
野大麦–内生真菌共生体作为生态草的潜力浅析
2021-11-03 02:42翟纡润陈振江魏学凯陈泰祥段廷玉李春杰
草业科学 2021年9期
翟纡润,陈振江,魏学凯,陈泰祥,段廷玉,李春杰
(兰州大学草地农业科技学院,甘肃 兰州 730020)
我国草地面积辽阔,分布广泛,种质资源丰富,各类天然草地有4.0 × 108hm2,约占国土面积的2/5[1],栽培草地面积约2.1 × 107hm2[2]。主要分布在西藏、陕西、甘肃、宁夏、内蒙古、新疆、青海、四川和黑龙江等13 个省份,其中西藏的天然草地面积最大,内蒙古栽培草地面积最大[3]。但在当代社会,经济的快速发展推动着城市化和工业化的进程,造成的环境污染问题日益严重,伴随着水土流失、过度开垦以及自然灾害等,我国草地退化程度不断加深。20 世纪60年代到21 世纪初,各类草地单位面积产草量降低了近40%, 严重地区下降近70%[4],草地的生态服务功能和生态质量持续下降,不仅严重限制畜牧业的发展,破坏草地生态环境,而且使得动物、微生物乃至人类的生存环境也受到严重威胁,因此保护生态环境刻不容缓。
生态草是以生态建设为主要目的的用草,生态草建设是对草地盖度在60%以下的生态脆弱区,选择合适的生态草种,采取“五字(封、造、补、种、改)”治理方针,来遏制草地“三化(沙化、荒漠化和盐碱化)”,达到恢复和改善植被的目的[5]。我国吉林省生态草建设已取得显著成效(表1),羊草(Leymus chinensis)、紫花苜蓿(Medicago sativa)、柽柳(Tamarix chinensis)和碱茅(Puccinellia distans)等作为生态草种被广泛应用,它们均具有较强的耐盐碱性、耐旱性,繁殖能力强,根系发达,对退化草地有显著的改善作用[6-9]。目前,土壤盐碱化已经成为世界性问题之一,我国盐碱化土壤的面积约1.0 × 108hm2,其导致农业落后、单产水平低,制约着经济发展[10]。前人研究发现,星星草(Puccinellia tenuiflora)[11]、碱茅[9]和碱蓬(Suaeda salsa)[12]等耐盐植物在滨海盐渍土种植两年后的土壤脱盐率分别为22.9%、28.5%和26.83%,土壤有机质、全氮、速效磷和速效钾等含量均呈上升趋势。因此,种植耐盐植物对盐渍化土壤有明显的修复效果,同时也取得了较好的经济效益。
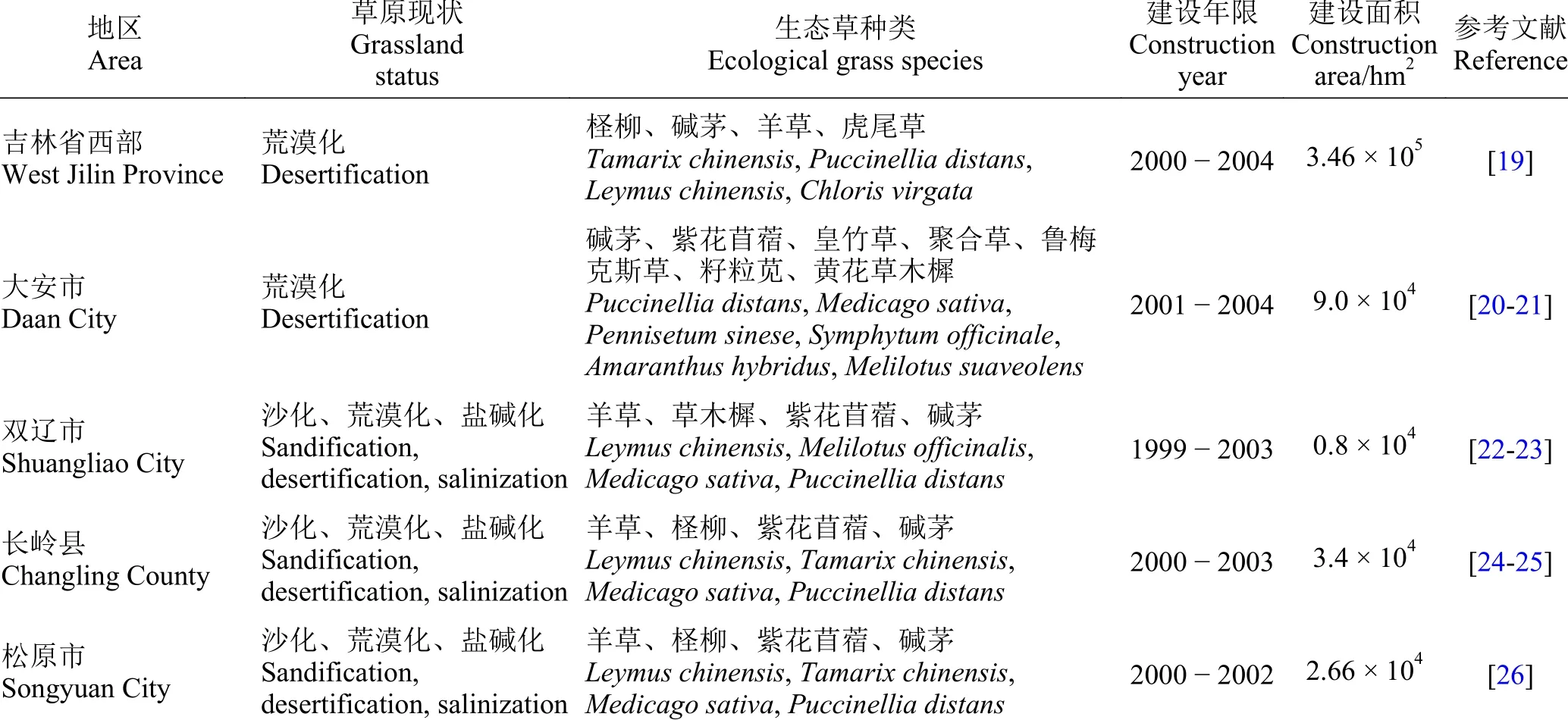
表1 我国吉林省生态建设成果Table 1 Ecological construction achievements in Jilin Province, China
野大麦(Hordeum brevisubulatum)在我国北方地区的轻盐渍化的草甸上分布广泛,其具有根系发达、分蘖株丛密,青草产量高,抗逆性强,适应性广等特点[13],其中对于野大麦耐盐性的研究相对较多,在植株发育的不同时期用NaCl、NaCl-Na2CO3等盐溶液或复合盐溶液进行处理,虽然对野大麦的生长发育有抑制作用,但也表现出较强的耐盐性[14-15]。除了用来放牧家畜、调制干草和青贮饲料等发挥饲用价值外,在改良盐渍化土壤、防风固沙、调节气候、美化环境以及维护生态平衡等方面均做出巨大贡献[13]。禾草内生真菌是指在禾草内度过全部或大部分生命周期,而禾草不显示外部症状的一类真菌,主要包括香柱菌(Epichloë)属真菌[16]。很多研究证明,禾草内生真菌可以提高宿主对生物胁迫和非生物胁迫的耐受性[17-18],这对野大麦–内生真菌共生体作为生态草提供了理论基础。本研究对野大麦–内生真菌共生体作为生态草的潜力进行阐述,以期充分利用野大麦–内生真菌共生体这一种质资源,服务于栽培草地建设、退化草地改良等方面,以加快生态环境改善的步伐。
1 野大麦的优良特性、利用价值及其与内生真菌的关系
野大麦亦称野黑麦、短芒大麦草、莱麦草,是禾本科大麦属多年生草本植物。野大麦属于强分蘖型禾草,生长繁茂,再生能力和自我繁殖能力极强,可快速形成优势种群[27],抑制毒害杂草的入侵。其种子成熟期较为一致,落粒性强,不易收集,但有利于自然繁殖[13],这也是作为生态草的优良特性之一。野大麦抗逆性强,可以在低洼积水处或干旱条件下生长,在极低温(−40 ℃)环境下能够安全越冬,表现出较强的耐涝性、耐寒性和耐旱性[28-29]。野大麦喜生于轻盐渍化的草甸、林缘等潮湿环境中,主要分布在我国华北、东北、西藏、青海、内蒙古等地[30],有研究显示,野大麦的耐盐能力与芨芨草(Achnatherum splendens)和碱茅等相似,其对盐碱土壤的改良效果仅次于碱茅(表2)。因而,野大麦可以作为水土保持植物,对盐渍化土壤进行利用、改良或在低湿低洼地区建立栽培草地等方面具有重要意义。野大麦生长发育快,花后营养期长,草质柔软,家畜喜食,由于茎秆木质纤维少,在抽穗期和开花期叶量较少时,适口性良好[34],可用于刈割、放牧、加工草粉、调制干草或青贮等,是一种优良的野生牧草。因此,野大麦具有良好的生态价值和饲用价值,对改良环境和促进经济发展具有重要意义。

表2 羊草、野大麦和碱茅耐盐性比较Table 2 Comparison of the salt tolerance of Leymus chinensis, Hordeum brevisubulatum, and Puccinellia distans
Wilson 等[35]于20 世纪90年代初首次在野大麦体内发现Epichloë属内生真菌,WBE1、WBE3 和WBE4为野大麦种子中分离获得的代表性菌株,其分类地位为Epichloë bromicola[36]。内生真菌与野大麦共同进化,具有非寄生性,形成了互惠互利的共生关系[37]。禾草内生真菌一方面可以通过拮抗作用、诱导宿主防御或产生抗虫生物碱Peramine 和Loline,提高宿主对生物胁迫和非生物胁迫的耐受性,如抗虫性[38]、抗病性[39]、耐寒性[40]、耐盐碱性[41]、耐旱性[42]和耐重金属胁迫[43]等,降低了各种不利的环境条件、病原体和害虫对宿主植物的危害;另一方面内生真菌产生次级代谢产物,如麦角生物碱和吲哚二萜等,对家畜产生毒性[44-45]。尽管有研究显示野大麦–内生真菌共生体可以产生麦角类生物碱[46],但分离自甘肃临泽野大麦的WBE1、WBE3 和WBE4 菌株仅可以产生抗虫生物碱Peramine 和麦角类生物碱前体化合物Chanoclavine I,而不能合成对家畜有毒的麦角类生物碱[47],这充分展现出野大麦–内生真菌共生体的经济优势和生态优势,为其作为生态草奠定理论基础。
2 内生真菌对野大麦的影响
2.1 内生真菌对野大麦生长发育的影响
植株的发芽指标、形态指标、生理指标、光合指标以及营养元素指标等客观反映其生长发育状况,而内生真菌侵染的植物其生长发育受到一定影响[48-49]。野大麦的种子存在较强的休眠性,刚收获的种子发芽率极低,仅为6.3%,将其贮藏2~5年后发芽率较高,但是贮藏7年后急剧下降,贮藏10年后发芽率仅为20%[50]。浸种、去稃、红外线照射等方法可以打破休眠,提高野大麦的种子发芽率[51]。禾草内生真菌主要通过宿主种子垂直传播[52],在幼苗生长发育较适宜的环境条件下,内生真菌对种子萌发及幼苗生长的影响不显著(P> 0.05),但在逆境条件下如人工老化处理[53]、盐分胁迫[39]、酸碱胁迫[54]、水分胁迫和温度胁迫[42],其扮演着重要角色,表现为侵染内生真菌(E+)的野大麦种子的发芽率、发芽势、发芽指数、胚根长和胚芽长等发芽指标显著高于未侵染内生真菌(E–)的种子(P< 0.05)。杀菌剂甲基托布津处理E+野大麦时,抑制了发芽指标、形态指标和营养元素指标,而对生理抗性指标和光合指标没有显著影响[55]。
在三因素试验中,低浓度茉莉酸(JA)与内生真菌互作可以减轻盐胁迫对野大麦的危害,且E+的发芽指标显著高于E– (P< 0.05),0.1 μmol·L−1的JA 处理是盐胁迫下野大麦种子萌发的最适浓度[56]。而当种带真菌、内生真菌和盐胁迫三者互作时,尽管E+和E–间的表现与上述一致,但野大麦接种种带真菌后对种子发芽指标均有抑制作用,其中燕麦镰刀菌(Fusarium avenaceum)、根腐离蠕孢(Bipolaris sorokiniana)和细交链孢菌(Alternaria alternata)的抑制作用更强[57]。此外,三者的交互作用也影响着野大麦生长及生理变化(表3)。内生真菌还可以调控地上和地下部分的氮、磷含量,改善地下部分对钠的吸收[58],丛枝菌根真菌(arbuscular mycorrhizal fungi, AMF)与内生真菌互作时,也表现上述特性,尤其是在胁迫条件下表现更为显著[59]。
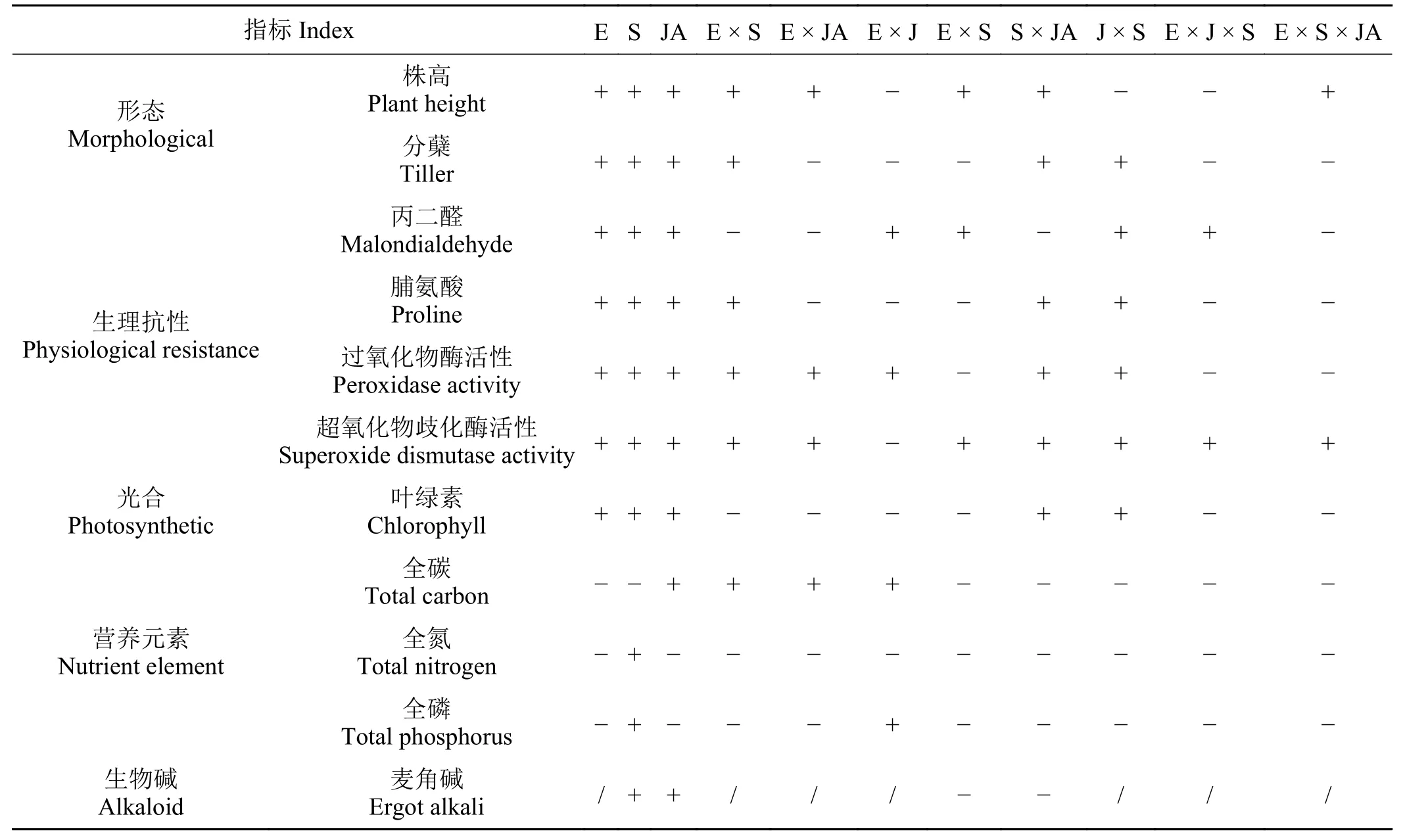
表3 内生真菌(E)、病原真菌(J)、茉莉酸(JA)和盐胁迫(S)互作对野大麦部分指标的影响Table 3 Effects of interactions between endophytic fungi (E), pathogenic fungi (J), jasmonic acid (JA), and salt stress (S)on the morphological indices, physiological resistance indices, photosynthetic indices,nutrient element indices, and alkaloid content of wild barley
内生真菌除影响野大麦生长外,其微生物群落和根际土壤的化学性质也与内生真菌有关[60]。这一系列的研究证明,内生真菌对提高野大麦生长的可抗逆性具有重要作用,内生真菌的侵染不仅促进野大麦种子萌发,而且与多种因素共同作用时,对生物量、株高和分蘖等都有积极作用,调节植物对营养元素的吸收和生理代谢活动,有利于野大麦在复杂的草地生态系统中形成稳定的群落,对改良土壤、改善生态环境做出重要贡献。
2.2 内生真菌对野大麦抗性的影响
前人的研究发现,内生真菌侵染禾草后,可以提高宿主抵抗不良环境的能力以及免受病虫的危害[61],可以将这一优良特性应用在野大麦的生态改良进程中。但当家畜不慎误食带有内生真菌的禾草,会导致中毒。研究人员早在1993年就有了一个明确的目标,即发挥禾草内生真菌的有益作用,但对采食家畜不产生毒性[62]。
2.2.1 抗病性
随着对生态环境的重视以及国民经济的快速发展,我国的城市绿化正在加速完善,栽培草地面积也在不断扩大,病害已经成为制约草地生产的主要限制因子,与其他科相比,在禾本科上发现的病害种类最多,高达1 289 种,其中有391 种为病原真菌[63]。Matthias 等[64]报道了Rph13基因座的分子遗传学特征,它是野大麦抗叶锈病的一种基因,随后在大麦品系PI 531 849 中被应用。
当野大麦内生真菌和病原菌互作时,发现不论是在温室还是离体叶片条件下,孢子浓度降低和病斑扩张速度减慢,以及各项生理指标均有所改善[65];在生态系统中有更强大的生长力和较高的存活率,例如,链格孢属(Alternariasp.)病原真菌[66]、球囊霉属(Glomusspp.) 菌根真菌[59]等。但也有研究表明,草地生态系统中病害的发生受多种因素的影响,导致内生真菌侵染的禾草对真菌病害的抗性表现出差异,可能内生真菌不能单独提高宿主的抗病性,而是通过提高其他抗性来间接提高抗病性[67]。
2.2.2 抗虫性
内生真菌可以产生抗虫生物碱,从而提高禾草的抗虫性[68]。野大麦在盐胁迫下提高了内生真菌的菌丝密度而间接提高了Peramine 的含量,增强了抗虫性[36]。Clement 等[38]也强调了内生真菌在赋予野生大麦抗虫性方面的潜在重要性,因此内生真菌为野大麦作为抗虫生态草的改良提供了有力的帮助。
2.2.3 耐盐碱性
我国西北内陆和滨海地区的盐渍土壤分布较多,不仅对西北地区的生态恢复与重建造成了障碍,而且严重制约当地经济发展。目前,土壤盐渍化导致的生态问题在世界范围引发广泛关注[69]。野大麦是泌盐盐生植物,在轻盐渍化的草甸上生长良好[13,30]。前人关于野大麦耐盐机制的研究有很多,通过克隆技术得到rbcS1、rbcS2[70]、HbCBL4、HbCBL10[71]、Bstl、Bst6、Bstl7、Bst22[72]等与盐胁迫诱导基因,为深入研究野大麦耐盐性分子机制奠定了基础。用转录组测序技术(RNA-Seq)探索野大麦的耐盐基因[73];利用RAPD-PCR 技术筛选野大麦耐盐新品系并进行突变品系鉴定和稳定性检验[74];在盐胁迫下探索野大麦的反应机制[75];利用组织培养和人工诱变技术,选育出“军需一号”耐盐碱的野大麦优良品种[76];筛选和鉴定野大麦的耐盐变异体[77]等,这些都为利用野大麦解决生态问题和改善土壤盐渍化奠定了基础。
内生真菌能够缓解盐过量引起的对植株的伤害,提高野大麦种子耐盐性,改善各项形态指标和生理抗性指标[78]。内生真菌侵染改变了野大麦多个代谢途径(如糖代谢、脂代谢、多胺代谢、氨基酸代谢、次生代谢物、信号转导等),导致其相关基因发生差异表达,例如内生真菌可以调控野大麦体内茉莉酸的合成,而茉莉酸信号通路与野大麦的耐盐机制有关[79];内生真菌的存在增强腐胺向亚精胺和精胺的转化以及从自由形态和可溶性共轭形态多胺向不溶性结合形态多胺的转移能力,降低了腐胺:(亚精胺 + 精胺)比值,这是内生真菌提高野大麦对盐胁迫抵抗能力的主要机理之一[41]。此外,转录因子(WRKY33、WRKY40、WRKY56、WRKY70) 调控着野大麦对盐胁迫的耐受性[79]。最新研究表明,在盐胁迫下,内生真菌通过改变野大麦的解剖结构,防止水分流失和运输能力下降,使植株保持较好的营养水平,从而抵抗盐胁迫的消极影响[80]。
2.2.4 其他非生物抗性
干旱胁迫影响草坪的持久性和草地生态系统的稳定性,进而影响栽培草地和生态环境建设。高渗透胁迫条件下,E+的野大麦种子萌发及幼苗的生长与E–相比具有显著优势(P< 0.05)[42]。除此之外,在其他逆境,如温度胁迫、水分胁迫、酸碱胁迫、人工老化以及多种因素互作条件下,内生真菌对野大麦的生长均起到一定的促进作用。
3 展望
生态草坪和草坪生态工程已成为我国草坪发展和草地生态系统改善的必由之路,野大麦作为潜在的生态草种,有诸多优良特性,尤其是与内生真菌形成的共生体具有多种抗性。但目前野大麦内生真菌共生体作为生态草广泛应用还面临许多困难。与羊草、紫花苜蓿等生态草相比,野大麦的品种较少且育成时间晚,目前仅有“军需一号”和“萨尔图”野大麦两个品种,又因其种子落粒性强,虽然对其自然繁殖有积极影响,但采种较为困难。
与黑麦草(Lolium perenne)、苇状羊茅(Festuca arundinacea)和醉马草(Achnatherum inebrians)内生真菌共生体体系的研究相比,野大麦 – 内生真菌共生体的研究较少,主要集中在生理生化方面的抗性研究和对畜牧业影响的研究,而缺乏在生态系统等方面探索,需要全面认识野大麦与内生真菌的互作机制,充分挖掘其在生态领域的巨大潜力。因此,为将野大麦 – 内生真菌共生体作为生态草在实际中广泛应用,应关注以下几点:
第一,加快野大麦育种的步伐。在华北、东北等野大麦广泛分布的地区挖掘内生真菌资源,筛选优良菌株,形成更加优良的野大麦–内生真菌共生体新品系;第二,优良的野大麦–内生真菌共生体进行种子扩繁,改善采种困难的现状,在盐渍化土壤上可大面积推广,发挥巨大的生态价值;第三,内生真菌与抗逆基因结合进一步提高野大麦的抗逆性。目前虽然野大麦抗逆基因的报道较少,但已经足够说明基因对提高抗性具有重要意义,内生真菌侵染对其抗性也有积极作用,而内生真菌与基因相结合对野大麦抗性的研究还尚未报道,应充分利用二者优势,挖掘内生真菌提高野大麦耐受性的基因,推进野大麦–内生真菌共生体的研究进程;第四,野大麦–内生真菌共生体可以结合农艺、工程以及最新研究技术,在具有不施用或少施用化学药剂、易养护管理、高经济效益等特点的初级生态草坪的基础上,建植高质量并兼具景观、生态、经济和社会效益的高级生态草坪;第五,内生真菌与禾草互作对生态系统的影响。前人研究发现昆虫食物网受禾草内生真菌的影响,内生真菌限制了植物向消费者的能量传递,食物网显示出在交替营养水平上昆虫的丰富度减少[81];因此有必要对野大麦–内生真菌共生体与生态系统间的相互作用进行探索,了解二者间的作用机制,为合理利用内生真菌提供理论基础。
综上所述,野大麦内生真菌共生体作为生态草有巨大优势,不仅在提高草地生态系统服务功能,保持草地生态系统的稳定和持续发展中发挥重要作用,而且与改善人类生存环境,实现人与自然和谐相处等方面具有重要意义。
猜你喜欢
厦门大学学报(哲学社会科学版)(2022年3期)2022-10-14
作文小学高年级(2022年5期)2022-06-16
环境卫生工程(2021年4期)2021-10-13
宝藏(2021年7期)2021-08-28
草地学报(2021年7期)2021-08-03
小学教学研究(2021年2期)2021-07-11
疯狂英语·新读写(2021年2期)2021-02-25
纺织服装周刊(2019年31期)2019-09-27
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
