细菌几丁质酶结构、功能及分子设计的研究进展*
2022-07-25 14:05颜子娟余俊红吴秀芸1王禄山
生物化学与生物物理进展 2022年7期
赵 沙 颜子娟 张 舒 余俊红 吴秀芸1,** 王禄山
(1)山东大学微生物技术国家重点实验室,青岛 266237;2)青岛啤酒股份有限公司啤酒生物发酵工程国家重点实验室,青岛 266000)
几丁质是自然界广泛存在的生物多糖之一,由N-乙酰-D-氨基葡萄糖(GlcNAc)经β-1,4-糖苷键连接聚合而成。几丁质作为结构多糖,存在于许多生物细胞外基质中,包括真菌细胞壁、昆虫外骨骼、甲壳类外壳和线虫卵壳等[1]。几丁质结构与纤维素类似,由单一单体聚合形成线型链状结构,相邻两个单体反向呈180°排列(图1a)。几丁质链通过分子内和分子间的氢键网络形成高度结晶的几丁质,构成抗降解屏障。根据几丁质链的方向差异可以分为α 型、β 型及γ 型几丁质[2]。不同于纤维素,几丁质的抗降解屏障不仅表现在高度结晶的整体结构[3],还表现在N-氨基乙酰化程度(degree of acetylation, DA) 和 糖 单 元 的 聚 合 程 度(degree of polymerization,DP)两个方面[2]。几丁质是重要的生物质资源,其降解产物GlcNAc和几丁质寡糖(chitin-oligosaccharide,CHOS)具有抗氧化、抗菌、抗肿瘤等多种特异性生物活性,被广泛应用于农业、食品工业和医药领域[4-6]。
利用几丁质生产高附加值产物主要有化学法和生物酶法两种方法。化学法需要使用高浓度的HCl或H2SO4,易造成危险和环境污染,生物酶法具有绿色、快速的特点,可以形成环境友好和生物相容的产物[7]。几丁质酶(EC 3.2.1.14)特异性水解几丁质糖苷键产生低聚寡糖。自然界中,几丁质酶广泛分布在古菌、细菌、真菌、高等植物及动物中[1],其中,细菌几丁质酶由于产生的丰度和商业潜能一直是研究的热点。细菌中几丁质降解酶基因存在明显的基因扩增及多结构域组合现象,并且不同家族、不同作用模式的几丁质酶系可以协同参与几丁质的高效降解[8],对生态系统碳氮比的平衡起着至关重要的作用[9]。因此,充分认识细菌几丁质酶的结构功能、降解模式及分子设计策略,对于高效生产高附加值产品,推动低品质生物质原料的绿色转化技术的快速发展具有重要意义。
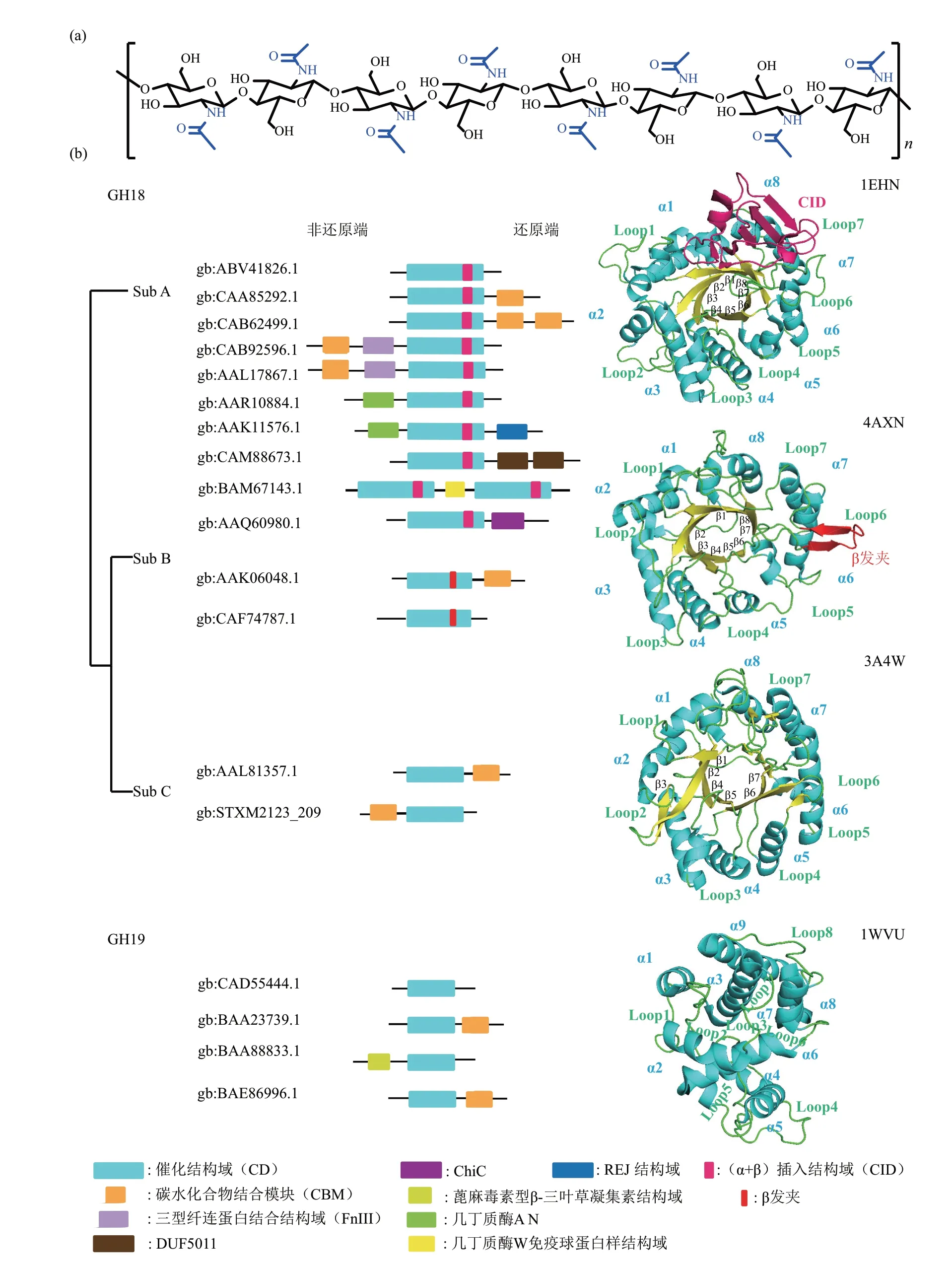
Fig.1 Structure of chitin and chitinase图1 几丁质与几丁质酶结构示意图
1 细菌几丁质酶的分类与结构特点
在碳水化合物活性酶数据库CAZy (http://www.cazy.org/)中,几丁质酶基于序列相似性主要被归类为GH18和GH19家族。其中,GH18家族几丁质酶广泛分布在古菌、细菌、真菌、高等植物及动物中,而GH19 家族几丁质酶主要分布于植物中,细菌中分布较少,且细菌中的GH19家族几丁质酶由植物几丁质酶进行基因水平转移产生[10]。截至2021年7月,GH18家族中被表征的细菌几丁质酶序列有192条,结晶结构有22个,而GH19家族被表征的细菌几丁质酶有15条序列,其中2个获得晶体并解析结构,分别为来源于天蓝链霉菌A3(2)(Streptomyces coelicolor A3(2))的Chi19G(PDB ⅠD 2CJL) 和 灰 色 链 霉 菌HUT 6037(Streptomyces griseus HUT 6037) 的ChiC (PDBⅠD 1WⅤU)。
几丁质酶一般由多个结构域组成,包括催化结构域(CD)、碳水化合物结合模块(CBM)和/或Ⅲ型纤维连接蛋白(FnⅢ)等底物辅助结合结构域(图1b)。GH18 家族几丁质酶催化结构域较为保守,为经典的(β/α)8TⅠM 桶结构,由8 个α 螺旋和8个β折叠经无规则卷曲(Loop)组成,催化裂隙位于桶结构的上方。GH18家族几丁质酶根据序列和结构特征又可被分为A、B、C 3 个亚家族。其中,Sub A 几丁质酶在Loop7 上具有(α+β)插入结构域(CⅠD),而Sub B和Sub C几丁质酶结构则不存在插入结构域,Sub B 在Loop6 上存在发夹结构[11]。GH19家族几丁质酶的催化结构域高度螺旋化,其底物结合裂隙相对较浅。有研究表明,GH18家族几丁质酶在催化裂隙中存在保守的催化序列DxDxE,其中,SubA 亚家族结构域CⅠD 中存在两个相对保守的序列YxR 与[E/D]xx[Ⅴ/Ⅰ],能够参与底物的结合和催化[12]。而GH19 家族几丁质酶催化中心具有一个相对保守的序列[FHY]-G-R-G-[AP]-x-Q-[ⅠL]-[ST]-[FHYW]-[HN]-[FY]-NY,并以两个Glu作为催化残基(质子酸与质子碱),二者结合可以作为GH19 家族几丁质酶识别的标志[10]。GH18 家族与GH19 家族保守结构的不同可能直接导致其作用机制的差异[13]。
2 细菌几丁质酶的作用机制
GH18 家族几丁质酶为底物辅助保留型催化断键 机 制 (substrate-assisted retaining catalytic mechanism),而GH19家族几丁质酶为单一置换反转型催化机制(single displacement inversion mechanism)。另外几丁质酶按照其作用模式分为内切几丁质酶和外切几丁质酶。前者可以随意切割聚合物的链内糖苷键生产寡糖,而后者只能从几丁质聚合物末端逐渐降解生成单体或二聚体[14]。GH18家族几丁质酶又可分为持续性几丁质酶和非持续性几丁质酶。持续性几丁质酶能够沿着底物糖链滑动,并在每次催化发生后不脱离几丁质链而进行持续水解[15],有效降解结晶几丁质。持续性几丁质酶多为外切几丁质酶,但也有例外,如人壳丙糖苷酶(human chitotriosidase,HCHT)是以持续性为降解模式的内切几丁质酶[16]。
2.1 酶与底物结合
不同几丁质酶底物结合裂隙的形状和组成不同(图2a)。GH18家族几丁质酶存在长且深的底物结合裂隙,至少包含5 个底物结合亚位点(Sub C 亚家族),Sub B 亚家族Loop 的差异使得活性亚位点增为7 个,Sub A 亚家族CⅠD 结构域的插入更加拓宽了酶分子与底物的结合裂隙,使亚位点扩展为8个。并且不同亚家族活性架构氨基酸的保守度明显不同,其中Sub A(PDB ⅠD 1EHN)亚家族的保守度最高,绝对保守氨基酸在-6 到+2 亚位点都有分布,Sub B(PDB ⅠD 4AXN)与Sub C(PDB ⅠD 3A4W)亚家族的保守度相对较高,在-3 至+2 亚位点均存在保守氨基酸。GH18 家族3 个亚家族都具有相同的催化断键位点(酸性的Asp和Glu)。而GH19家族几丁质酶(PDB ⅠD 3WH1)的底物结合裂隙较浅,一般可以容纳4个底物糖环,相对保守的氨基酸在-2 至+2 亚位点均有分布,但其保守性相对GH18家族几丁质酶较弱(图2a)。有趣的是,GH19 家族几丁质酶的正负亚位点结构对称,而GH18 家族几丁质酶正负亚位点结构具有不对称性,尤其是GH18 Sub A 亚家族。这表明在几丁质酶的进化过程中,通过增加结合底物的亚位点,并形成类似隧道的活性架构来获得持续催化的能力[17]。
GH18家族几丁质酶与底物C2官能团和突出的O6原子相互作用较多,在C2官能团方面,几丁质酶同时结合底物亚氨基氮原子和乙酰基氧原子,保守的GH18家族几丁质酶活性位点残基主要与底物的O7 原子相互作用;GH19 家族几丁质酶活性位点残基则不表现出对底物氮原子和氧原子的偏好。GH18 家族几丁质酶平均每个亚位点仅形成2 个氢键相互作用,GH19家族几丁质酶每个亚位点平均可形成4 个氢键相互作用,GH18 家族几丁质酶相对较少的氢键相互作用可加速持续性。GH18 和GH19家族几丁质酶活性中心表面都包含多个保守的芳香族氨基酸残基,这些残基通过疏水相互作用力结合几丁质底物[18-19],但GH18家族几丁质酶的CH-π 相互作用平均分布于8 个亚位点,并且具有相对较高的保守性,为持续性作用提供滑行的轨道[7],而GH19 家族几丁质酶的CH-π 相互作用主要存在于-2 及+2 亚位点,且保守性相对较低,可稳定底物结构(图2b)。综上所述,GH18 家族持续性几丁质酶活性架构亚位点的不对称性以及活性中心芳香族氨基酸的高度保守性可能是该家族几丁质酶具有持续性催化机制的结构基础[20]。

Fig.2 Conserved analysis of chitinase active center图2 几丁质酶活性中心保守性分析
2.2 催化机制
目前,随着对GH18家族几丁质酶催化机制的研究,越来越多的证据表明底物辅助保留模型能够更好地解释反应机理。以来自黏质沙雷菌(Serratia marcescens)的几丁质酶SmChiB(PDBⅠD 1E6N)为例阐述[21],当GlcNAc进入-1亚位点时,在氨基酸残基的作用下发生构象变化,由椅式构象变为船式构象,使得N-乙酰基的羧基氧原子靠近异头碳(C1)并发起亲核攻击,Tyr214 与羧基氧原子形成氢键稳定该结构。Asp142 进行翻转与N-乙酰基形成氢键,稳定中间体结构,另外Asp142 翻转后靠近Glu144 并与其形成氢键激活质子供体,糖苷键断裂,产物解离,恶唑啉离子中间体形成。后催化水分子对C1 发动亲核攻击,恶唑啉离子中间体解体,Glu144 恢复初始状态,Asp142 在Asp140 的辅助下翻转回初始位置,产物释放,水解完成(图3a)。但也有个例存在,如来自同一株菌的SmChiA(PDB ⅠD 1EHN)[22]的催化过程中并未形成恶唑啉离子中间体。在底物糖苷键质子化后,与Asp311相互作用的Asp313翻转到其替代位,并与催化残基Glu315 相互作用使得-1 亚位点GlcNAc的N-乙酰氨基团绕C2-N2旋转,N-乙酰氨基团的N-H翻转至另一位置完成水解过程。
GH19 家族几丁质酶采用单一置换反转机制,以来自蕊形真藓(Bryum coronatum)的GH19家族几丁质酶BcChi-A(PDB ⅠD 3WH1)[23]为例进行阐述。催化过程如下(图3b):催化反应发生前,Glu61作为催化酸,Glu89作为催化碱。GlcNAc到达-1亚位点时保持椅式构象不变。Glu61提供H离子使糖苷键中的异头氧原子质子化,糖苷键断裂,形成羰基碳鎓离子中间体,同时Gln164 与-1 亚位点的GlcNAc的N-乙酰氨基团形成氢键,从而阻止恶唑啉离子中间体的形成。Glu70作为一般碱起着稳定水分子的作用,当中间体形成后,被Ser102的羟基氧原子形成氢键固定的催化水分子可以被Glu70激活并发生亲核攻击。水解产物发生构型反转,产生α异头物,产物释放,水解完成。
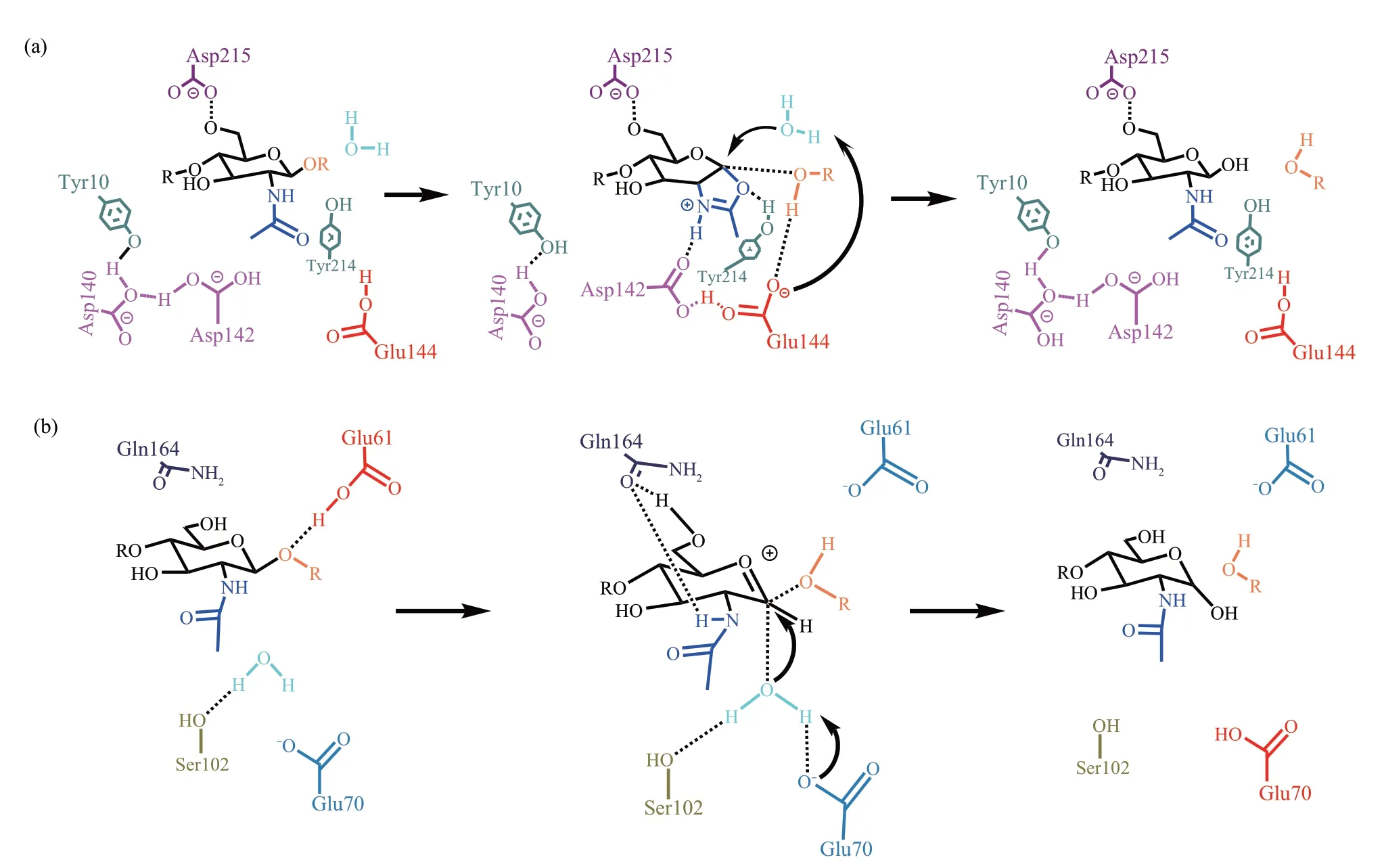
Fig.3 The schematic diagram of chitinase catalysis mechanism图3 几丁质酶催化机制示意图
3 持续性作用机制
由于结晶几丁质具有抗降解性,持续性几丁质酶要从结晶表面剥离出单条几丁质链,进而可以持续性降解,不断产生二糖单元。目前,持续性几丁质酶的结构基础在模式细菌S.marcescens中得到一定研究。S.marcescens包含两个持续性几丁质酶,SmChiA 和SmChiB,属于GH18 家族并具有保守的CⅠD结构域[12]。其中SmChiA从非还原端向还原端移动,在非还原端在释放(GlcNAc)2,而SmChiB则从还原端向非还原端移动,在还原端释放(GlcNAc)2(图4a),SmChiA的半衰期和速度都比SmChiB 大,表明不对称的亚位点结构决定了结晶多糖的降解方向和程度[24]。结晶结构表明两者的活性架构中芳香氨基酸呈线性排列[18]。这些芳香族残基(特别是Trp)可以与糖环的一侧或两侧形成堆积或疏水相互作用,并形成一个灵活的鞘,推动几丁质链沿着鞘滑动[25]。活性架构芳香族氨基酸的缺失通常会导致酶对结晶几丁质底物的亲和力、持续性和活性降低[7]。此外,活性架构中极性残基与芳香残基均呈线性排列(图4b),也可能参与持续性。Hamre 等[25]证明活性架构极性氨基酸Thr276 在持续性作用过程中发挥重要作用,突变体ChiA-T276A产物的[(GlcNAc)2]/[GlcNAc]比例降低近50%,几丁质降解效率降低26.7%。Jana等[26]通过定点诱变确定了S.marcescens几丁质酶SmChiB 活性架构中极性氨基酸的作用。但是,目前对极性残基在底物结合、糖苷酶作用模式和持续性中的作用的了解仍然相对有限。

Fig.4 The schematic diagram of the processive catalysis mechanism of chitinases图4 几丁质酶持续性催化机制示意图
尽管结晶几丁质和纤维素的物理和化学稳定性是相似的,但是几丁质酶移动产生双糖的速度是纤维素酶的10 倍[27-28]。几丁质酶快速单向运动的机制越来越受到人们的关注。Nakamura等[29]利用高精度单分子成像、X射线晶体学和全原子动力学模拟等技术研究了SmChiA单向持续性运动机理,分为化学步骤(底物辅助催化和产物释放)和机械步骤(脱晶和链滑动)。在另一项研究中,Nakamura等[30]通过单分子荧光成像成功测定了SmChiA 的基本速率常数,结果表明持续性几丁质酶SmChiA需要与内切几丁质酶以及辅助催化酶类AA10家族裂解多糖单加氧酶CBP21(LPMO)协同作用,才能有效地结合到链端并增加水解活性。为了研究几丁质酶的持续性,人们开发了各种各样的研究方法。经典的方法包括使用薄层色谱法(thin layer chromatography,TLC)、高效液相色谱法(high performance liquid chromatography,HPLC)等方法定量产物,计算G2/(G1+G3)的比例[31-32]以及可溶性还原糖的比例[33],使用水溶性壳聚糖(部分去乙酰化几丁质)作为底物测定产物中偶数糖与奇数糖的比例。此外,使用荧光标记底物[34-35]或安培生物传感器[36]已成功地确定有效结合酶的分数,以估计酶的持续性能力。近年来,高速原子力显微镜[37]或全内反射荧光显微镜[30,38]等单分子成像技术的应用,使人们能够直接观察有效结合的持续性酶。
4 细菌几丁质酶系协同降解模式
自然界中,几丁质经过脱乙酰化等修饰作用,其结构具有一定的复杂性,需要多种几丁质酶协同降解[2,39]。一些细菌可以分泌多种几丁质酶,利用特异的协同降解模式实现高效降解几丁质。代表菌株是模式菌S.marcescens,分泌GH18 家族几丁质酶SmChiA、SmChiB 和SmChiC、GH20 家族壳二糖酶(chitobiase)和LPMO[27,40-41];嗜热链霉菌Streptomycessp. F-3,胞外分泌6 种几丁质降解相关蛋白,除了GH18 家族几丁质酶SsChi18A、SsChi18B 和SsChi18C、GH20 家 族SsGH20A 和AA10 家族裂解多糖单加氧酶SsLPMO10A 外,还包含一种GH19 家族几丁质酶SsChi19A[11];红茶纤维弧菌(Cellνibrio japonicas)可分泌GH18家族的几丁质酶CjChi18B、CjChi18C、CjChi18D 及GH18 家族糖苷酶CjChi18A,及AA10 家族的CjLPMO[41]。几丁质降解酶系包含至少4 类酶:a. LPMO,如CBP21、SsLPMO10A 和CjLPMO,通过氧化裂解作用随机切割几丁质聚合物释放自由链末端;b. 内切几丁质酶,如GH18 家族的SmChiC、SsChi18B、SsChi18C、CjChi18C 和CjChi18D,GH19家族的SsChi19A,通过水解作用使糖苷键断裂,从而释放自由链末端或产生寡聚糖;c. 持续性外切几丁质酶,如SmChiA、SmChiB、SsChi18A 及CjChi18B,它们均属于GH18-Sub A 家族,可从几丁质链的还原端或非还原端持续性降解并产生二糖单元;d.壳二糖酶或N-乙 酰 氨 基 葡 萄 糖 苷 酶 (endo-β-Nacetylglucosaminidase,NAG),如GH20 家族的壳二糖酶、SsGH20A 和GH18 家族的CjChi18A,将(GlcNAc)2水解为GlcNAc。几丁质酶系协同降解过程如下[42]:LPMO 和内切几丁质酶在结晶几丁质表面作用释放自由链末端,持续性几丁质酶由链末端切入、剥离几丁质单链并进行持续性降解产生二糖单元,内切几丁质酶在作用过程中产生的寡聚糖,由内切几丁质酶或持续性几丁质酶进一步降解产生寡糖,最后壳二糖酶或β-N-乙酰氨基葡萄糖苷酶将寡糖降解为单糖,运送至胞内进一步降解(图5a-c)。研究表明,一些不具备几丁质水解活性的结合蛋白(chitin binding protein,CBP)可增加底物可及性与几丁质酶协同作用[43]。
在细菌中,磷酸烯醇丙酮酸依赖的磷酸转移酶系统(PTS)和ATP结合盒(ABC)转运体分别负责GlcNAc单体和低聚物的摄取[44],这些产物在胞内进行代谢,经EMP途径进入TCA循环,实现几丁质的彻底降解(图5d)。此外,弧菌几丁质水解酶的诱导受到组氨酸激酶和双组分系统的复杂调节(图5e)。在静息状态下,CBP 与膜蛋白ChiS 的周质域结合。当分泌的几丁质酶将几丁质降解为胞外空间的低聚糖时,低聚糖被孔蛋白运输并与CBP结合。在结合状态下,CBP/ChiS 复合物解离并转运信号表达几丁质降解基因。ChiS 的胞质结构域由3 个亚结构域组成:依赖ATP 的组氨酸激酶/磷酸酶(HK)结构域、Asp 响应调节器(RR)域和组氨酸磷酸化转移(HP)结构域[45]。
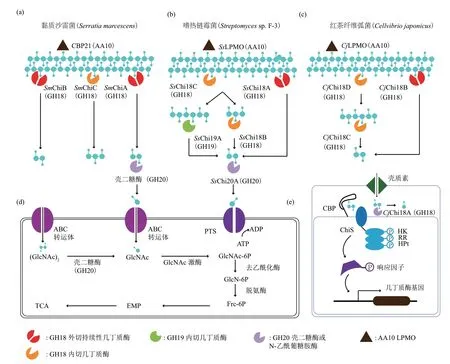
Fig.5 The shematic diagram of synergistic degradation of chitinases图5 几丁质酶协同降解示意图
5 细菌几丁质酶系的理性设计策略
为满足工业应用需求,提高几丁质酶的催化活性和热稳定性是目前几丁质酶设计的研究热点,表1总结了近年来对几丁质酶设计改造的实例。传统的设计策略定向进化技术通过随机诱变或重组技术获得突变体,不需对酶的结构和功能有深入的了解,但是构建的突变文库容量非常大,所以该方法对高通量筛选的方法要求非常高[46]。Songsiriritthigul 等[47]利用定向进化技术,对地衣芽孢杆菌(Bacillus licheniformis)来源的GH18 家族ChiA进行易错突变,从10个突变体中最终筛选到酶活性提高2.7 倍的几丁质酶突变体。Menghiu等[48]通过易错PCR 技术建立Bacillus licheniformisDSM8785 chiA 的突变体库,使用荧光激活细胞分选(FACS)技术筛选出酶活提高100%的突变体DH08,发现在DH08 中3 个位点发生突变分别是K128E、H130N和D220Y。
目前对几丁质酶进行设计使用最多的是半理性设计策略,该策略在对结构简单认识的基础上对特定结构域的残基进行突变,建立简洁的突变体库,通过组合筛选出最优突变体。例如,采用基于共识的半理性设计引入二硫键和Pro 提高了几丁质酶PpChi1 的热稳定性,50℃时半衰期值为野生型的26.3 倍,最适反应温度由45℃提高到52.5℃[49]。用来自环状芽孢杆菌WL-12 (Bacillus circulansWL-12)BcChiA1 的CBM12 代 替 白 长 链 霉 菌ATCC 27414 (Streptomyces albolongusATCC 27414)SaChiA4 的CBM5,使SaChiA4 对几丁质粉的活性提高了近54%(28×103U/g),对胶质几丁质活性提高了49%[50]。深绿木霉PTCC5220(Trichoderma atroνiridePTCC5220)的Chit42 缺乏1 个几丁质结合结构域,将S. marcescens SmChiB的CBM 与Chit42 融合,得到了一种具有更强几丁质结合能力的嵌合几丁质酶,并且嵌合几丁质酶对植物病原真菌具有较高的抗真菌活性[51]。
理性设计B.circulans的ChiA1 突变体突变T682A 导致了对几丁质底物更高的特异性[52]。突变S.marcescensChiB 的表面残基构成G188A/A234P 双突变体,其在57℃时,半衰期增加了10倍,表观Tm增加了4.2℃[53]。S.marcescensB4A 的几丁质酶突变体S390Ⅰ可耐受90℃高温[54]。苏云金芽孢杆菌WB7(Bacillus thuringiensisWB7)活性位点突变体E209Q拓宽pH[55]。
随着几丁质酶被表征的序列和结构不断增多,计算分析方法不断更新。可以基于蛋白质(亚)家族丰富的序列数据的祖先序列重构技术来对几丁质酶进行设计。研究表明,寒武纪前的酶分子比现存的酶更耐高温,所以将现存酶恢复成祖先酶的序列可以设计出耐热蛋白,该方法现已成功获得了更耐热的细胞色素P450 酶和更耐高温的酮醇酸还原异构酶[56]。另一方面,可以基于结构生物信息学进行理性设计,利用已有结构进行分子模建,构建系统发育树及酶活性中心序列谱。AlphaFold 利用多序列比对,将有关蛋白质结构的物理和生物学知识结合在深度学习算法的设计中,高度精确地预测蛋白质结构,为理性设计提供精确的模板[57-59]。通过对GH18 3 个亚家族和GH19 家族的活性中心序列谱分析发现,其活性架构亚位点数是从大到小变化的,并且保守性也是越来越低的。这种基于结构生物信息学的统计分析可以定位关键功能区和功能残基,建立小而精的突变文库,从而有效改变酶的稳定性、活性及底物特异性等[60]。
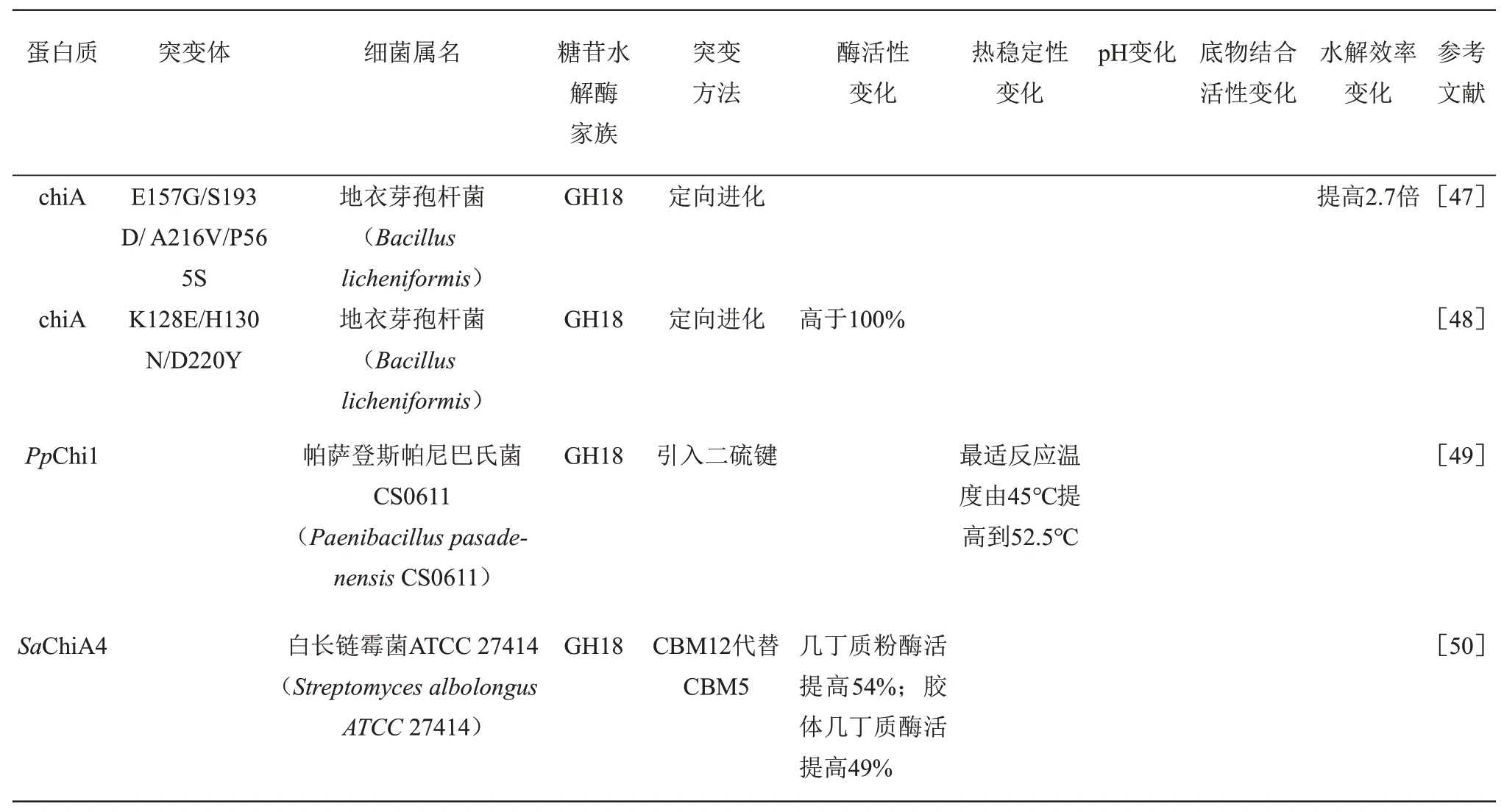
Table 1 Recent progress in engineering activity and stability of chitinases表1 几丁质酶改造实例
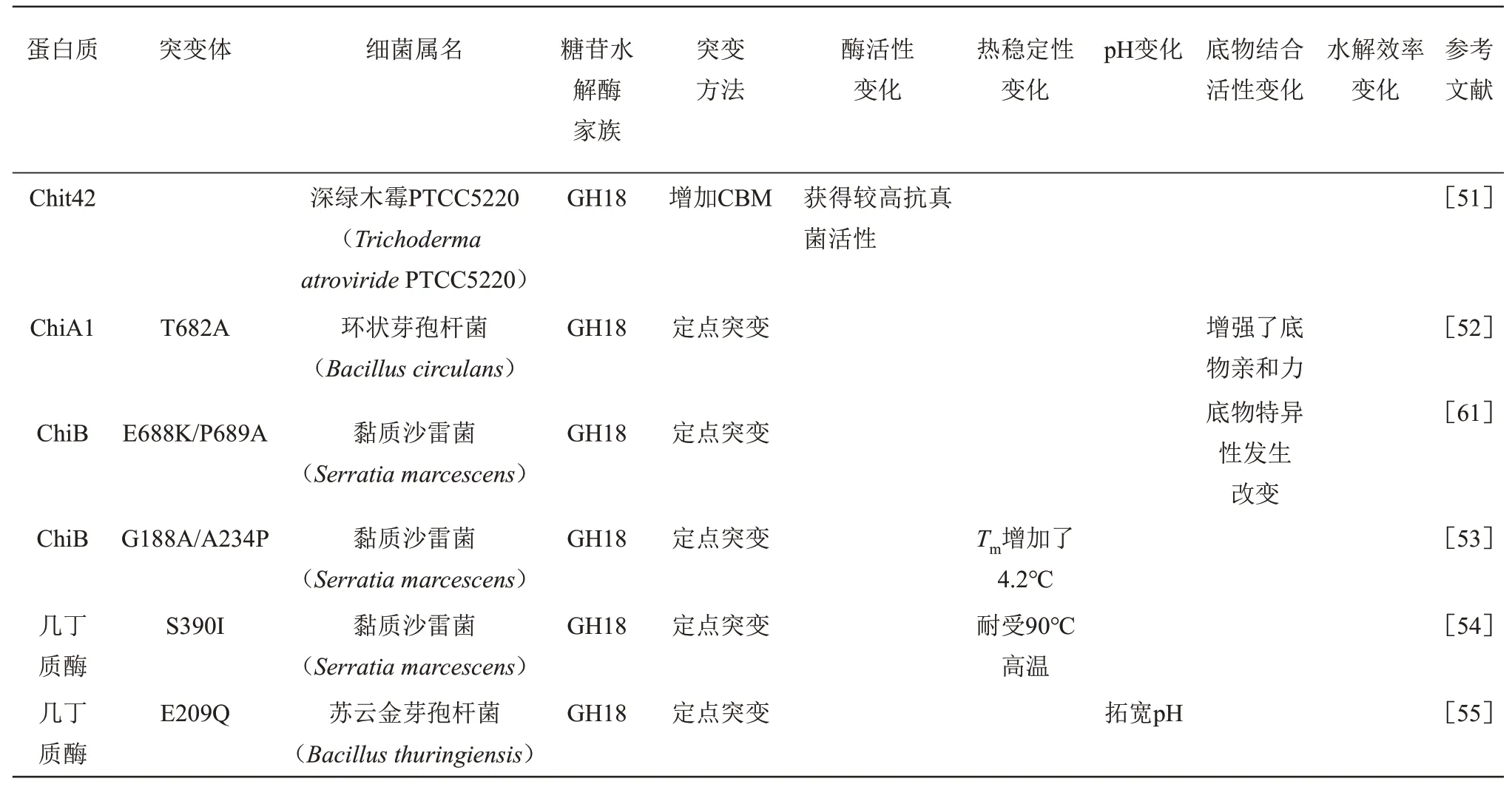
续表1
6 总结与展望
几丁质酶解产物CHOS 及GlcNAc 在农业、医疗、工业等方面发挥重要作用。CHOS被认为是潜在的益生元,容易被人体肠道吸收并可抑制一些食物致病菌的生长[62]。研究发现,CHOS 不仅能够影响各种复杂途径抑制结肠黏膜内的炎症[63],而且在新的抗癌治疗策略中得到了广泛的关注[64]。GlcNAc 能够促进成纤维细胞释放酸性黏多糖,恢复胃肠道保护结构的形成,是一种潜在的治疗炎症性肠病的候选药物[60]。在未来的研究中,可以运用代谢工程策略将代谢通量转向目标产物,也可以有效强化整个几丁质降解途径,促进目标产物合成。
虽然许多细菌几丁质酶被纯化和表征,但不同几丁质酶确切的相互作用和催化机制仍然是不清楚的,持续性几丁质酶前进的动力之源仍未得到解答,LPMO 氧化还原作用所需还原能的来源仍未知,进一步的机制研究是必须且迫切的。另外,GH19 家族几丁质酶的结构与功能有待进一步研究。几丁质协同降解酶系的精细分工与合作仍需进一步探究。目前,蛋白质工程设计改造现存的几丁质酶分子,以提高几丁质酶的热稳定性和催化活性,构建高效的协同降解酶系是工业亟待解决的关键问题。虽然人们已经尝试运用定向进化、半理性设计等方法来设计酶分子,但是仍需要建立一个普适的策略能够大幅度地提高酶分子的性质。前面已经提到了祖先序列重构和基于结构生物信息学的理性设计策略需要被分析和发展,依赖于结构和大量的时间、资源来训练数据集的分子动力学模拟和深度学习技术在用来改造酶分子的道路上仍任重道远。
猜你喜欢
蔬菜(2022年3期)2022-11-11
中国农业大学学报(2022年3期)2022-05-19
教育周报·教育论坛(2020年3期)2020-10-21
热带作物学报(2020年7期)2020-09-02
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
科技资讯(2017年12期)2017-06-09
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26