
天门冬科广义铃兰族的染色体数目与核型进化研究
2021-09-16 14:49张籍元张代贵聂泽龙
西北植物学报 2021年8期
张籍元,张 成,孟 然,张代贵,聂泽龙,孟 盈
(吉首大学 生物资源与环境科学学院植物资源保护与利用重点实验室,湖南吉首 416000)
广义铃兰族(Convallarieae)包括白穗花属(Speirantha)、铃兰属(Convallaria)、万年青属(Rhodea)、吉祥草属(Reineckia)、长柱开口箭属(Tupistra)和蜘蛛抱蛋属(Aspidistra),共约200余种。铃兰族植物皆为多年生草本,除铃兰属冬季倒苗外,其余5属皆为常绿。铃兰族中仅铃兰属广布于北半球,属于典型的北温带分布,其余属集中分布于东亚和东南亚。中国和日本是铃兰族植物的多样化中心,该族70%以上的特有种分布在中国和日本[1]。铃兰族中大多数种具有重要的观赏和药用价值,如吉祥草属、万年青属和蜘蛛抱蛋属等,其中蜘蛛抱蛋属药理活性良好,民间应用较多,其在保护心脑血管、抗菌消炎、抗肿瘤和降血糖等方面具有广阔的应用前景[2]。
铃兰族传统隶属于百合科,主要特征为叶簇生在根状茎上,花葶沿叶基部侧生,花序为简单的总状花序或间断的穗状花序。Krause提出广义铃兰族,并将百合科分为12亚科35族[3]。在Krause的百合科系统中,铃兰族又被分为铃兰亚族(Coralarinae)和蜘蛛抱蛋亚族(Aspidistrinae)[3]。Hutchinson认为铃兰族具根状茎是百合科最原始的类群,于是又把Evrardiello和开口箭属(Compylandra)2个属纳入铃兰族内,并将广义铃兰族划分为狭义铃兰族(包括铃兰属、白穗花属、吉祥草属和夏须草属)和蜘蛛抱蛋族(蜘蛛抱蛋属、万年青属和长柱开口箭属)。Dalgren基于形态特征的研究结果,提出将狭义铃兰族、蜘蛛抱蛋族、黄精族(Polygonateae)与沿阶草族(Ophiopogoneae)共同组成铃兰科(Convallariaceae)的观点[4]。《中国植物志》把Campylandra并入长柱开口箭属,早期的系统发育研究通常也将夏须草属和Comospermum归置于铃兰族[5]。铃兰族作为一个自然的单系类群,也得到了分子系统学证据的支持,但该族的系统位置依然存在争议,分子证据支持将它从百合科分出置于铃兰科中,后来将铃兰科扩大到假叶树科(Ruscaceae),APG IV系统最终将该族归为天门冬科[6]。
染色体数目与核型反映了物种在染色体水平上的整体特征,细胞学研究有助于揭示物种间的亲缘关系和起源演化,为植物系统分类提供可靠的细胞学佐证[7]。国内外学者已经对铃兰族各属植物的染色体进行了大量的研究工作[8-29]。对于铃兰族的细胞学研究始于1961年Roy发现广东蜘蛛抱蛋(A.lurida)的染色体为2n=36[28]。中国对铃兰族的研究比较晚,杨涤清在1983开展了单种属百穗花的染色体数目为2n=38[23],随后许炳声又报道了吉祥草的染色体核型[22]。洪德元等[8]综合分析整个铃兰族各属的染色体结构发现白穗花属核型与蜘蛛抱蛋核型较一致,认为两属可能起源于一个共同祖先。杨涤清等[24]对栽培的万年青与野生的万年青作了核型研究,发现二者核型很接近,同时认为万年青与开口箭属的核型无明显区别,万年青的核型与铃兰属也较为相似,通过对长柱开口箭(T.chinensis)等种的核型研究得出了长柱开口箭属核型与吉祥草属相近的结论。黄锦岭在对长柱开口箭属的国产种类进行了详细研究后,发现长柱开口箭属的核型结构在属类变异相当大,不仅具有对称性较强的单型性核型,也有高度不对称的两型性核型[9]。黄锦岭等[12]依据该属与其可能的近缘类群的核型相似性进行比较,推断蜘蛛抱蛋属植物染色体原始基数为 x =19,但这与其外部形态结构尤其是花部式样及花粉形态演化趋势并不相不符。王任翔等[19]的研究发现蜘蛛抱蛋属染色体数目2n=36和2n=38比率基本相同,在染色体数目2n=36的种类中,花部式样的形态结构较为简单,可能代表其原始性状;而染色体数目2n=38的种类中,花部式样的形态结构较复杂和特化,可能代表其进化性状。刘安莉等[16]通过研究属内形态特征和核型进化结果同意上述观点,由此推测蜘蛛抱蛋的核型可能是由不对称向对称性增强的方向演化。
关于广义铃兰族内各属的分子系统发育关系一直存在争论,因为相对孤立,连续的变异性状少,所以难以判断性状的演化方向和属间系统发育关系。Yamashita等[29-30]基于trnK和rbcL对铃兰族进行了系统发育分析,结果表明狭义铃兰族不是单系群,支持铃兰族和蜘蛛抱蛋族共同构成广义铃兰族,铃兰属、百穗花属和吉祥草属处于基部位置,开口箭属Compylandra与万年青属形成第二支,Tricalistraochracea与长柱开口箭属Tupistra形成的一支与蜘蛛抱蛋属互为姐妹关系,同时提议将开口箭属归为万年青属,Tricalistra归入长柱开口箭属。黄丹妮通过对65种蜘蛛抱蛋属植物和12个外类群进行系统发育分析表明,蜘蛛抱蛋属植物是单系类群,与外类群中的长柱开口箭属关系最近,也支持将开口箭属并入万年青属[31]。
植物染色体的数目和形态等是最稳定的细胞学特征之一,在不同的种间染色体有不同的数目、核型及大小等的特征区别,植物染色体的核型特征也是表明该种系统演化位置以及和近缘物种之间关系的重要依据。染色体数目可以体现自然进化的进程及方向,为植物进化和系统发育研究提供有价值的信息[7]。铃兰族具有较为稳定的染色体数目以及明显的核型变化,是研究植物染色体进化非常理想的类群。关于铃兰族的研究涵盖了形态学、分类学、细胞学、孢粉学和植物化学成分等[1, 11, 12, 29-35],但是该族属间以及各属下种间的系统关系一直存在争议,本研究通过广泛收集铃兰族植物的分子数据来重建系统发育树,明确属间以及各属下种间的系统关系。因此本研究全面收集整理铃兰族染色体数目和核型,结合分子系统发育树推测本族植物的染色体演化规律,也为铃兰族属间关系及物种进化关系提供了细胞学证据。
1 材料和方法
1.1 细胞学实验
本实验5种铃兰族植物材料及采集信息见表1,凭证标本存放于湖南省湘西自治州吉首大学生物资源与环境科学学院标本馆(JSU)。在天气晴朗的上午9:00-10:00从活体上取1~2 cm长的幼嫩根尖浸泡于饱和对二氯苯水溶液中,于室温下预处理4 h,然后蒸馏水洗净,再用卡诺固定液I(无水乙醇∶冰醋酸 = 3∶1)于4 ℃冰箱中固定30 min,然后用1 mol·L-1HCl∶45%的冰醋酸 = 1∶1混合溶液在60 ℃的水浴锅中解离1 min,1%的地衣红染液染色,过夜,常规方法压片,显微镜下观察、拍照。每个物种选择10个以上有丝分裂中期染色体分散良好的细胞拍照,计数,最后选择最好的细胞图用Photoshop CS6软件上进行处理,核型分析及命名按照Levan等[36]的方法进行,核型类型按照Stebbins的标准处理[37]。此外,也用Arano提出的不对称系数As.K%来评估核型不对称性,As.K%值越高,核型越不对称[38]。

表1 实验材料来源与凭证标本
1.2 染色体数据收集与核型划分标准
本研究对铃兰族植物已报道的染色体资料进行全面的收集整理及核型统计,推测铃兰族植物染色体非整倍化和多倍化的进化规律和地位,这对全面了解铃兰族及其各属的起源和演化有积极的推动作用。染色体数据的收集主要根据《中国有文献报道的植物染色体数目索引》和“Index to Plant Chromosome Numbers”数据库进行查询,前者主要包括发表在国内外杂志和期刊的资料,后者包括全世界发表的所有植物的染色体索引,并且可在网站上(http://www.tropicos.org/Project/IPCN)查询,同时重点查阅最近分散在国内外各主要出版物上的文章等。
本研究对铃兰族的核型划分标准和方法参考Yamashita等[29]观点长度上有两种变异模式:单型(表现为染色体间在长度上没有明显的差异,呈逐渐过渡的趋势)和三型(由3组不同长度的染色体组成,并结合中期染色体的基数和长度变异模式将铃兰族分为:单型,染色体基数x=19的铃兰型;三型,基数为x = 19时为长柱开口箭型,基数为x = 18时则被称为蜘蛛抱蛋型。
1.3 基于系统发育的染色体进化分析
基于前期铃兰族的分子系统学研究[29],本研究选取代表铃兰科各个属和主要分支的64个种,天门冬属Asparagus作为外类群。选取常用于该类群系统发育分析的基因片段,包括4个叶绿体基因片段(rbcL、matK、psbA-trnH和trnL-F)和1个核基因片段(ITS)进行测序分析。本研究对天门冬科10个种进行PCR及测序,DNA提取、PCR引物及反应程序和测序方法参考Wang等[39]。同时从国际生物数据库(NCBI)下载了54个种的基因序列。所有用于本研究的类群凭证标本和GenBank登录号等详细信息见表2和3。分子序列首先用MUSCLE进行比对,然后用PhyED软件手动调整[40]。预分析表明叶绿体与核基因序列的结果不存在明显的冲突,并且与所有序列联合的结果非常相似,故我们最终将5个片段进行串联进行最大似然法(Maximum likelihood,ML)分析,并且不同序列片段采取单独的进化模型。ML分析利用RAxML7.2.6进行运算,进行1 000次Bootstrap搜索,最后得到一致树[41]。运算产生的树文件使用软件Figtree1.4.2进行可视化,最后把染色体数目和核型映射到系统进化树上来探讨它们的进化关系。
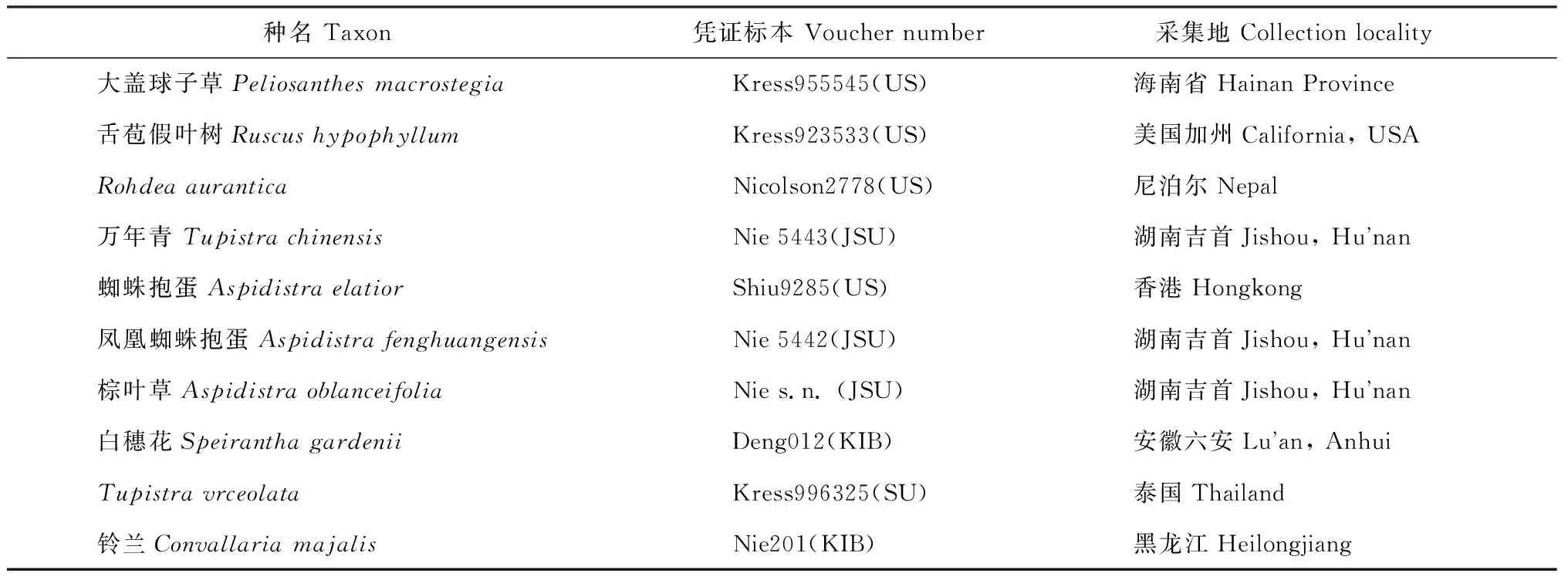
表2 实验材料来源与凭证标本

表3 本研究从GenBank下载序列的相关信息
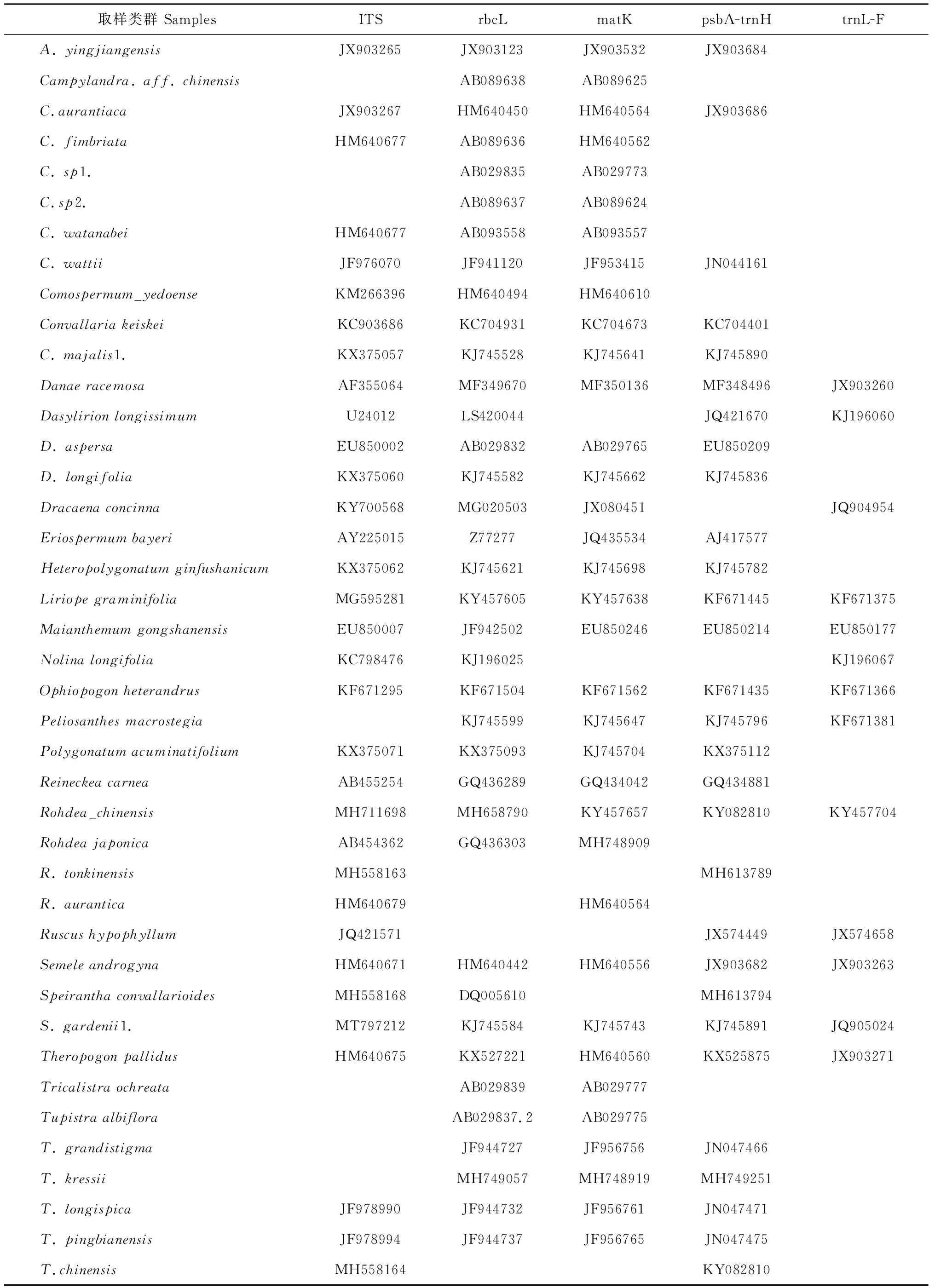
续表3 Continued Table 3
2 结果与分析
2.1 细胞学实验观察结果
本研究中铃兰族5个种植物的染色体核型公式及特征见表4,染色体形态和核型图见图1。结果表明这5个种中,除蜘蛛抱蛋染色体数目为2n=36外,其余4种均为2n=38,均为二倍体。这5个种中染色体均由m,sm和st的染色体构成,不对称性核型均为Stebbins’-2C型,核型不对称系数As.K%变化范围为65%~69%,并没有明显差别。但在染色体核型公式上有一定差别。其结果分别表述如下。

表4 5种铃兰族植物的染色体核型公式及特征

A.蜘蛛抱蛋;B. 海南蜘蛛抱蛋;C. 四川蜘蛛抱蛋;D. 凤凰蜘蛛抱蛋;E. 万年青. 标尺=5 μm图1 5种铃兰族植物中期染色体形态图A.Aspidistra elatior; B. A. hainanensis; C. A. sichuanesis; D. Aspidistra fenghuangensis; E. R. japonica. Bar=5 μmFig.1 Morphology and karyotype of somatic chromosomes of 5 species from the tribe Convallarieae
2.1.1 蜘蛛抱蛋AspidistraelatiorBlume蜘蛛抱蛋的染色体数目为2n=36,染色体基数为x=18,二倍体(图1, A)。在第1对(6.75~6.25 μm)与第2-9对(5.40~2.56 μm)之间在长度上有一个明显的差别(0.85 μm),第9对与第10-18对(1.66~0.80 μm)也存在一个明显的差别(0.90 μm)。核型公式为2n=36=16m+14sm+6st,最长染色体长度为6.75 μm,最短染色体长度为0.80 μm,最长染色体与最短染色体的比值为8.4,臂比小于2的染色体所占比例为61%。Stebbins’不对称核型为2C。核型不对称系数As.K%为67%。其核型表现为蜘蛛抱蛋型核型(三型,x=18)。蜘蛛抱蛋染色体数目有2n=36和2n=38两种,本种不同个体或不同居群间核型不一样,黄锦岭报道该种染色体数目为2n=38,其核型公式为2n=38=22m+4sm(2sat)+12st,第9对染色体短臂具随体[12]。虽然本研究的染色体数目与王任翔报道的一致,但在核型公式和随体方面却有所不同(2n=36=16m+4sm+12st(2sat)+4t,第10对染色体短臂具随体),具有2对端部着丝点的染色体[19]。本研究中并没有发现随体染色体的存在,也没有臂比值大于7的端部着丝点染色体(t)存在。可见,蜘蛛抱蛋在不同居群之间染色体核型和数目均有变异,是一个正在分化的类群。
2.1.2 海南蜘蛛抱蛋AspidistrahainanensisChun et F. C. How海南蜘蛛抱蛋染色体数目为2n=38,染色体基数为x=19,二倍体(图1,B)。核型公式为2n=38=20m+12sm+8st,在第1对(7.20~6.98 μm)与第2~6对(5.81~3.96 μm)之间在长度上有一个明显的差别(1.17 μm),第6对与第7~19对(2.89~1.10 μm)也存在一个明显的差别(1.07 μm)。最长染色体长度为7.20 μm,最短染色体长度为1.10 μm,最长染色体与最短染色体的比值为6.54,臂比小于2的染色体所占比例为66%。Stebbins’不对称核型为2C。长柱核型不对称系数As.K%为69%。其核型表现为开口箭型核型(三型,x=19)。海南蜘蛛抱蛋染色体数目有2n=36和2n=38两种报道,其中2n=38的占比为68%,该种与王任翔[19]报道的2n =36=20m(2sat)+14st+2t不一致,且我们也没有发现t型染色体。
2.1.3 四川蜘蛛抱蛋AspidistrasichuanensisK. Y. Lang et Z. Y. Zhu四川蜘蛛抱蛋的染色体数目是2n=38,染色体基数为x=19,二倍体(图1,C)。核型公式为2n=24m+6sm+8st,在第1对(7.48~6.95 μm)与第2-6对(6.09~3.95 μm)之间在长度上有一个明显的差别(0.86 μm),第6对与第7~19对(2.92~0.86 μm)也存在一个明显的差别(1.03 μm)。最长染色体长度为7.48 μm,最短染色体长度为0.86 μm,最长染色体与最短染色体的比值为8.7,臂比小于2的染色体所占比例为71%。Stebbins’不对称核型为2C。核型不对称系数As.K%为65%。其核型表现为长柱开口箭型核型(三型,x=19)。洪德元所报道的2n=22m+2sm+14st(2sat)核型中发现了0~5条数目不等的B染色体,但是本报道未发现B染色体。本报道与王任翔所报道2n=38=24m+4sm+10st(2sat),冉志华所报道的2n=38=24m+2sm+12st(2sat)及黄锦岭所报道的2n=38=20m+6sm+12st(2ast)颇为相似[12, 18-19],但本研究中并没发现有随体存在。
2.1.4 凤凰蜘蛛抱蛋AspidistrafenghuangensisK. Y. Lang凤凰蜘蛛抱蛋为武陵山区特有种,该种染色体数目是2n=38,染色体基数为x=19,二倍体(图1,D)。核型公式为2n=18m+14sm+6st,没有观察到随体,在第1对(7.86~6.95 μm)与2-6对(5.96~3.71 μm)之间在长度上有一个明显的差别(0.99 μm),第6对与第7-19对(3.06~0.88 μm)也存在一个明显的差别(0.65 μm)。最长染色体长度为7.86 μm,最短染色体长度为0.89 μm,最长染色体与最短染色体的比值为8.83,臂比小于2的染色体所占比例为55%。Stebbins’不对称核型为2C。核型不对称系数As.K%为65%。其核型表现为长柱开口箭型(三型,x=19)。与邓涛[42]所报道的2n=38=22m+6sm+10st核型公式有所差别,但核型不对称性一致。
2.1.5 万年青Rohdeajaponica(Thunb.) Roth本研究中染色体数目为2n=38,染色体基数x=19,二倍体(图1,E)。核型公式为2n=12m+24sm+2st,没有观察到随体,在所有染色体中不存在一个明显的差别,呈从大到小逐渐变化的趋势。最长染色体长度为4.99 μm,最短染色体长度为0.88 μm,最长染色体与最短染色体的比值为5.67,臂比值小于2的染色体数所占比例为53%。Stebbins’不对称核型为2C。核型不对称系数As.K%为67%。其核型表现为铃兰型(单型,x=19)。本结果与尚宗燕等[43]和Yamashita等[29]报道的染色体数目一致,但本研究结果的不对称性核型为更不对称的2C型,与它们报道的2B型不同。但在本研究中万年青属的最长与最短染色体之比(5.67)是5个种中最小的,同时臂比值小于2的染色体所占的比率也是最低的,仅为53%,总体来说万年青属的对称性比蜘蛛抱蛋属更为对称。万年青属与开口箭属、铃兰属和吉祥草属的核型颇为相似,均为铃兰型核型。
2.2 染色体收集结果
本研究共收集铃兰族中7属(包含Campylandra属)共81个种,253条染色体报道记录(表5)。这些报道主要集中在蜘蛛抱蛋属(170余种)内,其中187条染色体报道记录来自于蜘蛛抱蛋属,代表了48种,其染色体数目有两种(2n=36或2n=38), 染色体基数为x=18或19,绝大多数种为二倍体,少数为四倍体和六倍体;长柱开口箭属(29种)的记录有41条,代表21种,染色体数目均为2n=38,基数为x=19,均为二倍体;Campylandra属(16种)的报道有11条记录,代表了6个种,铃兰属7条记录代表全部2个物种,单种属万年青3条记录、吉祥草属2条记录、白穗花属1条记录,这5个属的染色体数目均为2n=38, 基数为19,均为二倍体。
表5 铃兰族已报道种的染色体信息



2.3 系统发育分析与染色体进化分析
我们把染色体数目和核型类型映射到铃兰族系统发育树上来探讨它们的染色体进化关系(图2)。分子发育树的结果表明铃兰族分为三支,第Ⅰ支为铃兰-白穗花分支,是铃兰族首先分化出来一支,它们都以x=19为基数;第Ⅱ支为万年表-吉祥草分支,染色体基数为x=19;第Ⅲ支为蜘蛛抱蛋-长柱开口箭分支,染色体基数为x=18和19。因此我们推测x=19铃兰族的原始基数,x=18是由x=19通过减少1对小染色体得到的。而在核型不对称性方面基部铃兰-白穗花分支大多具有比较对称的2B型核型,而处于进化位置上的蜘蛛抱蛋-长柱开口箭大多具有更不对称的2C或3C型,因此我们推测本属植物在核型不对称性方面是从对称向不对称的方向进化的。另外,结合从长度变异和染色体基数来看,基于Yamashita和Tamura的核型分类法,铃兰-白穗花分支具有铃兰型核型位于基部,其进化方向是从铃兰属型向长柱开口箭属型,再到蜘蛛抱蛋属型的演化。

单型. 指染色体间在长度上没有明显的差异,呈逐渐过渡的趋势,染色体基数x=19的铃兰型;三型. 指由3组不同长度的染色体组成,并结合中期染色体的基数和长度变异模式将铃兰族分为:染色体基数为x = 19时为长柱开口箭型,染色体基数为x = 18时为蜘蛛抱蛋型图2 基于叶绿体片段rbcL、matK、psbA-trnH 和trnL-F和核基因ITS铃兰族分子分子系统树Unimodal refers to the Convallaria type which shows Unimodal trend of gradual transition without obvious difference in length between chromosomes, with chromosome number x=19; Trimodal refers to the three groups of chromosomes with different lengths, combined with the base number and length variation pattern of metaphase chromosomes, can be divided into two groups: Tupistra type when the chromosome number is x=19, and Aspidistra type when the chromosome number is x=18Fig.2 The phylogenetic tree based on 4 chloroplast gene(rbcL, matK, psbA-trnH and trnL-F) and nuclear gene ITS of Convallarieae and allied tribe
3 讨 论
3.1 蜘蛛抱蛋属染色体核型特征
蜘蛛抱蛋属是铃兰族中拥有物种数目最多,形态变异最复杂多样,生长的地理环境也最为广泛的一个属[13]。结合前人报道的187条记录,加上本研究对蜘蛛抱蛋、海南蜘蛛抱蛋、四川蜘蛛抱蛋、凤凰蜘蛛抱蛋的实验结果,共191条记录代表48个物种。本属植物近年来发表了百余新种,目前为止已经包括170余种[16, 44-50],因此还有大量的物种急需开展细胞学研究。从现有的染色体数据来看,蜘蛛抱蛋属的染色体进化表现非常复杂,既有多倍化和非整倍化的演化,同时还兼具二倍体水平上的结构变异以及随体,B染色体的有无等特征。在蜘蛛抱蛋属中二倍体水平上的结构变异占主导地位,多倍化和非整倍性变异仅在少数种中发现。本属植物的染色体通常由3种类型的染色体组成:第1对为大型的中部着丝粒染色体(m),第2至第9或10对为中等大小的亚中部或亚端部着丝粒染色体(sm或st),其余为小型的中部或亚中部着丝粒染色体。李光照[2]把本属植物的核型分为两类:一类为2n=36=18(L+M)+18S(2sat),即9对大中型染色体和9对小型染色体;另一类为2n=38=16(L+M)+22S(2sat)。本研究中的蜘蛛抱蛋为第一种类型,由9对大中型染色体和9对小染色体组成,但四川蜘蛛抱蛋、海南蜘蛛抱蛋、凤凰蜘蛛抱蛋与李光照[2]发现的规律有所不同,仅有6对大中型染色体,其余皆为小型染色体,而且在本研究中4个种都没有发现随体染色体和B染色体。这表明染色体在分裂时可能受各种因素影响或在减数分裂时染色体互换和重组过程中发生了不同染色体之间的染色质重组,从而导致染色体的长度组成比例发生变化。不同物种之间,甚至在同一物种的不同居群间或同一居群不同个体间也会发生染色体结构上的变异,而且这种变异通常与外部形态变异相关。如小花蛛蛛抱蛋、四川蛛蛛抱蛋中就有不同居群间核型多态性与表型变异存在密切相关性[2]。
从现有的统计结果来看,染色体2n=36的类群比2n =38的类群通常都多1 对中型染色体,少2对小型染色体,这可能是由于2对小的染色体融合成为1对中型染色体。染色体融合与断裂事件在植物进化的过程中起到了一定的作用,这在许多类群中也有发现,如陈耀华等[51]发现在石蒜属植物中有染色体的融合的情况发生。周勇等在研究3种禾本科植物时发现水稻的12条染色体经过2次染色体融合事件最终形成了高粱的10条染色体,随后又经历了染色体融合事件,最终形成了玉米的10条染色体[52]。当然在蛛蛛抱蛋属中2n=36和2n=38之间的变异机制有待进一步研究。
3.2 铃兰族染色体核型进化模式
近年来基于分子系统发育结果来探讨染色体进化机制已经在很多类群中广泛应用[53-58]。中国学者大多认为蜘蛛抱蛋属为二型性核型,分为大中型染色体和小型染色体两种[2, 8, 12-17, 19, 26],Yamashita发现第1对大的中部着丝粒的染色体在蜘蛛抱蛋属中是非常显著的特征,因此把第1对大的染色体单独分出来,因此从长度上来说定义为三型更为合理,并结合分子系统发育分析结果,把铃兰族植物的核型分为3种类型:x=18的三型染色体仅在蜘蛛抱蛋属中发现,因此命名为蜘蛛抱蛋核型;x=19的三型染色体命名为长柱开口箭核型;x=19的一型染色体为铃兰型。本研究基于4个叶绿体基因和ITS核基因的联合分析,重建铃兰族代表类群的系统发育关系,并结合染色体数目和核型分析发现:白穗花属和铃兰属形成一支位于整个族的基部,它们均具有x=19的基数,染色体在长度上呈逐渐变小的趋势,也就是铃兰型;族中第二个分支由吉祥草属、开口箭属和万年青属组成,它们核型特征与第一分支相似,也是铃兰型;第三支由蜘蛛抱蛋属、长柱开口箭属和Tricalistra组成,其中蜘蛛抱蛋属独立构成一支,而Tricalistra嵌在长柱开口箭属中。在第三个分支中蜘蛛抱蛋属的核型既有长柱开口箭型也有蜘蛛抱蛋型,结合分子系统发育结果来看,铃兰族中处于基部的类群以及与之亲缘关系最近的开口箭属的染色体基数均为x=19,因此推测在蜘蛛抱蛋属中染色体的原始基数应为x=19,而x=18是后来演生出来的基数。换而言之,铃兰族中染色体核型进化的趋势是从铃兰型演化为长柱开口箭型,再到蜘蛛抱蛋型,本研究结果与Yamashita等[29]基于rbcL和trnK两个叶绿体片段分析的结果一致,但本研究中选取更多的序列联合分析,得到更高的支持率,3个分支的演化关系更为清楚,但要解决蜘蛛抱蛋属和长柱开口箭属下种间的关系还需要更多分子数据的进一步研究。
刘安莉等[16]认为染色体基数x=18的种类大多数种的花被阔钟状或钟状,根状茎小于10 mm以下,花梗较长,花被片反卷,花被内基部无明显突起,雄蕊着生位置明显高于柱头或与柱头等高,柱头较小,表面结构及纹饰简单,果实表面光滑等特征是原始的类型;染色体基数为x=19的类群中,有诸多演生性状,如花的形态和结构较复杂,花被坛状,雄蕊着生位置低于柱头,柱头较大,表面形状及纹饰特化成蘑菇状等特征。通常认为植物的外部形态特征的变异是由基因和环境变异共同决定的,受光照、温度、水分、海拔、营养物质等环境因子影响,具有很强的表型可塑性[59-62]。因此推测某一个特征是否为原始性状,不能仅从结构简单或复杂来判断。现代分子技术手段为推测物种演化方向提供了最为可靠的证据,因此利用分子系统发育手段综合考虑它与近缘类群在外部形态、染色体核型特征的区别与联系,从而对性状和核型的演化方向作出科学合理的判断。
3.3 铃兰族染色体核型不对称性进化模式
中国学者对于蜘蛛抱蛋属的核型不对称性也进行了大量的探讨,他们普遍认为蜘蛛抱蛋属植物花的形态特征与核型进化相关,花形态结构越简单,核型不对称程度越强,形态结构越复杂特化,核型不对称程度就越弱,因此推测在蜘蛛抱蛋属内核型不对称性是由不对称性向对称性的方向演化[2, 16]。在铃兰族中Stebbins’不对称性核型为2B、2C,少数为3C,仅在长梗开口箭中发现有1A。结合分子系统发育结果来看,位于铃兰族最基部的铃兰属为2B型,其次吉祥草属、万年青属也为2B型,其他种类基本为2C型,因此可以推测铃兰族的染色体核型不对称性总体表现为由对称向不对称性增强的趋势。
3.4 铃兰族的染色体数目与核型演化趋势与意义
结合本研究对铃兰族各属的细胞学报道以及对本族染色体资料进行了统计分析,结果表明在铃兰族的染色体基数与系统关系方面,铃兰族的染色体数目变化和分子系统发育相对一致。铃兰族的染色体基数是x = 18、19,根据中期染色体的基本数目和形态将铃兰族分为单型和三型2种核型类型。单型主要包括吉祥草属、万年青属、Campylandra、铃兰属和白穗花属,染色体基数均为x=19,而三型包括蜘蛛抱蛋属、长柱开口箭属和Tricalistra,染色体均为x=19,部分蜘蛛抱蛋为x=18。我们认为中期染色体的三型染色体核型是由单型染色体核型衍生而来。然而,需要通过更多分类群的系统发育分析以及更多类群更精确的核型分析来进一步确认。铃兰族绝大多数种是2n=36、38的二倍体,其核型具有稳定性,存在少数多倍化现象,铃兰族染色体核型的差异正是环境因素和结构变异共同作用的结果。蜘蛛抱蛋属植物的染色体倍性变异多数为2x和4x两种,泰国蜘蛛抱蛋为2n=114,为染色体基数x=19的六倍体。西林蜘蛛抱蛋和十字蜘蛛抱蛋两种为四倍体种2n=4x=76,且为同源四倍体,植物多倍化可能是对恶劣生存环境适应的结果,在蜘蛛抱蛋属中发现多倍体的类群在外部形态方面与干旱环境相适应有一定的相关性[13]。整倍体变异和非整倍体变异不是该属植物演化的主要趋势,蜘蛛抱蛋绝大部分类群是二倍体,这表明该属植物的系统发育主要是在二倍体水平上进行演化。
王任翔等[19-20]认为蜘蛛抱蛋属植物的演化与高等植物核型的一般进化趋势向不对称性增强的方向演化的规律正好相反,也与该属的近缘属长柱开口箭属中的核型演化趋势相反。但是根据蜘蛛抱蛋属植物染色体不对称系数可以看出,染色体数目不同的两组间染色体不对称指数的变化相对较小,而x= 18组的染色体不对称指数明显低于x=19组。我们的结果表明,x=19为族内染色体原始基数,x=18为派生基数,而x=18是从x=19演化而来。通过对染色体长度研究表明本族染色体低基数的x=18由x=19通过染色体融合或丢失进化而来,而核型则由比较对称的单型向不对称的三型演化。所以祖先类群的核型对称性高于派生类群,蜘蛛抱蛋属的核型可能是向对称性增强的方向演化。因此,从目前已有的核型资料看,铃兰族植物符合Stebbins所提出的核型不对称性增加的趋势。长柱开口箭属中绝大部分种的染色体属于一型,染色体长短是递增排列的较为对称,伞柱长柱开口箭染色体中短染色体较明显,但是中长染色体与长染色体的长度很接近,染色体长度不对称因此属于二型染色体,伞柱开口箭这个种是能够代表长柱开口箭属向蜘蛛抱蛋属演化的一个证据,而从染色体的类型来看开口箭属比蜘蛛抱蛋属更为原始[10]。由研究结果看来蜘蛛抱蛋属和长柱开口箭属之间的核型演化关系和系统发育关系很明显较亲密,两属确定为姐妹类群。本研究揭示铃兰族由高基数向低基数减少以及单型向三型演化的趋势,按照染色体长度铃兰族核型演化趋势由对称向不对称演化,核型在属间和属内都有很大变化。铃兰族的核型的进化趋势表明铃兰属为拟原始特征,由此衍生出长柱开口箭属,进而衍生出蜘蛛抱蛋属。
猜你喜欢
四川劳动保障(2021年9期)2022-01-18
无锡职业技术学院学报(2019年4期)2019-12-27
中学时代(2019年1期)2019-11-13
小学生必读(中年级版)(2018年6期)2018-09-05
小学生导刊(2018年1期)2018-03-15
小学生学习指导(低年级)(2017年9期)2017-08-07
作文新天地(初中版)(2016年12期)2017-01-05
作文与考试·小学高年级版(2016年3期)2016-04-20
留学生(2015年5期)2015-12-26
中学语文(2015年35期)2015-12-08
