
敲减ATF3对结直肠癌肿瘤干细胞自我更新与上皮间质转化及免疫抑制因子分泌的影响*
2021-12-21 08:21张勇夏悦明赖晓兰闵松林雷文俤
西部医学 2021年12期
张勇 夏悦明 赖晓兰 闵松林 雷文俤
(宁德市医院 1.普外二科;2.血液科;3.急诊科,福建 宁德 352100)
结直肠癌(colorectal cancer, CRC)是世界上第三大常见的胃肠道癌症类型,其发病率和死亡率均较高。近年来,每年约有180万新病例被诊断为CRC,占所有癌症发病率的约10%,且每年有80万的病例死于CRC[1-2]。各种环境因素和不良生活方式均会导致CRC的发生,例如吸烟、酒精摄入等。传统上CRC的疗法包括腹腔镜切除手术、放疗和化疗,但治疗后仍然会出现副作用,甚至癌症复发,因此迫切需要探究CRC新的治疗策略。结直肠癌肿瘤干细胞(colorectal cancer stem cells,CRC-CSCs)与其他肿瘤干细胞一样,具有自我更新和无限增殖的能力,可介导CRC肿瘤的生长、转移与复发。目前研究表明,CRC治疗后复发的主要原因可能是CRC-CSCs无法根除[3],因此,靶向抑制CRC-CSCs的恶性生物学行为可能是CRC治疗的重要策略。活化转录因子3(Activating transcription factor 3, ATF3)是具有碱性亮氨酸拉链结构的转录因子,属于ATF/CREB家族的成员,可在多种条件下调节基因的表达[4]。研究表明,ATF3在动脉粥样硬化、心肌肥大以及多种癌症发病机制中均发挥重要作用,且已发现ATF3与CRC恶性演进相关[5-7]。因此,本研究以分选CD133+表达的CRC-CSCs为对象,探究靶向敲减ATF3对CRC-CSCs自我更新、上皮间质转化等恶性生物学行为的影响。
1 材料与方法
1.1 材料 人结直肠癌细胞系HCT116购自中国上海科学院细胞库,胰蛋白酶、胎牛血清、牛血清白蛋白、双抗、McCoy’s 5a培养基以及DMEM/F12培养基购自Hyclone公司,TRIzol试剂、PrimeScriptTM1st Strand cDNA Synthesis Kit、SYBR®Premix Ex TaqTMII试剂盒购自日本Takara公司,CCK-8试剂盒和ELISA试剂盒购自凯基生物公司,免疫荧光染色试剂盒购自博士德生物公司,结晶紫染液、BCA蛋白质检测试剂盒、ECL化学发光液购自碧云天公司,抗体CD133、E-cadherin、Vimentin、Fas、FasL购自英国Abcam公司,辣根过氧化物酶标记的山羊抗兔购自昊鑫生物公司,sh-ATF3慢病毒载体由上海生工生物公司设计构建。
1.2 方法
1.2.1 HCT116细胞的培养 取HCT116细胞复苏,添加含10%胎牛血清与1%双抗(青霉素与链霉素)的McCoy’s 5a培养基,置于37℃、CO2恒温孵育箱进行培养,待细胞融合度达80%时,采用胰蛋白酶进行消化,并以1∶3的比例传代培养。
1.2.2 流式细胞术分选CRC-CSCs细胞 取对数生长期的HCT116细胞,加入含20%胎牛血清与 0.4%牛血清白蛋白的PBS封闭液,调整细胞浓度为2×106个/mL,置于4℃下作用 2 h,离心收集细胞,加入50 μL封闭液重悬细胞,然后分别加入5 μL CD133-PE,在4℃下避光孵育30 min,离心收集细胞,再使用1 mL预冷的PBS重悬细胞,加入磁珠4℃下避光孵育15 min,采用终浓度为2 μg/mL的嘌呤霉素室温处理10 min 以去除空白死细胞,按照说明书进行操作,通过流式细胞仪分选获得CD133+表达的CRC-CSCs细胞。在分选获得的CRC-CSCs细胞中加入含5 mg/mL胰岛素、0.4% BSA、2% B27、10 ng/mL FGF、10 ng/mL EGF 的无血清DMEM/F12 培养基中培养以进行后续实验。
1.2.3 慢病毒转染CRC-CSCs细胞 取生长状态良好的CRC-CSCs细胞,置于37℃、CO2恒温孵育箱中培养,待细胞融合度达80%时消化,调整细胞密度为4×105个/mL,取100 μL接种于24孔板进行转染。实验设置分组包括对照组、sh-NC组、sh-ATF3组,其中sh-ATF3组转染sh-ATF3慢病毒载体,sh-NC组转染阴性对照慢病毒载体,对照组细胞正常培养。以感染复数为20的比例加入慢病毒,转染培养12 h,收集两组细胞,培养于新鲜DMEM/F12培养液中,采用0.5 μg/mL嘌呤霉素进行筛选,每隔1天换液1次,持续2周。
1.2.4 实时荧光定量PCR检测敲减效率 收集转染后的3组CRC-CSCs细胞,加入TRIzol试剂提取细胞总RNA,通过紫外分光光度计检测提取的总RNA纯度与浓度。根据PrimeScriptTM1st Strand cDNA Synthesis Kit说明书进行操作,将总RNA进行反转录获得cDNA,以cDNA为模板,进行实时荧光定量PCR检测mRNA表达水平,步骤按照SYBR®Premix Ex TaqTMII试剂盒进行,以β-actin作为内参基因。扩增条件为:95℃ 5 min,循环1次;95℃ 30 s、60℃ 30 s、58℃ 30 s,此步骤一共循环42次。采用2-ΔΔCt法计算各基因的相对表达水平,利用Primer Premier 5软件设计基因引物,具体序列如下:ATF3上游引物5′-GACAAGGGAGACCTGGAGAA-3′,下游引物5′-GAGAAGGACAAGAAAGCCACA-3′;β-actin上游引物5′-ATGGTGGGAATGGGTCAGA AG-3′,下游引物5′-TCTCCATGTCGTCCCAGTTG。
1.2.5 CCK-8检测CRC-CSCs细胞活性 取对数期生长的转染后的3组CRC-CSCs细胞,调整细胞密度为4×105个/mL,取100 μL接种于96孔板中,置于37℃、CO2恒温孵育箱中培养12、24、48、72 h,每孔加入10 μL 的CCK-8试剂液,继续置于恒温孵育箱中孵育1 h,采用酶标仪检测450 nm处吸光度(A)值。
1.2.6 CRC-CSCs细胞成球能力检测 收集转染后的3组CRC-CSCs细胞,进行消化、离心,加入含生长因子的无血清培养基并吹打分散为单细胞悬浮液,调整细胞浓度以1×103个/孔的密度接种在96孔板,放置于37℃、CO2恒温孵育箱中培养,1周后于倒置显微镜下观察并计数每视野下形成的细胞球,以>50 μm作为一个细胞球。
1.2.7 平板克隆形成实验检测CRC-CSCs细胞克隆形成能力 收集转染后的3组CRC-CSCs细胞,离心并吹打分散为单细胞悬浮液,调整细胞浓度,以1×103个接种到无菌培养皿中,接种时轻晃培养皿使细胞分散均匀,置于37℃、CO2恒温孵育箱中培养,14 d后出现明显的细胞集落,PBS洗涤,加入4%多聚甲醛固定菌落,0.5%结晶紫染液染色15 min,流水冲洗干净,干燥后置于光镜下观察并计数细胞克隆形成数目,将≥50个细胞聚集作为1个细胞菌落。
1.2.8 免疫荧光染色检测E-cadherin、Vimentin表达 收集转染后的3组CRC-CSCs细胞,PBS洗涤后,加入 4%多聚甲醛室温固定10 min,自来水冲洗干净,滴加 0.5% TritonX-100 透膜处理15 min,加入10%山羊血清室温下封闭2 h后,接着加入E-cadherin(1∶100)、Vimentin(1∶100)一抗工作液,4℃下孵育过夜;次日,弃去一抗液,PBS洗涤后加入荧光二抗工作液(1∶500),暗室室温孵育1 h,洗去未结合二抗,滴加 DAPI 室温染色 10 min,PBS再次洗涤后封片,于荧光显微镜下观察并拍照。
1.2.9 Western Blot检测E-cadherin、Vimentin蛋白表达水平 取转染后的3组CRC-CSCs细胞,分别用PBS洗涤后,添加新鲜配制的RIPA裂解溶液,冰上静置裂解20 min后,离心收集各组上清液,BCA法测定蛋白质浓度。配制12%SDS-PAGE凝胶,分别取50 μg的各组蛋白样品上样,经过电泳分离蛋白质。并电转至PVDF膜(4℃,300 mA恒流1.5 h),接着置于5%脱脂奶粉中室温封闭2 h,TBST洗膜,将膜与E-cadherin(1∶1000)、Vimentin(1∶1000)、Fas(1∶1000)、FasL(1∶1000)一抗工作液共同置于4℃下孵育过夜;次日,洗膜后,加入辣根过氧化物酶标记的山羊抗兔(1∶5000)二抗工作液,37℃孵育1 h,TBST再次洗膜,滴加ECL化学发光液显色曝光,凝胶成像系统扫描图像后,Quantity One软件分析各条带的灰度值,并以GAPDH条带灰度值作为参照。
1.2.10 ELISA法检测细胞分泌的TGF-β1、VEGF、IL-6、IL-10水平 收集培养48 h后的3组CRC-CSCs细胞培养液上清,采用ELISA 法检测细胞培养液上清中TGF-β1、VEGF、IL-6以及IL-10的含量,实验步骤严格按照ELISA试剂盒说明书进行,采用酶标仪检测450 nm处各孔的A值。
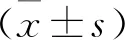
2 结果
2.1 CRC-CSCs细胞鉴定 通过流式细胞术分选CRC-CSCs细胞,CD133+CRC-CSCs细胞在分选前占23%,而分选后CD133+CRC-CSCs细胞达到了93%,见图1。

图1 流式细胞术分选CD133+CRC-CSCs细胞
2.2 转染后各组CRC-CSCs细胞中ATF3 mRNA表达比较 qRT-PCR检测结果显示,与对照组比较,sh-ATF3组CRC-CSCs细胞中ATF3 mRNA相对表达量显著下降(P<0.05),而sh-NC组与对照组的ATF3 mRNA相对表达量之间无统计学差异(P>0.05),见图2A。表明在CRC-CSCs细胞中敲减ATF3成功。
2.3 敲减ATF3对CRC-CSCs细胞活性的影响 与对照组比较,sh-ATF3组ATE3 mRNA表达水平降低(P<0.05),sh-NC组和对照组比较ATE3mRNA表达水平差异无统计学意义(P>0.05);CCK-8检测各组CRC-CSCs细胞活性结果显示,与对照组比较,sh-ATF3组CRC-CSCs细胞在48 h和72 h活性均显著降低,差异具有统计学意义(P<0.05);而在不同时刻,sh-NC组与对照组活性比较差异均无统计学意义(P>0.05),见图2B。

图2 各组CRC-CSCs细胞中ATF3 mRNA表达及活性比较
2.4 敲减ATF3对CRC-CSCs细胞成球能力的影响 成球实验结果显示,对照组、sh-NC组和sh-ATF3组CRC-CSCs细胞的成球个数分别为(87.56±5.89)、(84.70±5.06)、(20.12±1.58)。与对照组比较,sh-ATF3组细胞球体积明显缩小,成球个数也显著减少(P<0.01);sh-NC组和对照组比较细胞球体积无明显变化,成球个数之间差异无统计学意义(P>0.05),见图3。表明敲减ATF3能抑制结直肠癌肿瘤干细胞细胞球的形成。

图3 各组CRC-CSCs细胞成球能力检测
2.5 敲减ATF3对CRC-CSCs细胞克隆形成能力的影响 平板克隆形成实验结果显示,对照组、sh-NC组和sh-ATF3组CRC-CSCs细胞克隆形成数量分别为(87.56±5.89)、(84.70±5.06)、(20.12±1.58)个,sh-ATF3组CRC-CSCs细胞克隆形成数量较对照组减少,差异具有统计学意义(P<0.01);而sh-NC组细胞克隆形成数量与对照组比较,差异无统计学意义(P>0.05),见图4。

图4 平板克隆形成实验检测各组CRC-CSCs细胞克隆形成能力
2.6 敲减ATF3对CRC-CSCs细胞上皮间质转化过程的影响 免疫荧光染色结果显示,对照组和sh-ATF3组CRC-CSCs细胞中E-cadherin荧光染色较弱,E-cadherin蛋白表达少,而Vimentin荧光染色较强,蛋白表达相对较多;sh-ATF3组CRC-CSCs细胞中E-cadherin荧光染色明显增强,Vimentin荧光染色强度明显减弱,表明E-cadherin蛋白表达增加、Vimentin蛋白表达下降,见图5。Western Blot检测结果显示,与对照组比较,sh-ATF3组CRC-CSCs细胞中E-cadherin蛋白相对表达量较高,Vimentin蛋白相对表达量较低,差异具有统计学意义(P<0.05); sh-NC组与对照组比较,细胞中E-cadherin与Vimentin蛋白表达差异无统计学意义(P>0.05),见图6。
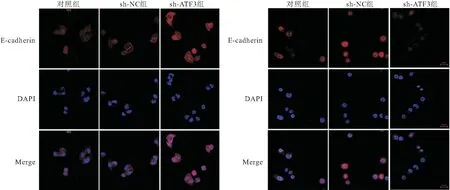
图5 免疫荧光染色检测各组CRC-CSCs细胞中E-cadherin、Vimentin表达

图6 Western Blot检测各组CRC-CSCs细胞中E-cadherin、Vimentin蛋白表达量
2.7 敲减ATF3对CRC-CSCs细胞分泌TGF-β1、VEGF、IL-6及IL-10的影响 ELISA实验检测结果显示,与对照组比较,sh-ATF3组CRC-CSCs细胞上清液中TGF-β1、VEGF、IL-10的含量均明显下降,差异具有统计学意义(P<0.01),而IL-6含量变化无统计学差异(P>0.05);sh-NC组与对照组比较,各组细胞上清液中TGF-β1、VEGF、IL-6、IL-10的含量之间的差异均无统计学意义(P>0.05),见表1。Western Blot检测结果显示,与对照组比较,sh-ATF3组CRC-CSCs细胞中Fas蛋白相对表达量较高,FasL蛋白相对表达量较低,差异具有统计学意义(P<0.05),sh-NC组和对照组比较,细胞中Fas和FasL蛋白相对表达量差异无统计学意义(P>0.05),见图7。

表1 各组CRC-CSCs细胞分泌TGF-β1、VEGF、IL-6、IL-10的含量比较

图7 Western Blot检测各组CRC-CSCs细胞中Fas、FasL蛋白表达量
3 讨论
CRC是全球常见的癌症类型之一,给患者和社会均带来了巨大负担。常规化学疗法常运用于CRC患者的临床治疗中,但是,除了具有潜在不良反应外,化疗药物还会诱导毒性产生限制化疗效果。CRC-CSCs细胞具有多向分化潜能和自我更新能力,致瘤性较高。此外,CRC-CSCs细胞在表型和分子结构上均与肿瘤内的其他细胞不同,其对化学药物和放射疗法都具有抵抗力,常规化疗药物只能消除肿瘤细胞,但对CSCs细胞的影响并不大,这在很大程度上导致了CRC患者治疗后的复发[3,8-9]。因此,确定新颖而特异的方法来靶向根除CRC-CSCs细胞,对于CRC患者的来说至关重要。
肿瘤是由多种具有异质性的细胞组成的复合物,可以轻松适应其环境并逃脱机体的免疫系统。肿瘤微环境在决定各种癌症的进展和反应中起着至关重要的作用,其可诱导非CSC向CSC的转化,此过程受许多因素的影响。而在多项研究中已将肿瘤微环境的改变与不同癌症干细胞表面标志物的表达联系起来[10]。CRC-CSCs细胞可以使用不同的细胞表面标记物进行识别,如CD133、CD24、CD44、CD29和ALDH1等[11]。其中,CD133是一种跨膜蛋白,被认为是许多类型肿瘤中癌症干细胞的标志物。研究表明,CD133+的CRC细胞对常规化疗具有抗性,且CD133高表达是CRC预后不良的一项重要标志[12-14]。目前,通过靶向肿瘤干细胞的表面标记物来消除CRC-CSCs细胞已成为CRC治疗的新靶点。此外,靶向CRC-CSCs细胞相关信号通路也是一种抑制CRC-CSCs细胞的方法,例如Valverde等[15]通过COX-2抑制剂靶向CRC-CSCs细胞中对于维持细胞更新能力起重要作用的WNT/β-catenin信号途径后发现,CRC-CSCs细胞生长受到了明显的抑制。Network等[16]研究表明了BMI1在CRC细胞中呈现过表达,能够诱导肿瘤发生转移,敲低BMI1可抑制癌症干细胞的自我更新能力。
ATF3是一种适应性基因,可对细胞外和细胞内的变化做出反应并调节细胞周期与凋亡,现已证明ATF3参与了癌症的易感性,其表达变化可引起相关信号途径发生改变,从而影响肿瘤的发生与发展[5,17]。据报道,在乳腺癌中ATF3能够促进癌细胞的增殖、迁移和侵袭,与癌症转移密切相关,因此可被用作乳腺癌死亡相关的独立预测因子[18]。本研究结果显示,在分选的CRC-CSCs细胞中敲减ATF3表达后,细胞活性下降,细胞的成球个数与菌落克隆形成数目也均下降,同时,细胞中E-cadherin蛋白表达较高而Vimentin蛋白表达较低,由此说明敲减ATF3可能抑制了CRC-CSCs细胞的恶性生物学行为。
肿瘤免疫逃逸与肿瘤的发生密切相关,通过分泌免疫抑制分子来抑制免疫功能,从而发挥免疫监视逃避的作用,这也是CRC发展的一项重要机制[19]。研究表明,CRC-CSCs细胞可能参与了免疫逃逸和免疫抑制,这有助于维持肿瘤相关的巨噬细胞以及调节性T细胞介导的免疫抑制的清除作用[20]。因此,靶向CRC-CSCs已被证明是克服免疫抑制作用的一种潜在方法。本研究结果显示,在敲减ATF3的CRC-CSCs细胞上清液中免疫抑制因子TGF-β1、VEGF、IL-10的含量均明显下降。TGF-β1作为肿瘤细胞分泌的优势免疫抑制分子,与肿瘤的浸润、转移以及分化等密切相关,VEGF参与调控肿瘤血管的生成与增生,IL-10在免疫反应和炎症中起多方向作用,由活化的T淋巴细胞、B淋巴细胞、巨噬细胞和单核细胞所产生[21-22]。本研究发现在敲减ATF3后可上调CRC-CSCs细胞中Fas蛋白表达,并且抑制FasL蛋白表达。而免疫保护作用由Fas/FasL介导,FasL诱导的T细胞凋亡是Fas介导的凋亡过程相关因子,其细胞外结构域可以与配体FasL结合,并且传递死亡信号至胞质区。当FasL与Fas结合后,可形成能够将凋亡信号传递至caspase 8的活性三聚体,从而激活一系列酶联反应,最终破坏肿瘤细胞[23]。
综上所述,本研究表明敲减ATF3可抑制结直肠癌肿瘤干细胞自我更新、上皮间质转化,下调相关免疫抑制因子的表达,其机制可能与调控Fas/FasL途径相关,这为CRC的诊疗提供一定的理论依据。尽管目前靶向肿瘤干细胞的研究发展趋势迅速,但仍然存在许多丞待解决的问题,了解相关分子信号传导途径维持并调节肿瘤干细胞的作用可能是改善癌症治疗的主要手段。此外,在了解肿瘤干细胞分子和基因特征后,进一步考虑研究以检测和靶向其他关键失调的途径,从而寻找肿瘤干细胞自我更新的新治疗靶标,可为阐明根除大多数恶性肿瘤细胞提供有效途径。
4 结论
敲减ATF3可抑制结直肠癌肿瘤干细胞自我更新与上皮间质转化,下调免疫抑制因子TGF-β1、VEGF、IL-10表达,并且上调Fas蛋白,下调FasL蛋白的表达。
猜你喜欢
环球时报(2022-09-20)2022-09-20
中老年保健(2022年1期)2022-08-17
保健医苑(2022年5期)2022-06-10
今日农业(2020年24期)2020-12-15
江西医药(2020年4期)2020-04-28
中华建设(2019年7期)2019-08-27
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
小资CHIC!ELEGANCE(2015年15期)2015-09-01
