
克里雅河尾闾绿洲胡杨种群年龄结构对地下水埋深的响应
2021-09-16 14:37史浩伯师庆东周小龙万彦博
西北植物学报 2021年8期
史浩伯,师庆东,3*,戴 岳,3,周小龙,3,万彦博,彭 磊
(1 新疆大学 资源与环境科学学院,乌鲁木齐 830046;2 绿洲生态教育部重点实验室,乌鲁木齐 830046;3 新疆大学干旱生态环境研究所,乌鲁木齐 830046)
植物种群的年龄特征是数量特征中的重要指标之一[1],对研究种群的未来发展动态与群落的更新演替均能起到良好的分析预测作用[2-4],在探讨某一生境内的植物在生长发育过程中所呈现出的特征时,往往可以根据现阶段的年龄结构给出其在现状方面的良好解释[5],有助于深化在种群尺度上对植株生长特性的认知,了解植被格局的形成机制,这对于受不良环境条件胁迫以及处在濒危阶段的植物保护工作有着重要的指导意义[6]。同时,在植物格局的形成过程中并不仅受生存环境的影响,生物学特性也是植物群落在构建过程中不可忽视的因素[7],同样可能影响种群的结构动态和繁殖过程,而竞争关系作为自然界同种或异种个体之间争夺共同资源的常见现象,在植物功能性状的形成、多样性格局的演变以及共存机制方面均具有一定的驱动意义[8-9]。
胡杨(Populuseuphratica)是大陆性干旱气候条件下的常见树种,广泛分布于中国西北以及中亚等干旱地区,耐寒耐涝,对恶劣环境的抗性极强,是荒漠区特有的森林资源与关键种,在遏制荒漠化进程与风沙防治方面具有重要作用[10]。目前针对胡杨的研究以塔里木河流域与黑河流域居多,在宏观尺度上的研究主要集中于胡杨的生活史特征、空间分布格局与动态等方面[11-13],同时在生理特征与遗传分化等微观尺度上也取得了丰硕的成果[14-15],多元化的研究方向与研究成果的叠加足以说明胡杨对于荒漠生态系统的重要性,因此在对干旱生境演变与荒漠植物种群演替的研究过程中,胡杨往往具有较强的指示作用。
克里雅河是中国新疆南部的第四大水系,其尾闾达里雅布依深入塔里木盆地腹地300 km,是极具科学研究价值的“原始绿洲”[16],当地种群结构简单,自我更新与繁殖能力均受到了干旱条件的制约[17],而胡杨作为当地的主体树种,在维持绿洲生态系统稳定,减缓沙漠腹地荒漠化对绿洲侵占进程方面具有决定性意义。本研究为阐明地下水埋深制约条件下沙漠腹地胡杨种群的生长状况及演替动态,通过小尺度试验拟解决两个科学问题:(1)当前环境条件下的胡杨种群呈现出怎样的更新特征?(2)不同地下水埋深条件下的胡杨种群年龄结构有何差异?通过上述分析旨在说明地下水因子对胡杨年龄结构与更新特征的影响,为宏观尺度上的绿洲胡杨林保护及生态修复等方面提供参考依据。
1 研究区概况
达里雅布依位于世界第二大沙漠塔克拉玛干沙漠腹地,是中国塔里木盆地南缘第二大河流克里雅河的尾闾绿洲,该绿洲因其与世隔绝、交通不便而受到干扰因素较少,基本保持了自然条件下的状态,具有相对“原始”特征。达里雅布依地理位置为38°16′~38°37′ N,81°05′~81°46′ E,海拔为1 100~1 300 m,总面积约342 km2。该区域为暖温带干旱沙漠气候,年降水量不超过20 mm,潜在蒸发量达2 480 mm,年均温12.1 ℃,昼夜温差大,气候极端干旱。绿洲内部具有网状河相沉积特征,干枯河道分布于绿洲内部,土壤以沙土为主,天然植物群落则主要由胡杨、柽柳(Tamarixchinensis)、骆驼刺(Alhagisparsifolia)、芦苇(Phragmitescommunis)等物种组成[18]。
2 研究方法
2.1 野外试验
课题组于2018年结合生境异质性在绿洲内布设了19口地下水观测井,并于2019、2020年7月前往达里雅布依绿洲进行野外实地踏查,通过典型样地法选取胡杨种群分布集中且林分保持良好的地段,以所选择的观测井为中心布设11个20 m×20 m的植被样方,样地设置如图1所示。在运用手持GPS对样方进行定位后开展胡杨种群逐木测量工作,利用卷尺、游标卡尺等工具对胡杨的胸径、株高、冠幅等基本数据进行测量,并统计种群的个体数量,同时基于地下水观测井内的HOBO水位自动记录仪获取地下水埋深数据。
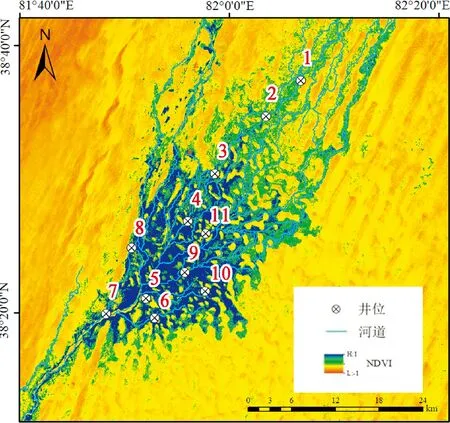
1~11.代表样地位置图1 研究区样地位置示意图1~11 represent the location of the selected sample plotsFig.1 Sketch map of study area
2.2 数据处理与分析
2.2.1 地下水埋深梯度划分采用K-均值聚类方法划分地下水埋深梯度,将其梯度分为三个等级(图2):0~3 m的地下水埋深为第一梯度等级(A);3~6 m的地下水埋深为第二梯度等级(B);6~9 m的地下水埋深为第三梯度等级(C)。
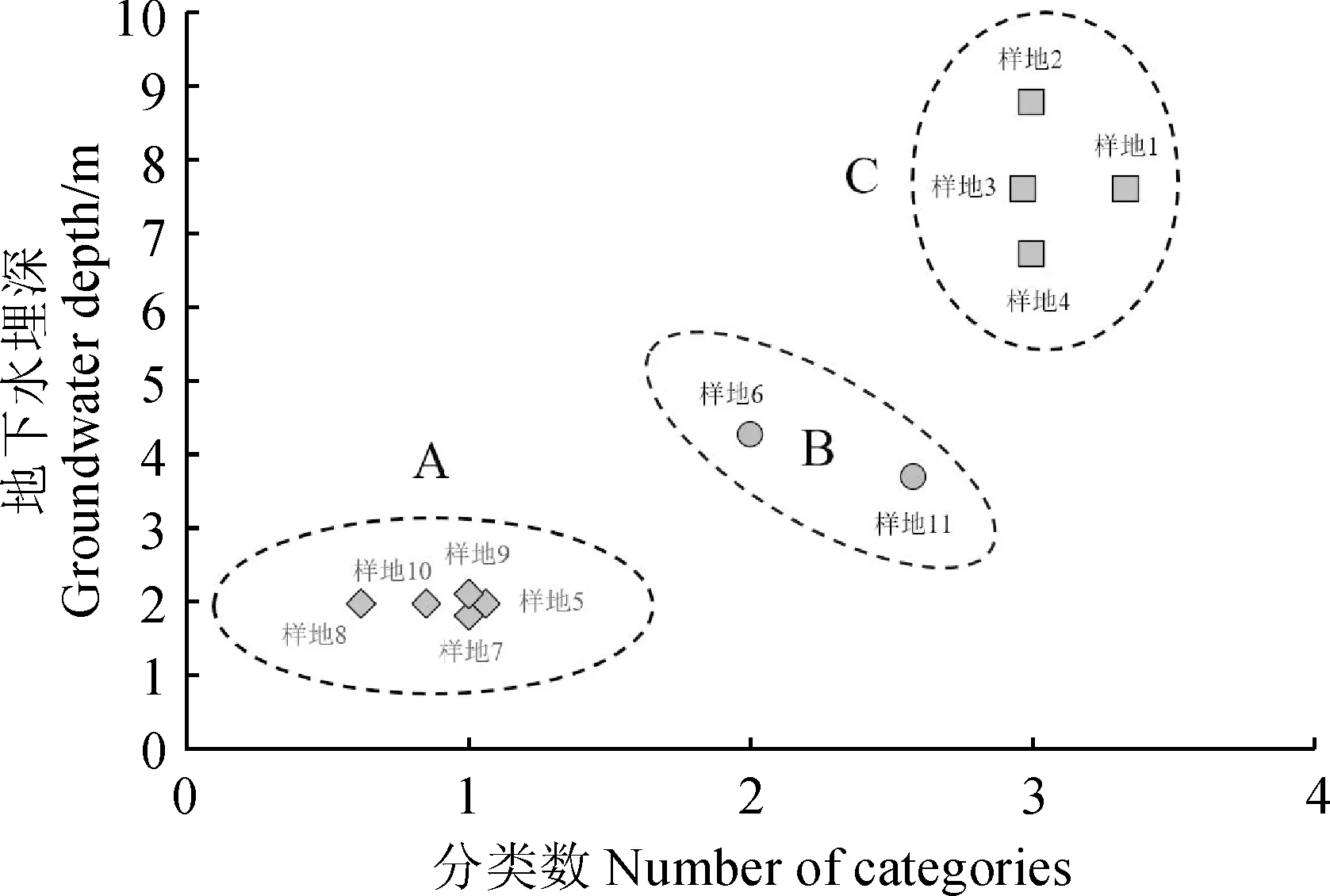
图2 研究区绿洲地下水埋深梯度聚类结果Fig.2 Gradient clustering results of groundwater depth
2.2.2 龄级划分结合胡杨的生活史特点可将其划分为12个龄级[19],分别是Ⅰ龄级: 0 cm<胸径≤4 cm;Ⅱ龄级: 4 cm<胸径≤8 cm;Ⅲ龄级: 8 cm<胸径≤12 cm;Ⅳ龄级: 12 cm<胸径≤16 cm;Ⅴ龄级: 16 cm<胸径≤20 cm;Ⅵ龄级: 20 cm<胸径≤24 cm;Ⅶ龄级: 24 cm<胸径≤28 cm;Ⅷ龄级: 28 cm<胸径≤32 cm;Ⅸ龄级: 32 cm<胸径≤36 cm;Ⅹ龄级: 36 cm<胸径≤40 cm;Ⅺ龄级: 40 cm<胸径≤44 cm;Ⅻ龄级: 胸径>44 cm。同时结合胡杨生活史特点,可将胡杨的年龄等级进行汇总,进一步分为幼龄林(Ⅰ~Ⅲ)、中龄林(Ⅳ~Ⅶ)、成熟林(Ⅷ~Ⅺ)和老龄林(Ⅻ)4个等级,以便降低年龄结构划分的复杂程度。
2.2.3 年龄结构动态变化分析胡杨种群的年龄结构是一种静态的生态学特征,应通过量化手段将其呈现出时空尺度上的直观动态变化,文中选用相邻龄级个体数量动态变化指数(Vn)与种群整体年龄结构动态系数(Vpi)进行种群发育动态分析[20],方法如下:
式中,Sn与Sn+1分别表示第n与第n+1龄级种群的个体数量,Vn存在值域[-1,1],当Vn取值为正、负、零时分别代表该种群在两个相邻龄级的个体数量呈增长、衰退、稳定的结构动态关系。Vpi是由不同年龄级间的动态变化指数对个体数Sn加权分配得来的,乘以权重后因为并不存在最大年龄级,所以将k这一最大龄级排除,最终Vpi的结果也存在正、负与零3种不同类型的值,且意义与Vn一致。
2.2.4 年龄结构动态预测运用一次移动平均法能对以年龄级为生长周期单位的胡杨种群数量动态进行有效预测[21],方法如下:
式中,n代表期望预测的时间长度,Mt代表n个生长周期后t龄级种群的数量大小,Xk是当前k龄级种群的大小,本研究分别对未来2个和5个龄级时间后的胡杨种群年龄结构进行预测。
2.2.5 生长空间竞争指数模型已有研究表明,干旱地区的胡杨冠幅与胸径的生长状况与地下水埋深有显著关系[22-24],因此如何建立以水资源作为限制条件的竞争关系模型极为重要。本研究基于马履一提出的生长空间竞争指数模型[25]进行推导,以此适应荒漠生态系统胡杨的生长特性。已知生长空间竞争指数(growth space competition index,GSCI)公式如下:
GSCI=CASi/BAi
由于胡杨生长受环境条件的限制导致树冠形状呈多样化,为更准确地说明其径向生长情况,故选用投影面积(SA)代替原始公式中的树冠表面积(CAS),具体公式为:
SA=CAS= π· [(a+b) / 4]2
式中,CASi代表竞争木的树冠表面积,BAi代表竞争木的胸高断面积,a为东西方向的冠幅,b为南北方向的冠幅。GSCI值越小,反映单位面积需要的树冠面积越小,其竞争能力越强,同时作为树冠覆盖程度的无量纲度量,是一种相比于生态过程更注重竞争结果的度量方法。
3 结果与分析
3.1 绿洲胡杨种群年龄结构与动态分析
达里雅布依绿洲内的胡杨整体年龄结构呈“不规则形”(图3)。根据本研究拟定的龄级结构划分标准,中龄林、成熟林在现有植株数量中占比最少,分别为0.19与0.14;其次是幼龄林数量,占比0.25;Ⅻ龄级以上的老龄林数量最多,占比0.40。总体来看,在绿洲胡杨种群的年龄结构中老龄化现象明显,在中龄林与成熟林两个年龄组分中存在断层现象,但幼龄林组分尚具规模,其补员作用能够在一定程度上缓解种群在更新方面的压力。
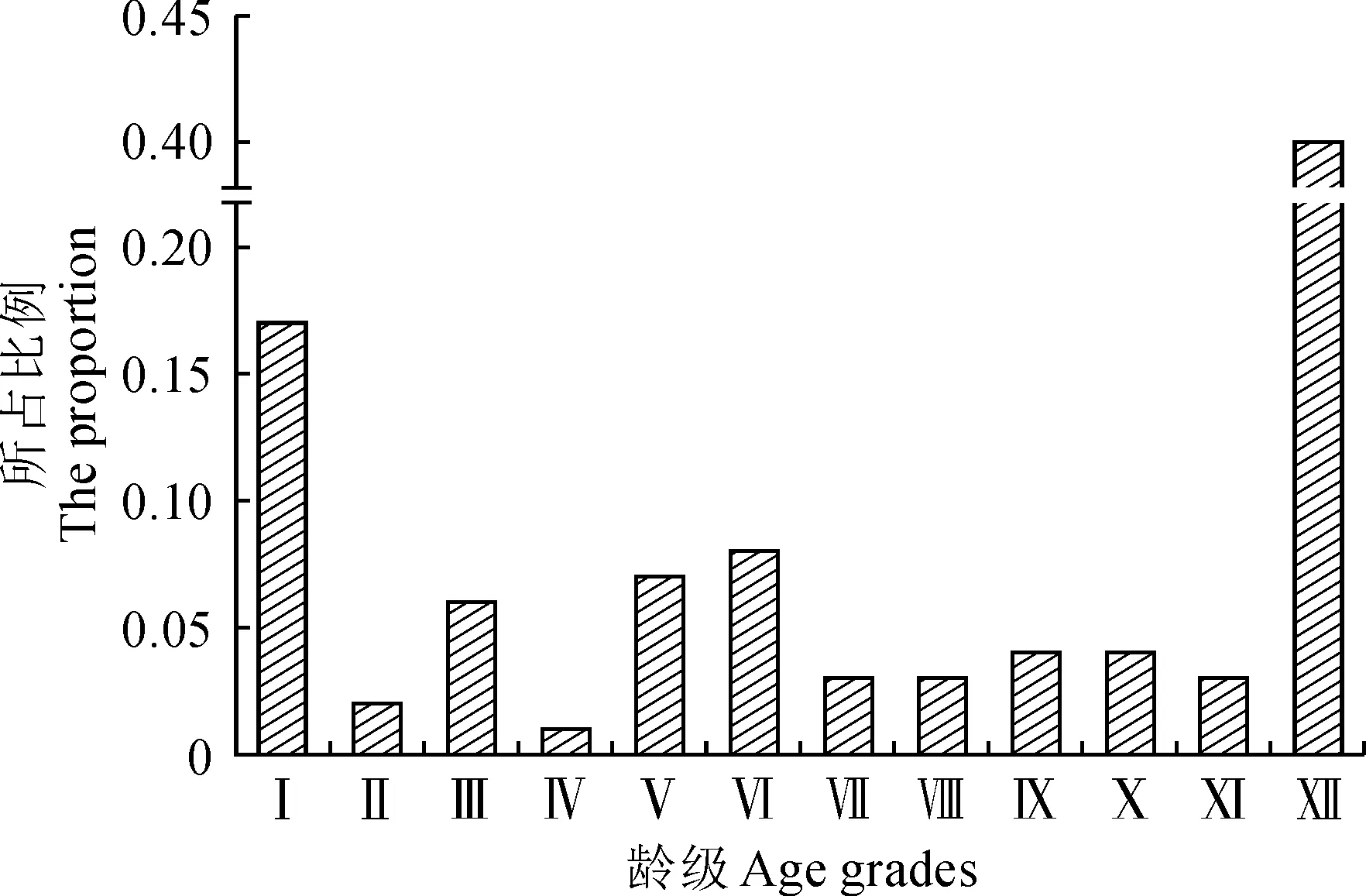
Ⅰ~Ⅻ.龄级;DBH. 胸径; Ⅰ. 0 cm
将种群动态量化后所得出的相邻年龄级之间的动态变化指数(表1)表现出较强的随机性:当前绿洲内的Ⅰ龄级的胡杨幼苗数量具备更新基础,与Ⅱ龄级之间呈增长的结构动态关系,与之相似,Ⅲ和Ⅳ、Ⅵ和Ⅶ、Ⅹ和Ⅺ三对相邻龄级之间同样存在增长关系;而在Ⅱ和Ⅲ、Ⅳ和Ⅴ、Ⅴ和Ⅵ、Ⅷ和Ⅸ、Ⅺ和Ⅻ五对龄级之间的相邻年龄结构动态呈现出衰退趋势;Ⅶ和Ⅷ、Ⅸ和Ⅹ两对相邻龄级的种群动态则呈稳定关系。绿洲胡杨整体种群的年龄结构动态指数Vpi=29.55%,其数值趋近于零,说明种群整体现阶段较为稳定,虽具有一定的增长趋势但并不明显。
3.2 年龄结构与生长空间竞争指数变化关系
胡杨个体的生长空间竞争指数与龄级的线性拟合关系呈现一定的规律性(图4),拟合方程为y=66.79±4.28x-2.08±0.6,R2=0.69。这一结果说明胡杨种群的年龄结构与个体竞争能力之间存在幂函数关系,表明依据胸径特征划分出的龄级能够在一定程度上说明物种的个体竞争能力,即随着胡杨的龄级的上升,个体的竞争能力会逐步加强,但加强的幅度持续变低。
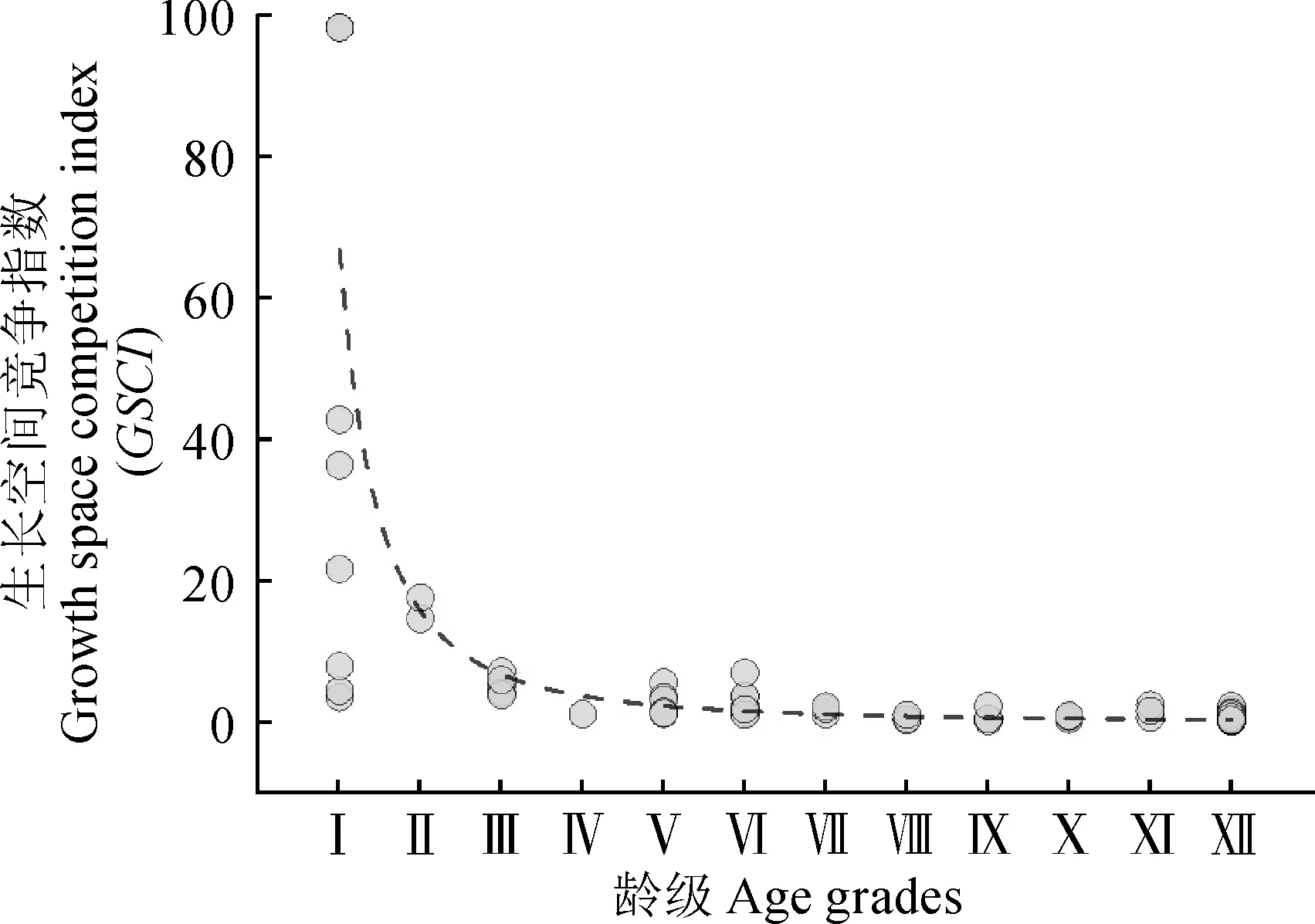
图4 研究区绿洲胡杨龄级与生长空间竞争指数变化关系Fig.4 Relationship between P. euphratica age grade and growth space competition index
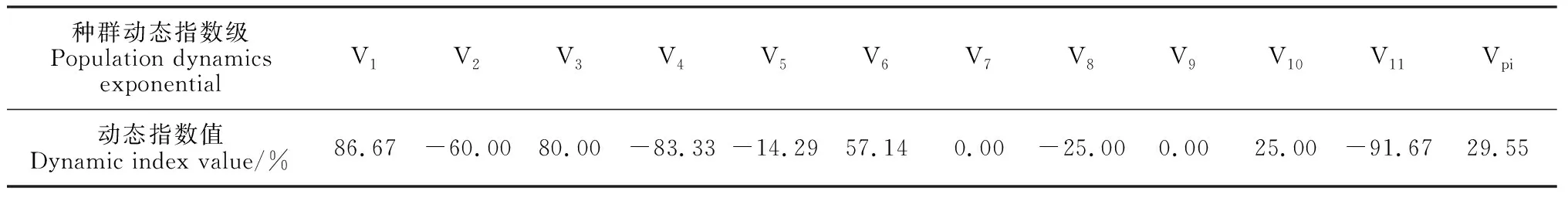
表1 胡杨种群龄级结构动态指数
3.3 地下水埋深梯度下的种群年龄结构变化
胡杨种群年龄结构与个体竞争能力存在较好的线性关系,因此可以通过不同地下水埋深梯度下的具体生长空间竞争指数来判断生境对种群生长的维持情况。如图5所示,在地下水埋深处于0~3 m的第一梯度(A)时,胡杨种群的生长空间竞争指数的变异系数平均值为1.09,该梯度下胡杨个体竞争能力的离散程度与其他两个梯度相比处于中等水平,各样地种群具体的年龄结构可归为两种类型,一类是仅存在胸径大于44 cm的老龄林群落,另外一类则是年龄结构复杂,不同龄级个体均有出现的多龄级群落;在地下水埋深处于3~6 m之间的第二梯度(B)时,竞争指数的变异系数平均值为2.176,离散程度较高,虽各自样地均仅包括2个龄级,但总体的误差值高达2.167,说明了这一地下水埋深梯度下胡杨个体的竞争能力存在较高的可波动范围;在地下水埋深处于6~9 m的第三梯度(C)时,竞争指数的平均变异系数为0.634,且离散程度极低,原因在于该梯度下各样地的胡杨种群均无幼龄林出现,成熟林与老龄林居多,占个体总数的84.85%,且现存个体龄级之间次序紧密,在年龄结构方面呈现出典型的老龄化特征。
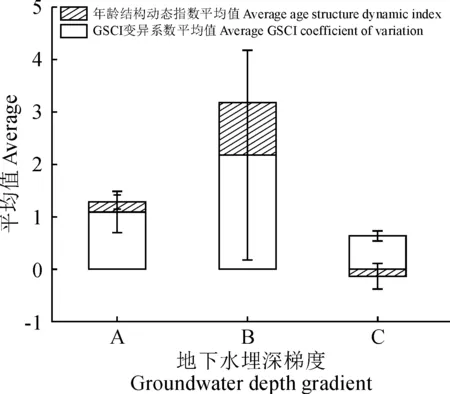
图5 研究区绿洲不同地下水埋深梯度下胡杨年龄结构动态与个体竞争指数变异特征Fig.5 Age structure dynamics and individual competition index variation characteristics under different groundwater depth gradients
而在胡杨种群生长动态变化特征方面,不同地下水埋深梯度下的种群年龄结构动态也呈现出一定的差异(图5)。当所处环境的地下水埋深位于第一梯度(A)时,动态指数的平均值为0.194,标准误差为0.135,不同区域的种群生长动态存在稳定与增长两种不同的水平;而在地下水埋深处于第二梯度(B)时,种群的年龄动态呈现极为明显的增长特征,且不存在误差值;在地下水埋深处于第三梯度(C)时,年龄结构动态指数平均值为-0.134,该梯度下的绝大部分区域均呈现出衰退的种群年龄结构特征,而仅存的具有增长趋势的区域其增长动态也并不明显,但正是这种正负关系的存在导致了该梯度范围内出现了误差值较大的现象。
3.4 时间序列上的种群动态预测
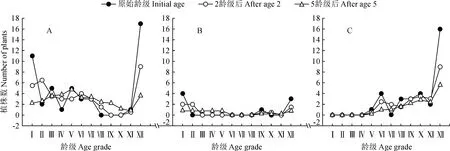
绿洲胡杨种群的年龄结构动态受所处地下水梯度的差别影响而有所不同(图6)。在地下水埋深0~3 m的第一梯度(图6,A)中,随着时间的推移老龄林与幼龄林均有所降低,中龄林逐渐成为林地中的重要组分,该区域的种群年龄结构逐渐变得稳定,但直至5个龄级时间后依然存在增长的趋势;在地下水埋深3~6 m的第二梯度(图6,B)中,2个龄级时间后其结构依然为增长型,5个龄级时间后的年龄结构逐渐趋于稳定,且以幼龄林与中龄林为主,但该环境在初始条件下的个体数目较少,对时间序列上预测的波动幅度产生了一定的影响;在地下水埋深6~9 m的第三梯度(图6,C)中,干旱的环境条件下幼龄林的不足导致胡杨年龄结构呈持续性的衰退趋势,加强对幼苗的抚育工作是维持种群持续发展的主要手段。
A.0~3 m;B.3~6 m;C.6~9m图6 研究区绿洲不同地下水埋深梯度下胡杨种群动态预测结果Fig.6 Prediction results of population dynamics under different groundwater depth gradients
4 讨 论
4.1 水文动态对胡杨种群更新演替的影响
克里雅河在维吾尔语中有“漂移不定”的意思,流域的形成依赖于昆仑山远古冰川与融雪补给,受季节性洪水的影响改道频繁[26],其尾闾达里雅布依绿洲的网状河相也因此不能保证各条河道均有稳定的地表水经过[27]。本研究中第一资源梯度中的样地均处于绿洲东南部,即克里雅河主河道与绿洲网状河道的交汇处,在理论上该区域的多数河道均应依靠季节性洪水存在稳定的地下水埋深补给。第二资源梯度下的样地则主要分布在绿洲南部与中部,最南端的河道属于独立的水系支流,干涸程度较重,而中部地区的河道已与主干河道存在一定距离,季节性洪水对这两处区域的地下水的补给强度远不及第一梯度区域,但其之所以呈现出典型的增长型年龄结构群落,很可能是6~9月的克里雅河丰水期产生的河水漫溢作用影响了胡杨实生苗的更新,提高了幼龄林的个体数量[28-29]。而第三资源梯度的区域主要分布于绿洲北部,即使是洪水期对地下水的补给作用也相对乏力,幼龄林在低水位的情况下无法生长,受此影响导致胡杨林表现为衰退型群落。综上所述,浅层地下水作为荒漠植被生长的重要资源,是促使其更新演替的动力源泉,对胡杨的龄级结构组成有重要影响。
4.2 年龄结构与种内关系的互作机制
本项研究中选取的生长空间竞争指数是基于干旱区胡杨生长属性特征进行量化,在忽略外界干扰情况下最大程度反映不同个体维持自身基本属性所需的地下水埋深阈值。由图4和图5可知,伴随着龄级的增长,植株个体的竞争能力也随之上升,进而使同一区域内的种内竞争现象愈发激烈。而地下水作为荒漠区植被生存所依赖的关键因子,对维持物种的种内竞争关系具有重要作用。在地下水埋深处于0~3 m的情况下能够同时维持单一老龄林的强烈竞争作用与多龄级条件下复杂的竞争机制;在地下水埋深处于3~6 m的情况下,能够维持龄级较少的增长型群落,但这种对竞争关系的维持作用存在一定的波动阈值,不能同时满足多龄级的存在;而在地下水埋深处于6~9 m的情况下,种群年龄结构已处于衰退状态,激烈的竞争关系则会进一步排斥低竞争优势的幼龄林,相关研究表明,随着龄级增长至老龄林,林木已摆脱被压状态,因此低龄级林木成为了竞争关系中的主要被排斥对象[30],在无低龄级与充足地下水补给的情况下,这种衰退情况会持续加剧,直至区域性的种群完全丧失。
4.3 时间序列上的胡杨种群动态
如果以地下水埋深三种不同的梯度条件作为限制因子研究种群动态,能够发现不同区域的种群在面临环境条件差异时,表现出的动态特征也有所不同。地下水埋深第一梯度(A)下的胡杨种群在演替过程中表现出的增长特征极为明显,在5个龄级时间后其年龄结构依旧完整,表现出较强的活力;而地下水埋深第二梯度(B)很可能受到了试验过程中样地数量的影响,导致初始条件下的胡杨个体数量较少,但在时间序列预测上依然表现出了明确的动态特征,即在5个龄级时间后趋于稳定。受地下水埋深的影响下,该种群的演替特征应处于高水位埋深与低水位埋深的过渡阶段,即虽然无法满足年龄结构的完整,但依旧具有一定的更新能力;地下水埋深的第三梯度(C)缺失了幼龄林这一整体组分,同时老龄林所占比例极高,导致在没有补员作用下的胡杨林呈现出明显的衰退趋势。上述分析中所用的时间预测分析手段具有较高的准确性,适用于多种限制条件下的预测[31],同时其弊端也很明显,预测的结果仅基于现状进行研究而往往忽视了生物的扩散与再定殖过程[32],因此在现实的发展动态中很可能受到生物因素的影响,但在胡杨林的保护与可持续发展方面,则可以根据预测的动态结果制定针对性的幼苗抚育、生态修复等工作,这对于促进胡杨林的天然更新,维持种群的稳定性均具有重要意义。
5 结 论
(1)达里雅布依绿洲胡杨老龄林数量居多,其次是幼龄林。整体年龄结构动态存在微弱的增长趋势,但不同生境之间的差异较大,多数区域的龄级更新依旧乏力。
(2)胡杨个体的竞争能力与径级存在幂函数关系,受控于对地下水埋深差异导致维持的种内竞争机制存在一定的差异,即随着地下水埋深的增加,所能维持的竞争关系愈发简单,而个体竞争能力较低的幼龄林比例也随之降低。
(3)随着时间的推移,0~6 m的地下水埋深依旧能够保障胡杨种群正常的更新演替,而大于6 m后的地下水埋深则会使胡杨种群丧失幼龄林这一组分,呈现出明显的衰退趋势。
猜你喜欢
湖南生态科学学报(2022年4期)2022-11-11
数学物理学报(2021年6期)2021-12-21
金桥(2021年11期)2021-11-20
北广人物(2020年47期)2020-12-09
河南农业大学学报(2020年2期)2020-05-22
数学物理学报(2019年5期)2019-11-29
山西林业科技(2019年1期)2019-05-27
创新作文(1-2年级)(2018年6期)2018-01-22
甘肃林业(2016年4期)2016-11-07
数学年刊A辑(中文版)(2016年1期)2016-10-30
