性 (如,对光照依赖程度) 以复杂的方式紧密地相互交织,同时作用于林分发展过程。林分早期死亡的是聚生在一起的阳性小树,后期死亡的是规则分布的耐荫大树[63]。这也刚好与本研究结果相符。总之,枯立木的形成及其分布格局应是多种时空因素综合作用的结果,并可能在区域和林分间呈现复杂性[12,26,64]。
4 结 论
枯立木是森林生态系统重要的结构性和功能单位,探讨林分水平上枯立木空间结构有助于了解天然林动态。本研究综合地分析了中国西南地区南盘江流域松栎混交林中枯立木的空间结构,详细地展示了枯立木空间结构随着起测径的增加而发生的变化。本研究发现起测径显著地影响枯立木的分布格局、大小分布、树种分布和相邻木距离分布。换句话说,统计对象的数量差异会导致空间结构的分析结果不同,这种差异是一个渐变的过程。一方面,这意味着不同大小个体在森林生态系统过程中扮演不同角色。同种小树即阳性灌木树种易发生聚集性死亡,是“演替驱动”死亡的重点对象,而异种大树则趋向于随机死亡,符合早期次生群落演替的基本规律。另一方面,同类型林分枯立木空间结构的差异很小,需要使用相同的起测径进行对比分析。总之,本研究弥补了研究区域和内容上的空白,为粗木质残体调查、管理和以及生物多样性保护提供依据。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
阿来研究(2020年1期)2020-10-28
林业调查规划(2020年3期)2020-06-03
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
新世纪水泥导报(2016年1期)2016-07-01
中央社会主义学院学报(2016年2期)2016-05-04
小说月刊(2015年12期)2015-04-23
