
覆盖雷竹笋箨叶碳氮代谢特征及其与竹笋食味品质的关系*
2022-12-23 09:04董亚文陈双林郭子武杨丽婷谢燕燕
林业科学 2022年8期
徐 森 董亚文 陈双林 郭子武 杨丽婷 谢燕燕
(中国林业科学研究院亚热带林业研究所 杭州 311400)
竹笋食味品质是体现竹笋经济价值的重要因素,不同竹种以及同一竹种不同区域、环境条件和栽培措施下竹笋品质差异明显(郭子武等, 2019)。研究发现,土壤氯质量浓度对雷竹(Phyllostachysviolascens‘Prevernalis’)笋呈味物质和粗糙度物质含量有重要影响,高氯施肥下竹笋甜味物质含量及甜味和鲜味氨基酸比例均明显提高,竹笋食味品质明显改善(杨丽婷等, 2020)。而光照与酚类和纤维类物质的合成密切相关,遮光处理下毛竹(Phyllostachysedulis)笋单宁和纤维类物质含量显著降低,而竹笋的甜味明显升高(艾文胜等, 2012)。也有研究表明,3月气温与厚竹(Phyllostachysedulis‘Pachyloen’)笋单宁含量呈明显正相关,与粗纤维含量呈明显负相关,而年降雨量影响厚竹根系吸收和竹笋生长速率,与竹笋碳水化合物等的合成密切相关(李苑等, 2018)。可见,温度、养分和土壤等环境因素对竹笋食味品质有显著影响,探索异质环境下竹笋食味品质形成机制,对于竹笋品质改良技术的研发及应用有重要参考。
雷竹隶属竹亚科(Bambusoideae)刚竹属,是我国优良的笋材兼用竹种。20世纪90年代以来,依据雷竹生物学和生态学特性,浙江省杭州市临安区和余杭区等竹产区推广应用雷竹林地覆盖竹笋早出技术,林地覆盖后笋期明显提前,竹林经济效益显著提高(郭子武等, 2013c)。覆盖栽培不仅会引起雷竹林土壤养分、水分和土壤孔隙度等土壤理化性质的明显变化,同时雷竹鞭系生长区域的温度和光照等环境条件亦会产生剧烈变化(Lietal., 2017; 郭子武等, 2013b),这种变化对立竹叶片养分特征(陈珊等, 2014a)、鞭根养分和抗性特征(时俊帅等, 2019a; 陈珊等, 2015)等的影响研究已有大量报道,但对箨叶碳氮代谢特征的影响及其与竹笋食味品质关系的研究未见报道。箨叶是竹子分类的重要器官,着生于箨鞘顶端,是竹笋出土时最先感光器官(徐森等, 2022),许多竹种竹笋在土中时,苦涩味轻、甜味强,但出土后,食味品质就发生明显变化,苦涩味明显增强,甜味降低,这表明箨叶与竹笋食味品质有明显的相关关系。鉴于此,本试验开展了自然经营雷竹林和覆盖雷竹林箨叶养分含量、碳氮代谢酶活性和竹笋食味品质的差异探究,分析雷竹林地覆盖对竹笋箨叶碳氮代谢特征的影响,以及与竹笋食味品质存在密切的关系,进而探讨箨叶对竹笋食味品质的作用,为高品质竹笋培育提供参考。
1 研究区概况
试验地位于浙江省杭州市临安区太湖源镇(119.62°E,30.33°N),该地为亚热带季风气候,年平均气温15.4℃,极端高温和低温分别为40.2℃和-13.3℃,1月和7月平均气温分别为3.2℃和29.9℃,年降水量1 250~1 600mm,年日照时数1 850~1 950 h,全年平均无霜期235天,土壤为红壤。该镇雷竹资源丰富,现有竹林面积1万多hm2。
2 材料与方法
2.1 样地设置和样品采集
2020年1月和3月,在试验区选取立地条件一致的覆盖(LF)和未覆盖(LZ)雷竹试验林各3块,每块试验林面积不小于0.4 hm2,其中覆盖雷竹林具体措施为: 11月中下旬对林地施肥和浇水,后采用有机材料双层覆盖法进行地表覆盖,即下层覆盖10cm左右稻草作为增温材料,上层覆盖20~30cm左右砻糠或竹叶作为保温材料,至翌年3月自然出笋时清除覆盖物。在各覆盖雷竹试验林中挖取刚露出覆盖物的竹笋20株,共计60株覆盖笋; 未覆盖雷竹试验林中挖取出土约15cm的竹笋20株,共计60株未覆盖笋。鲜笋标记编号后放在冰盒中立即带回实验室。
2.2 试验方法
2.2.1 箨叶生理指标测定 清洗竹笋样品,并擦干表面残留水分,每株样笋自下而上依次选取6~8片箨叶,混合均匀后取部分鲜样剪碎装瓶于-80℃低温冰箱内保存,供测定箨叶碳氮主要代谢酶活性,另一部分鲜样于烘箱中105℃杀青30min,再85℃烘至恒质量,研磨成粉末用于箨叶养分含量的测定。箨叶蔗糖合成酶、蔗糖磷酸合成酶、中性转化酶和酸性转化酶活性均采用高俊凤(2006)的方法测定,淀粉酶活性采用李合生(2007)的方法测定,谷氨酸合酶活性采用熊丹(2012)的方法测定,硝酸还原酶和谷氨酰胺合成酶活性均采用苏州科铭生物技术有限公司生产的试剂盒测定,C、N含量分别采用重铬酸钾容量法和凯氏定氮法(鲁如坤, 2000)测定,可溶性糖和淀粉含量采用蒽酮比色法(周永斌等, 2009)测定。
2.2.2 竹笋食味品质测定 剥去样笋笋壳,切取可食用的鲜嫩部分用粉碎机粉碎,取部分匀浆用于竹笋呈味物质的测定; 另一部分匀浆于烘箱中60℃烘至恒质量后用于纤维类物质的测定。可溶性糖的测定参照NY/T 1278—2007 铜还原碘量法(中华人民共和国农业部, 2007),总酸的测定参照GB/T 12456—2008滴定法(中华人民共和国卫生部, 2008),单宁的测定参照NY/T 1600—2008分光光度计法(中华人民共和国农业部, 2008),草酸的测定参照俞乐等(2002)反相高效液相色谱法,游离氨基酸的测定参照莫润宏等(2012)日立 L-8900氨基酸分析仪方法,纤维素和木质素的测定参照王玉万等(1987)硫酸水解法。
2.3 数据处理
采用SPSS 23.0统计软件进行独立样本t检验,分析(α=0.05)雷竹覆盖笋和自然笋箨叶养分含量、碳氮代谢酶活性及竹笋食味品质间的差异,采用Canoco5.0软件进行冗余分析(Redundancy analysis,RDA)对雷竹笋箨叶碳氮代谢指标和竹笋呈味物质、粗糙度物质的关系进行分析,采用Origin 2018进行图表制作。
3 结果与分析
3.1 雷竹箨叶养分含量的覆盖效应
由图1可知,覆盖后箨叶可溶性糖、淀粉和NSC含量均显著升高,升幅分别为22.88%、34.66%和32.55%,N含量显著降低,降幅为17.65%,而C含量无明显变化。以上结果表明,覆盖对箨叶主要养分特征有明显的影响,这可能与覆盖条件下雷竹林土壤湿度、温度和光照等环境条件的剧烈变化,从而明显影响竹林光合产物的代谢利用和氮素的吸收利用等有关。
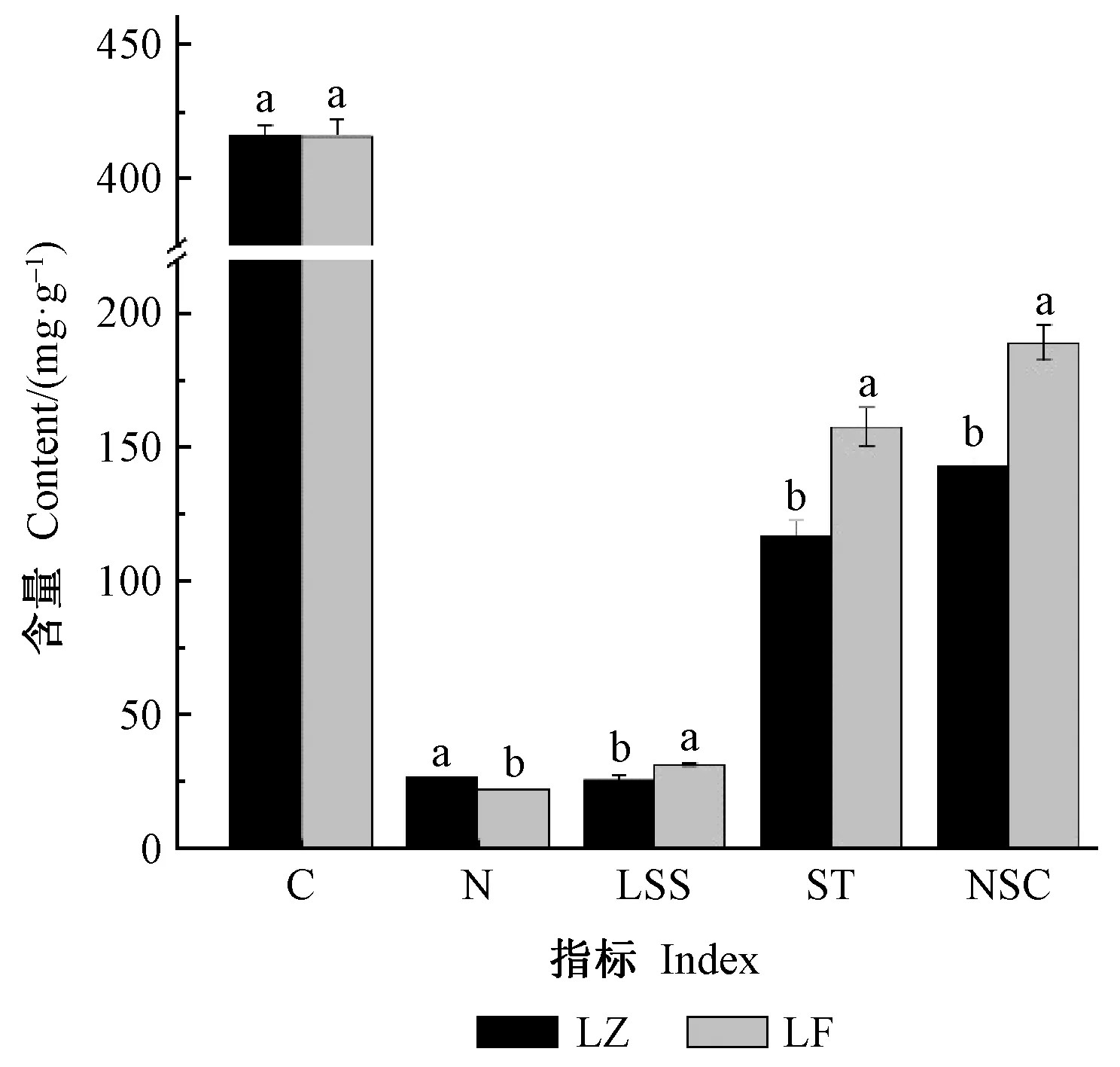
图1 雷竹箨叶养分含量
3.2 雷竹箨叶碳氮主要代谢酶活性的覆盖效应
由表1可知,覆盖后雷竹箨叶蔗糖合成酶、谷氨酸合酶、硝酸还原酶和谷氨酰胺合成酶活性均显著升高,其中覆盖笋箨叶蔗糖合成酶、谷氨酸合酶、硝酸还原酶和谷氨酰胺合成酶活性分别是自然笋的1.24、3.06、1.34和4.30倍,而箨叶蔗糖磷酸合成酶、酸性蔗糖转化酶、中性蔗糖转化酶和淀粉酶活性均无显著差异。以上结果表明,覆盖对雷竹笋箨叶碳代谢酶活性的影响总体上并不明显,但对箨叶氮代谢酶活性有明显影响。
3.3 覆盖对雷竹笋食味品质的影响
由图2可知,覆盖后雷竹笋可溶性糖含量和糖酸比及甜味氨基酸含量及其比例均显著升高,升幅分别为43.64%、89.53%、82.72%和118.89%,竹笋总酸、草酸、纤维素和苦味、芳香类氨基酸含量及鲜味氨基酸含量及其比例均显著降低,降幅分别为24.47%、32.60%、13.20%、21.10%、12.49%、68.25%和62.00%,而其他食味品质指标间无显著变化。以上结果表明,覆盖栽培可以显著提高竹笋甜味,降低竹笋苦味、酸涩味和粗糙度,明显改善竹笋食味品质。

表1 雷竹箨叶碳氮代谢酶活性
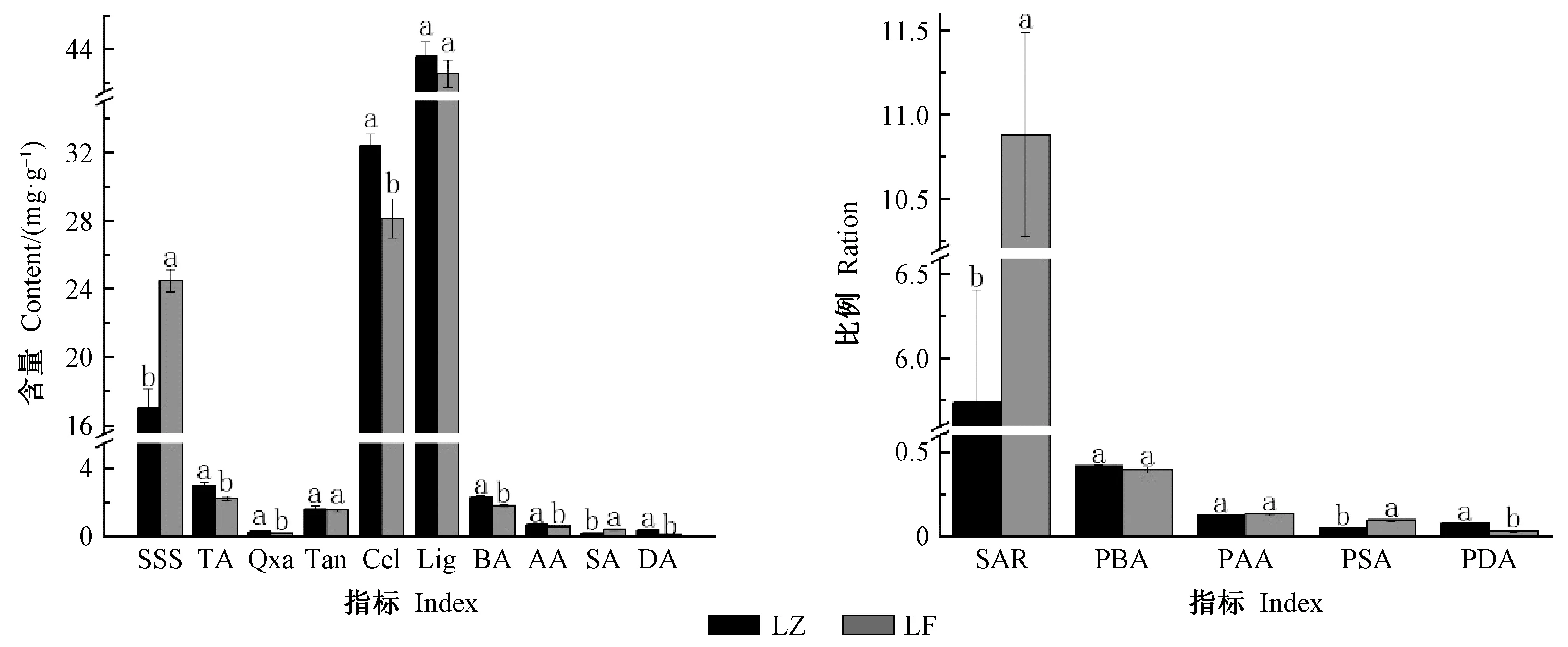
图2 雷竹笋呈味物质和粗糙度物质
3.4 雷竹笋箨叶碳氮代谢指标与食味品质的RDA分析
除趋势对应分析(detrended correspondence analysis,RDA)结果如图3所示,雷竹笋箨叶碳氮代谢指标对竹笋食味品质累计解释率为87.71%,可以较好的解释箨叶碳氮代谢指标与竹笋呈味物质和粗糙度物质的关系。其中,箨叶N含量与竹笋总酸、草酸、纤维素、苦味氨基酸、芳香类氨基酸含量和鲜味氨基酸含量及其比例间两两呈显著正相关; 而与可溶性糖含量、糖酸比和甜味氨基酸含量及其比例间两两呈显著负相关; 箨叶可溶性糖、淀粉、NSC含量则相反。蔗糖合成酶、硝酸还原酶、谷氨酸合酶、谷氨酰胺合成酶活性与竹笋可溶性糖含量、糖酸比和甜味氨基酸含量及其比例间两两呈显著正相关; 而与竹笋总酸、草酸、纤维素、苦味氨基酸、芳香类氨基酸含量和鲜味氨基酸含量及其比例间两两呈显著负相关; 淀粉酶、蔗糖磷酸合成酶、酸性转化酶和中性转化酶与竹笋食味品质指标间无明显相关性。说明覆盖雷竹笋可以通过调节箨叶养分含量和碳氮代谢酶活性以适应环境条件的剧烈变化,同时对竹笋呈味物质和粗糙度物质产生影响,进而影响竹笋食味品质。

图3 雷竹箨叶碳氮代谢指标与竹笋食味品质的RDA分析
4 讨论
植物通常按照自身物质合成需求调节其养分元素的含量,植物C、N 含量反映了植物应对外界环境条件的生态策略和适应特征(杜尧等, 2014; 陈珊等, 2014b)。本研究中,雷竹笋箨叶N含量对覆盖栽培的响应敏感,而箨叶C含量无明显变化,引起N含量显著降低的原因可能是覆盖栽培下雷竹生长速率加快,其干物质含量累积较多,竹鞭和根系的养分传输能力不足以支撑其高生长速率进程,一定程度上也引起了养分稀释效应(Yangetal., 2006; 郭子武等, 2013a); 而C在植物体内起骨架作用,为植物生理代谢活动提供糖类等物质,其含量有较好的稳定性(许雪贇等, 2018)。
NSC是植物碳供应水平的重要度量指标,可以反映植物的生长状况及其应对外界环境胁迫的缓冲能力(Myersetal., 2007),其中淀粉是植物主要的储备物质,而可溶性糖是碳水化合物运输和转化的主要形式,可被随时调动参与植物的生理代谢活动(郭子武等, 2015a)。本研究中,覆盖笋箨叶可溶性糖、淀粉和NSC含量均显著升高,说明覆盖栽培下雷竹笋用于自身生长代谢和储存的非结构性碳显著增加,竹笋生长活性增强,这可能与覆盖条件下环境的剧烈变化和有机覆盖物分解提高了林地土壤养分有效供给能力和微生物活性等密切相关(陈珊等, 2014a)。可见,雷竹笋主要通过调节箨叶N和非结构性碳水化合物含量来提高竹笋对覆盖栽培环境的适合度。
植物蔗糖的合成和分解依赖于蔗糖合成酶、磷酸蔗糖合成酶以及蔗糖转化酶等的协同调控,而硝酸还原酶是调控植物氮素同化速率的首个关键酶,谷氨酸合酶/谷氨酰胺合成酶循环是植物氮同化的代谢中心(吴能表等, 2014; Leaetal., 2003)。本研究发现,雷竹覆盖笋箨叶蔗糖合成酶活性显著提高,一方面可能是由于覆盖下竹笋处于避光环境,竹笋通过采取提高糖代谢的方式减少能量消耗,另一方面可能是光照不足的条件下蔗糖的生产和消耗加剧所致(孙恒等, 2020)。氮代谢酶活性与土壤养分、水分等密切相关,覆盖栽培可以明显提高土壤硝态氮、铵态氮以及土壤水分含量(叶莉莎等, 2016),进而影响箨叶氮代谢速率,这也说明覆盖栽培可以明显提高箨叶碳氮同化能力,尤其是氮素同化能力。
覆盖栽培后竹笋处于避光环境,且土壤水分相对充足,笋芽处于“暗形态建成”阶段,一定程度上促进糖类物质的合成,而抑制纤维类、总酸和草酸等的合成(徐森等, 2021; 郭子武等, 2015b); 同时覆盖栽培下土壤养分、温度等发生变化,可能影响了竹笋对氮素的吸收转运及代谢过程,进而改变竹笋氨基酸组分含量及其比例(李明良等, 2015)。进一步分析雷竹笋箨叶碳氮代谢特征与竹笋食味品质的关系,箨叶N含量与竹笋酸味、苦味、鲜味、粗糙类和芳香类物质含量呈显著正相关,而与竹笋甜味物质含量呈显著负相关,这与王荣萍等(2013)在无核黄皮(Clausenalansium)上的研究结果和哈地尔·依沙克等(2018)对灰枣(Ziziphusjujuba)的研究结果相反,其原因可能与覆盖栽培明显提高竹笋生长活性,导致其总生物量较高从而引起N的稀释有关。箨叶可溶性糖、淀粉、NSC含量和蔗糖合成酶、硝酸还原酶、谷氨酸合酶、谷氨酰胺合成酶活性与竹笋甜味物质含量呈显著正相关,而与竹笋酸味、苦味、鲜味、粗糙类和芳香类物质含量呈显著负相关。碳代谢是碳水化合物转化与积累的重要环节,而氮代谢是合成蛋白质和氨基酸的主要途径(宁宇, 2019),覆盖栽培下,环境水分、温度和光照等的变化影响雷竹笋碳氮代谢效率,促进甜味物质等合成,而竹笋出土后受光照影响,单宁和草酸等酸涩类物质合成加剧,同时出土后竹笋组织老化速度加快,粗糙度提高(时俊帅等, 2019b),进而导致覆盖笋食味品质的明显提升。可见,雷竹笋通过调节箨叶主要养分含量和碳氮代谢酶活性来适应覆盖条件下环境的剧烈变化,从而使竹笋食味品质也发生明显协同变化。
5 结论
雷竹笋通过调节箨叶碳氮代谢来提高竹笋对覆盖栽培环境的适合度,且林地覆盖栽培下竹笋甜味明显提高,酸涩味和粗糙度明显降低,竹笋食味品质明显改善。同时,雷竹箨叶养分含量和氮代谢酶活性与竹笋食味品质密切相关,覆盖下雷竹箨叶碳氮代谢特征及竹笋食味品质表现出协同变化关系。
猜你喜欢
学苑创造·A版(2021年10期)2021-10-30
林业科学研究(2021年2期)2021-04-10
粮油食品科技(2020年6期)2020-12-08
红楼梦学刊(2020年2期)2020-02-06
幼儿画刊(2019年8期)2019-11-04
科学与财富(2019年24期)2019-08-06
世界竹藤通讯(2018年3期)2018-06-22
意林·少年版(2017年4期)2017-03-07
—— 品尝员的识别能力与适口性
北方水稻(2016年4期)2016-01-09
天然产物研究与开发(2014年8期)2014-04-27
