
山桐子健康和感病植株不同器官区细菌群落结构、多样性与功能预测分析*
2022-12-23 09:04薛晓焱蔡齐飞耿晓东周慧娜李明婉王艳梅
林业科学 2022年8期
李 志 薛晓焱 刘 震 蔡齐飞 耿晓东 冯 建 周慧娜 张 涛 李明婉 王艳梅
(河南农业大学林学院 郑州 450002)
山桐子(Idesiapolycarpa)是大风子科(Flacourtiaceae)山桐子属落叶阔叶乔木,树形优美,木材松软,果实艳丽,在不同立地环境均具有较好的适应性,是良好的新型用材树种和观赏树种(陈珺肄等, 2019)。山桐子果实含油率高且营养物质丰富,其果肉和种子中不饱和脂肪酸含量可达到75%和85%以上,是优良的木本油料树种,可作为开发高档保健型木本食用油的原料,素有“树上油库”的美誉(代莉, 2014)。山桐子作为木本油料树种,具有抗逆性强、适生地广、“不与粮争地”的优势,在林业、食品、医药及生态等多个领域有巨大的开发前景(陈钊等, 2019; 房丽莎等, 2020)。近年来,国家正式制定颁布了山桐子、山桐子油、山桐子饼粕3个行业标准,并将山桐子纳入到了国家储备林建设的政策体系中。
在自然界,植物生长环境中有大量微生物,寄主在植物宿主体内或附着其表面,这些微生物群体统称为植物微生物组。细菌是这些微生物群落中最丰富的类群(Mülleretal., 2016)。同种植物不同器官区微生物存在一定差异,李聪聪等(2020)比较了高寒草甸优势植物叶内、根内、土壤原核微生物组的组成,发现植物器官对微生物具有强烈选择性,只有部分土壤微生物能够进入植物根内和叶内; Wu等(2020)研究结果表明柑桔(Citrusreticulata)叶片和韧皮部的微生物群落数量远低于根系和土壤。细菌与植物形成复杂、动态的关联,包括互利共生或致病,Zarraonaindia等(2015)的研究发现葡萄(Vitisvinifera)叶子和果实中的甲基杆菌(Methylobacteriumspp.)可以通过产生植物激素的生产来刺激植物的发育; 尹诗琳等(2021)的研究发现,芽孢杆菌属(Bacillus)对桑树(Morusalba)病原菌存在着某种程度的拮抗作用,而泛菌属(Pantoea)可能对桑轮纹病的发生有协同或者伴生作用。
山桐子林木出现感病现象,部分植株根系变黑腐烂,树皮出现褐色水渍病斑,叶片边缘枯萎且叶面有褐色斑点,病害发生原因及致病因子尚未明确。目前,山桐子相关研究主要集中在栽培技术、油脂提取、开发利用等方面,而微生物病害方面的研究相对薄弱。 李佛生等(2021)对山桐子内生真菌进行分离,鉴定出4种山桐子组织来源共同的内生真菌为芽短梗霉(Aureobasidiumpullulans); 有研究报道了山桐子叶斑病的病原真菌链格孢菌(Alternariaalternata)(Sunetal., 2015)。 细菌是否为造成山桐子病害的关键因子还有待进一步探索。研究基于高通量测序技术,以健康和感病山桐子植株为对象,探究山桐子土壤、树皮及叶片中细菌群落组成和多样性,分析其差异,探讨健康和感病山桐子不同器官区的优势菌属及其潜在功能,探究各器官区的关键菌属与山桐子感病之间的关系,以期为山桐子病害防治及经营管理提供理论基础。
1 材料与方法
1.1 试验地概况
研究试验地位于河南农业大学林业试验站,属于暖温带大陆性气候,年平均气温14.2℃,极端最高气温43℃,极端最低气温-17.9℃,年平均降水量650.1mm,全年日照时数约2 400 h。试验地0~20cm土壤的基础化学性质为: pH 8.0,土壤全碳(TC)17.76 g·kg-1,全氮(TN)0.81 g·kg-1,有效磷(AP)2.43 mg·kg-1,有效钾(AK)75.78 mg·kg-1。
1.2 样品采集
2020年5月,在样地内采集健康山桐子植株和感病山桐子植株的根际土壤、树皮、叶片样品,以及样地非根际土壤样品。土壤样品采集时,每样株随机选取3个样点,去除植株基部表层土壤及杂物,取5~20cm土层侧根或须根表面0.4cm左右的土壤作为根际土壤,空地同土层随机取3个样点作为非根际土壤,将每个样点采集的土壤分别装入无菌自封袋中充分混匀,共计9份土样,土壤样品编号为健康植株根际土壤(G)、感病植株根际土壤(BG)、非根际土壤(FG)。用无菌剪刀和镊子采集植物树皮和叶片,每样株2个部位分别采3份,共6份皮样、6份叶样,编号分别为健康植株树皮(P)、感病植株树皮(BP)、健康植株叶片(Y)和感病植株叶片(BY)。采集好的样品用冰盒保存带回实验室后,置于-80℃冰箱保存,供检测使用。
1.3 测定指标及方法
对健康、感病山桐子植株根际土壤、树皮、叶片及非根际土的总DNA进行提取,提取的DNA用1.2%琼脂糖凝胶进行电泳,用紫外分光光度计检测其质量和浓度。选择引物338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′),对样品细菌16 S rRNA 基因的V3-V4 区进行PCR 扩增(Claessonetal., 2009)。采用的25μL扩增体系包括5×reaction buffer 5μL,5×GC buffer 5μL,dNTP(2.5mmol·L-1)2μL,Forwardprimer(10μmol·L-1)1μL,Reverseprimer(10μmol·L-1)1μL,DNA Template 2μL,ddH2O 8.75μL,Q5 DNA Polymerase 0.25μL。扩增参数为98℃ 2min,98℃ 15 s,55℃ 30 s,72℃ 30 s,72℃ 5min,30个循环,10℃保存。Illumina miseq测序平台进行高通量测序,测序由上海派森诺生物科技有限公司完成。
1.4 数据处理
利用QIIME2(2019.4)软件调用DADA2进行质控、去噪、拼接、去嵌合体,采用97%的序列相似度作为OTU(operational taxonomic unit,OTU)的划分阈值,获得的OTU代表序列用于细菌群落组成分析、α多样性分析和β多样性分析、菌群功能预测等; 运用R软件等工具,绘制稀疏曲线; 根据ASV/OTU丰度表制作upsetR图; 对样本中相对丰度前20位的属进行聚类分析并绘制热图; 根据样本差异距离进行主坐标分析(Principal coordinate analysis,PCoA); 利用SPSS 24.0分析α多样性指数在各样本间的差异(P<0.05)。
2 结果与分析
2.1 微生物测序结果分析
对山桐子林地非根际土壤及健康、感病山桐子植株的根际土壤、树皮、叶片进行高通量测序,21个样本(3个重复/处理)中的细菌16 S rRNA V3-V4序列过滤后,共获得1 193 103条高质量序列。稀疏曲线可以用来比较测序数量不同的样本物种丰富度,也可说明样本的取样大小是否合理。根据样本的测序数据,绘制对应的稀疏曲线(图1)。结果表明,各样本的稀疏曲线均趋于平缓,说明该测序深度包含了样本中绝大多数细菌类型,测序深度充足,可以反映真实的细菌群落生物多样性。
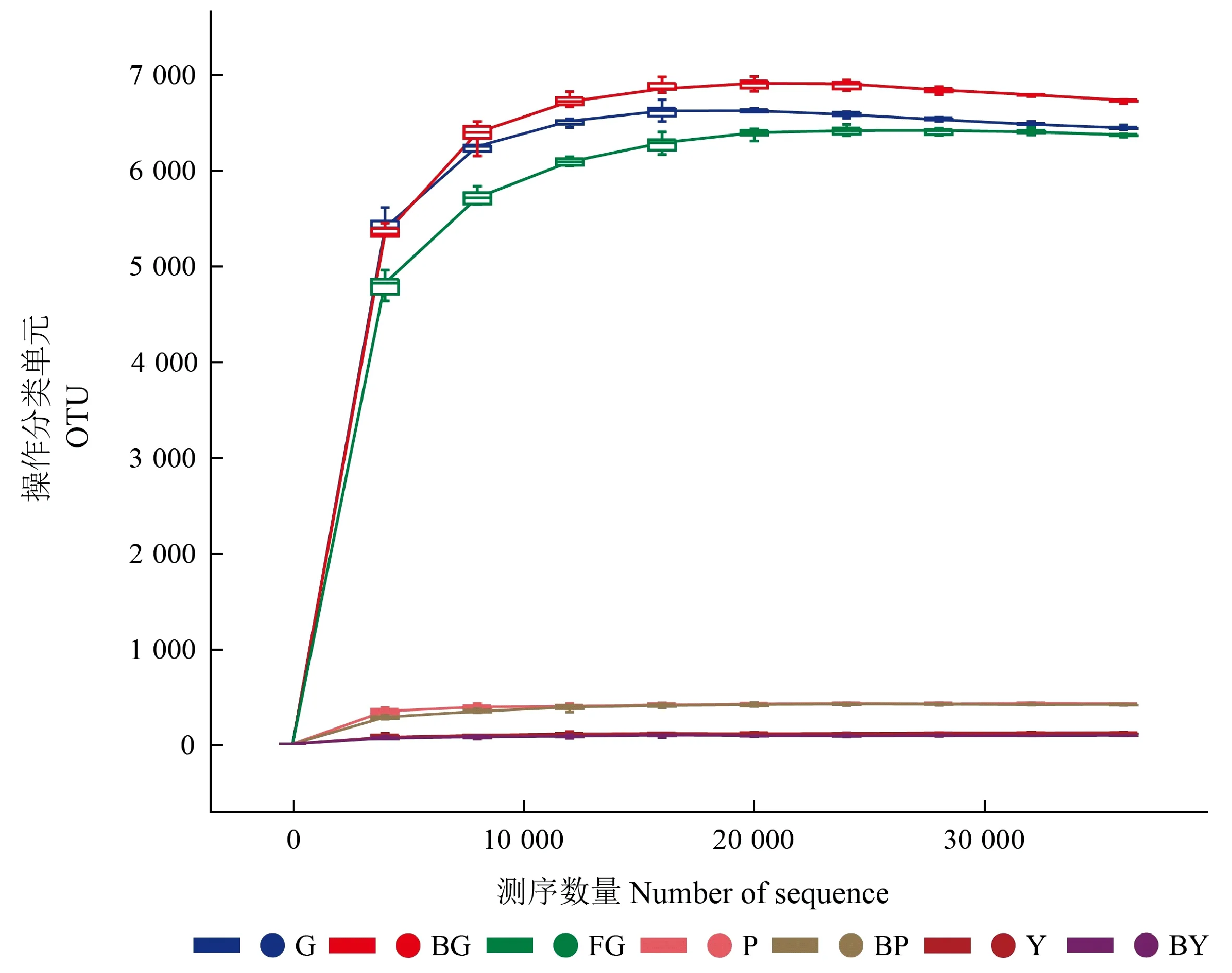
图1 各区域样本中细菌稀释曲线
2.2 山桐子健康、感病植株不同器官区细菌群落结构
2.2.1 山桐子健康、感病植株不同器官区OTU数 使用upset图分析山桐子不同器官区细菌的物种组成(图2)。土壤样本健康植株根际土壤感病植株根际土壤(G)、感病植株根际土壤(BG)、非根际土壤(FG)中包含的OTU 数目分别是17 236、17 403、15 461 个,3组共有OTU 数量为1 100个,各自特有OTU 数目分别是14 517、14 577、12 776个,根际土壤样本特有的OTU 数均高于非根际土壤,预示着根际土壤中含有较多的特有微生物种类; G和BG两组特有OTU分别占其总OTU的84.22%和83.76%。树皮样本P、BP中的特有OTU数量为737、857个,分别占其总OTU的74.29%和80.77%。叶片样本Y、BY中特有OTU数量为181、156个,分别占其总OTU的69.35%和70.91%。以上数据表明,健康植株根际土壤和树皮中特有OTU数量少于感病植株,叶片特有OTU数量多于感病植株,7组样本中特有OTU数量均占较大比例。
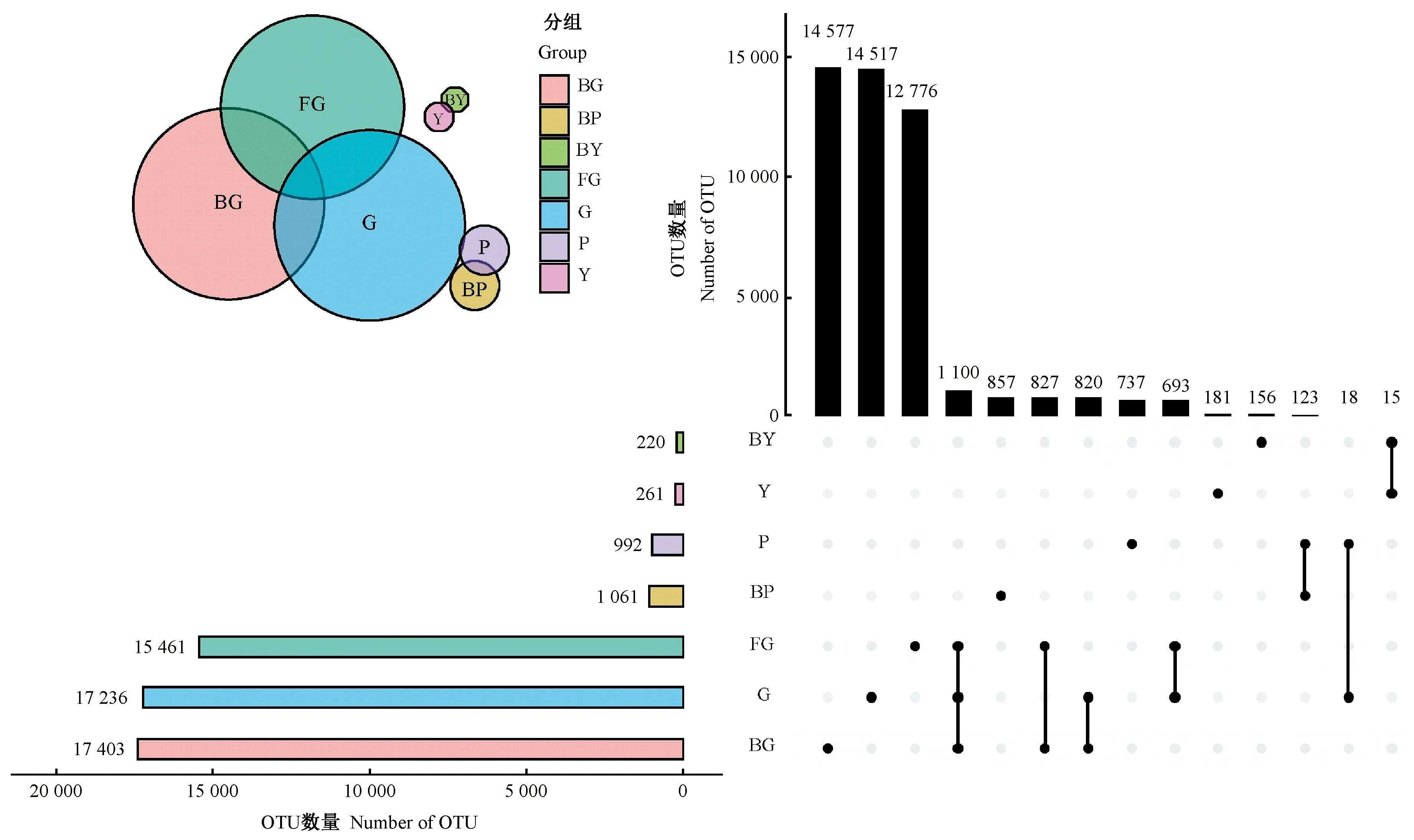
图2 各器官区细菌群落
2.2.2 山桐子健康、感病植株不同器官区细菌门、纲、目、科水平群落结构 对各样本中得到的OTU序列进行分类注释,所有样本的细菌OTU隶属于37个门,118个纲,291个目,511个科,1051个属,2772个种。
各分类水平上物种相对丰度前10位的细菌分布见图3。在门水平上(图3a),土壤样本的优势菌门为变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria); 其中变形菌门在G、BG和FG中所占比例为BG>G>FG,在根际土壤中相对丰度较大; FG中的酸杆菌门和绿弯菌门(Chloroflexi)占比数量显著高于根际土壤(P<0.05),且绿弯菌门为FG的优势菌门。树皮、叶片样本的优势菌门为蓝细菌门(Cyanobacteria)和变形菌门。在树皮样本中,变形菌门在P、BP中的相对丰度分别为30.52%、62.26%,P的相对丰度显著低于BP; 蓝细菌门在P、BP中的相对丰度分别为61.57%、28.39%,P的相对丰度显著高于BP(P<0.05)。在叶片样本中占比最多的是蓝细菌门84.58%(Y)、88.42%(BY),其次是变形菌门15.16%(Y)、11.33%(BY),2个门的细菌占总数量的99.7%以上。
在纲水平上(图3b),变形菌门中α-变形菌纲(Alphaproteobacteria)、γ-变形菌纲(Gammaproteo-bacteria)和放线菌门中放线菌钢(Actinobacteria)在G和BG中占比较为接近,但均高于FG,其中放线菌纲在根际与非根际土壤中差异达到显著水平(P<0.05); 而放线菌门中嗜热油菌纲(Thermoleophilia)在FG中的相对丰度显著高于根际土壤。在树皮样本中,蓝细菌门中产氧光细菌纲(Oxyphotobacteria)在P(61.57%)中的相对丰度显著高于BP(28.39%),γ-变形菌纲在BP(43.82%)中的相对丰度显著高于P(5.15%)。纲水平上,叶片样本主要纲为产氧光细菌纲和α-变形菌纲,两者在Y和BY中占比接近。
在目(图3c)、科(图3d)水平上,产氧光细菌纲中叶绿体(Chloroplast)是树皮、叶片的主要组成,在P、BP、Y、BY中占比分别为61.56%、28.39%、84.54%、88.41%; α-变形菌纲中立克次氏体目(Rickettsiales)及其下属线粒体(Mitochondria)的相对丰度在P、Y、BY 3组样本中占比分别为14.03%、14.06%和10.76%; γ-变形菌纲中肠杆菌目及其下属肠杆菌科(Enterobacteriaceae)在树皮样本BP(41.03%)中的相对丰度显著高于P(1.02%)。
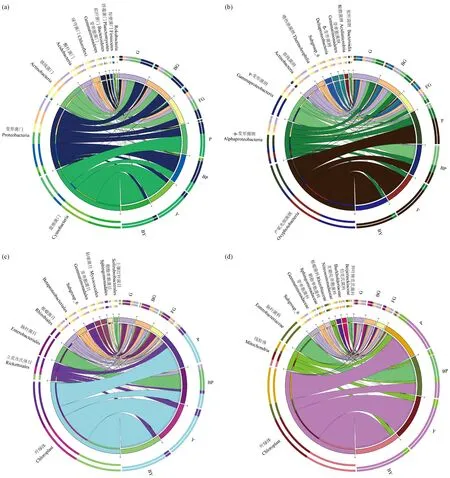
图3 各分类水平上相对丰度前10物种在各样本的分布和丰度
2.2.3 山桐子健康、感病植株不同器官区细菌属水平群落结构 对3组土壤样本中相对丰度排名前20位的细菌属进行UPGMA聚类分析,结果表明(图4),根际土壤G、BG具有相似的细菌群落结构,聚为一类; FG与根际土壤差异较大,单独为一类。20个细菌属可以分为3组(Ⅰ、Ⅱ和Ⅲ)。FG中Ⅰ组细菌属丰度较高,主要包括变形菌门中的bacteriap25、Haliangium、MND1; 放线菌门中的Gaiella、MB-A2-108、67-14、IMCC26256; 绿弯菌门中的A4b、KD4-96; 酸杆菌门中的Subgroup_17、RB41; 以及浮霉菌门的OM190,Rokubacteria的Rokubacteriales,共13个菌属。G中Ⅲ组细菌属丰度较高,主要包括放线菌门中的类诺卡氏菌属(Nocardioides)、芽球菌属(Blastococcus)、土壤红杆菌属(Solirubrobacter); 酸杆菌门中的Subgroup_6; 变形菌门的鞘氨醇单胞菌属(Sphingomonas),共5个菌属。BG中Ⅱ组细菌属丰度较高,主要包括变形菌门中Dongia以及放线菌门中的链霉菌属(Streptomyces),共2个菌属。
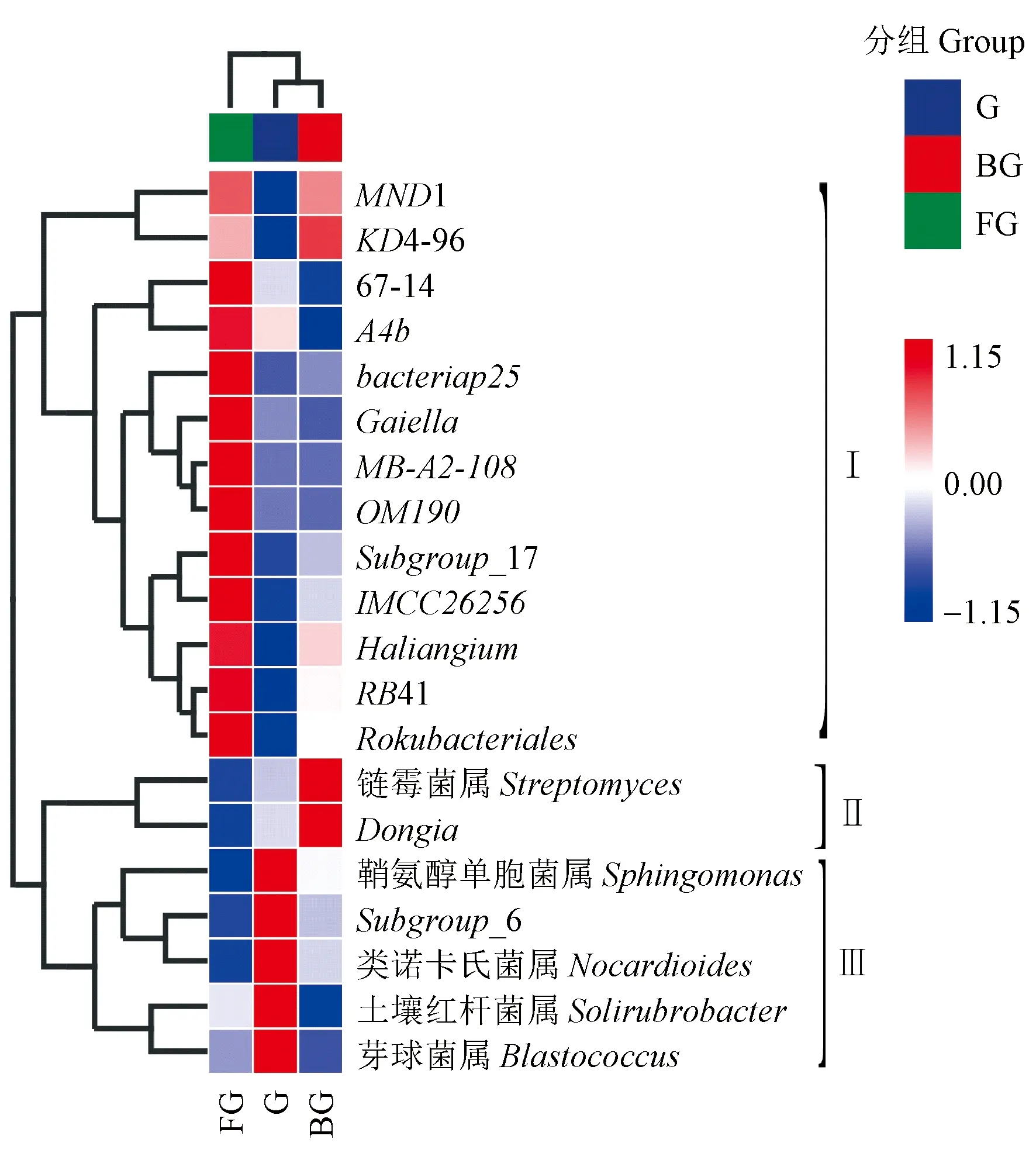
图4 土壤在属水平上的细菌丰度聚类热图
选取2组树皮样本中丰度排名为前20位细菌属进行UPGMA聚类,绘制物种丰度聚类热图(图5)。由图5可知,2组树皮中的细菌群落在属水平上差异明显。P中丰度较高的细菌属,主要包括变形菌门中的鞘氨醇单胞菌属、不动杆菌属(Acinetobacter)、Aureimonas; 放线菌门中的Quadrisphaera、贫养杆菌属(Modestobacter)和拟杆菌门的薄层菌属(Hymenobacter),共6个菌属。BP中丰度较高的细菌属,主要包括变形菌门中的Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、果胶杆菌属(Pectobacterium)、甲基杆菌属(Methylobacterium)、假单胞菌属(Pseudomonas)、小坂菌属(Kosakonia)、Brenneria、Hafnia-Obesumbacterium、玫瑰单胞菌属(Roseomonas); 厚壁菌门中的类芽胞杆菌属(Paenibacillus)、丹毒丝菌属(Erysipelothrix)、Saccharibacillus; 放线菌门的芽球菌属,共12个菌属。
对树皮样本中的细菌进行优势菌属分析,在健康和感病山桐子树皮细菌中,细菌丰度10%以上且存在显著差异(P<0.05)的属有2个,分别是Brenneria和Hafnia-Obesumbacterium,在健康山桐子树皮中相对丰度仅为0.79%和0,显著低于感病山桐子树皮中Brenneria(18.33%)和Hafnia-Obesumbacterium(17.5%)的相对丰度。
选取2组叶片样本中丰度排名为前20位细菌属进行UPGMA聚类(图6)。由图6可知,Y中丰度较高的细菌属,主要包括变形菌门中的鞘氨醇单胞菌属、Burkholderia-Caballeronia-Paraburkholderia、短波单胞菌属(Brevundimonas)、不动杆菌属、慢生根瘤菌属(Bradyrhizobium)、肠杆菌属(Enterobacter)、Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium、假单胞菌属、贪铜菌属(Cupriavidus)、草螺菌属(Herbaspirillum); 以及绿弯菌门的KD4-96,共11个菌属。BY中丰度较高的细菌属,主要包括厚壁菌门中的魏斯氏菌属(Weissella)、芽孢杆菌属、乳杆菌属(Lactobacillus)、链球菌属(Streptococcus); 以及变形菌门中的Escherichia-Shigella、Phreatobacter、Pusillimonas,共7个菌属。
2.3 不同器官区细菌群落多样性分析
2.3.1 α多样性分析 对山桐子植株不同器官区细菌多样性指数进行差异分析(表1)。采用Chao1指数反映物种丰富度; 采用Shannon指数和Simpson指数反映细菌群落的多样性; Goods coverage指数反映群落覆盖度。山桐子植株不同器官区细菌的Chao1、Shannon、Simpson指数值都表现为土壤样本最高,其次是树皮样本,最后是叶片样本,说明土壤细菌群落丰富度和多样性是最高的。土壤各组的Chao1指数由大到小排列依次为: BG>G>FG,感病植株根际土壤细菌群落丰富度最高; Shannon指数和Simpson指数由大到小依次为: G>FG>BG,说明健康植株根际土壤细菌群落多样性最高。叶片中Y组的Chao1、Shannon、Simpson指数均高于BY,表明健康植株叶片细菌群落丰富度和多样性高于感病植株叶片。但健康植株与感病植株土壤、叶片细菌群落的Chao1指数、Shannon指数和Simpson指数差异均未达到显著水平(P>0.05)。树皮细菌群落Chao1指数表现为P高于BP,而Shannon、Simpson指数均表现为BP高于P,其中Simpson指数差异达到显著水平(P<0.05),说明健康植株树皮细菌群落丰富度较高,但细菌群落多样性低于感病植株树皮。

图5 树皮在属水平上的细菌丰度聚类热图
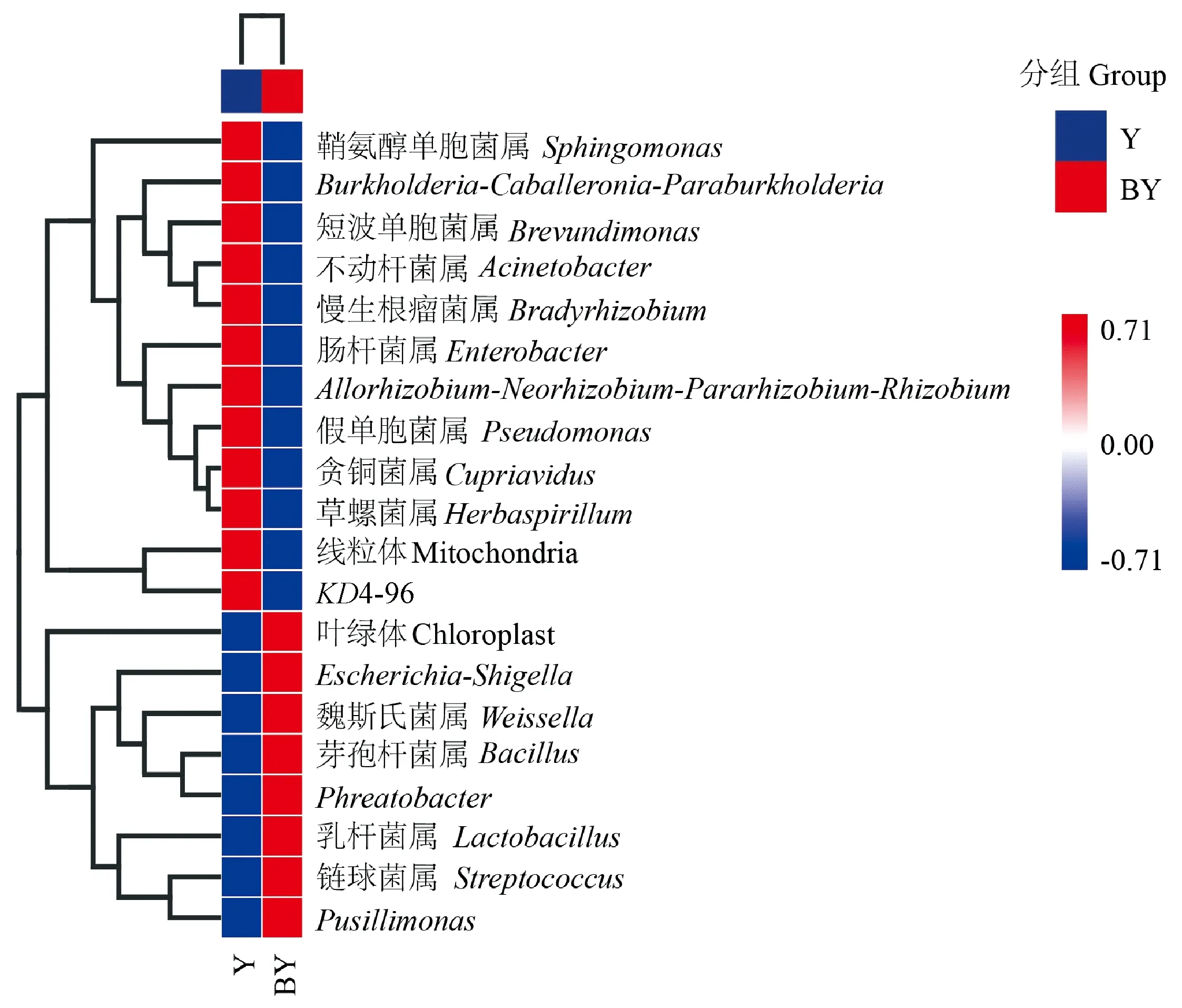
图6 叶片在属水平上的细菌丰度聚类热图
2.3.2 β多样性分析 为进一步明晰样本间在群落物种组成上的差异性,采用PCoA分析山桐子不同器官区细菌群落物种组成的相似度。如图7所示,PCo1和PCo2分别解释变量方差的42.9%、8.6%,两者累计贡献率达51.5%。PCo1可以将土壤的细菌群落与树皮、叶片明显区分开,土壤分布在PCo1负方向,树皮和叶片主要分布在正方向,说明树皮和叶片与土壤细菌群落差异较大。不同健康状态山桐子的土壤和树皮细菌群落结构存在差异性,其中健康、感病树皮细菌组成差异明显。而Y和BY相聚较近,表明健康和感病山桐子叶片细菌组成相似度较高。
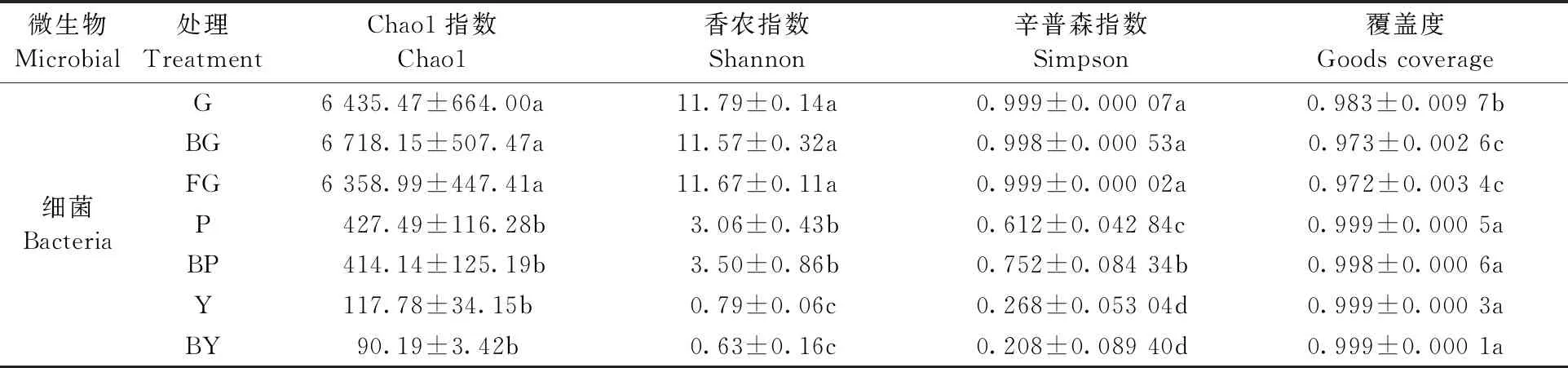
表1 不同器官区细菌Alpha多样性指数①

图7 不同器官区细菌PCoA分析
2.4 菌群代谢功能预测分析
使用PICRUSt2软件对健康、感病山桐子中不同器官区的细菌微生物群落进行代谢功能预测。通过KEGG数据库比对,结果共注释到6条一级代谢通路,包括生物进程(cellular processes)、环境信息处理(environmental information processing)、遗传信息处理(genetic information processing)、人类疾病(human diseases)、代谢(metabolism)和生物体系统(organismal systems); 一级代谢通路中代谢、遗传信息处理为主要代谢通路,占全功能分类的80.68%~82.24%、11.26%~13.48%。在二级代谢通路中共注释到34种代谢通路子功能,选取丰度占总丰度比值大于1.00%的子功能进行统计(表2)。山桐子不同器官区的微生物群落功能相似,但在子功能通路丰度上存在较大差异; 感病、健康山桐子的代谢功能,在土壤和叶片样本中组内差异较小,树皮样本组内差异较大。细胞进程中的细胞运动(0.64%~2.72%)和环境信息处理中的膜运输(1.58%~2.32%),在子功能通路丰度上呈P低于BP。遗传信息处理中复制和修复(4.73%~4.87%)、折叠、分类和降解(3.08%~4.46%)、翻译(2.52%~3.22%)为主要子功能,在子功能通路丰度上呈P高于BP。与代谢有关的主要子功能有11种,在各样本中丰度均较高,其中5种子功能通路丰度为土壤>树皮>叶片,包括碳水化合物代谢(12.01%~13.21%)、氨基酸代谢(11.63%~13.13%)、萜类化合物和聚酮类化合物的代谢(7.02%~9.72%)、外源生物降解与代谢(4.49%~7.74%)、脂质代谢(4.06%~7.25%); 辅助因子和维生素的代谢(11.4%~16.58%)、能量代谢(5.01%~9.51%)2种子功能通路丰度表现为叶片高于土壤和树皮; 氨基酸代谢、能量代谢、聚糖生物合成与代谢、辅助因子和维生素的代谢、其他氨基酸代谢在子功能通路丰度上呈P高于BP; 脂质代谢、外源生物降解与代谢在子功能通路丰度上呈G高于BG,P低于BP。

表2 KEGG 数据库主要代谢通路丰度
对已有代谢通路的丰度数据进行分析,使用metagenomeSeq方法对样本组进行两两比较,分析相同器官区、不同样本之间在功能上的差异(图8),筛选条件为P<0.05。分析结果表明,在土壤中,BG组12,14和16元大环内酯的生物合成(Biosynthesis of 12-, 14-and 16-membered macrolides)、植物昼夜节律(Circadian rhythm-plant)、轴突导向(axon guidance)、倍半萜生物合成(sesquiterpenoid biosynthesis)、Ⅱ型聚酮主链的生物合成(biosynthesis of type II polyketide backbone)、Ⅱ型聚酮产品的生物合成(biosynthesis of type II polyketide products)、糖胺聚糖降解(Glycosaminoglycan degradation)和溶酶体(Lysosome)通路相关功能基因对照G组显著增加,而萘降解(Naphthalene degradation)通路相关功能基因对照G组极显著降低。在树皮中,BP组的D-精氨酸和D-鸟氨酸代谢(D-Arginine and D-ornithine metabolism)和金黄色葡萄球菌感染(Staphylococcus aureus infection)通路相关功能基因对照P组极显著增加,而类胡萝卜素生物合成(Carotenoid biosynthesis)、弓形体病(Toxoplasmosis)和志贺菌病(Shigellosis)相关功能基因对照P组显著降低。
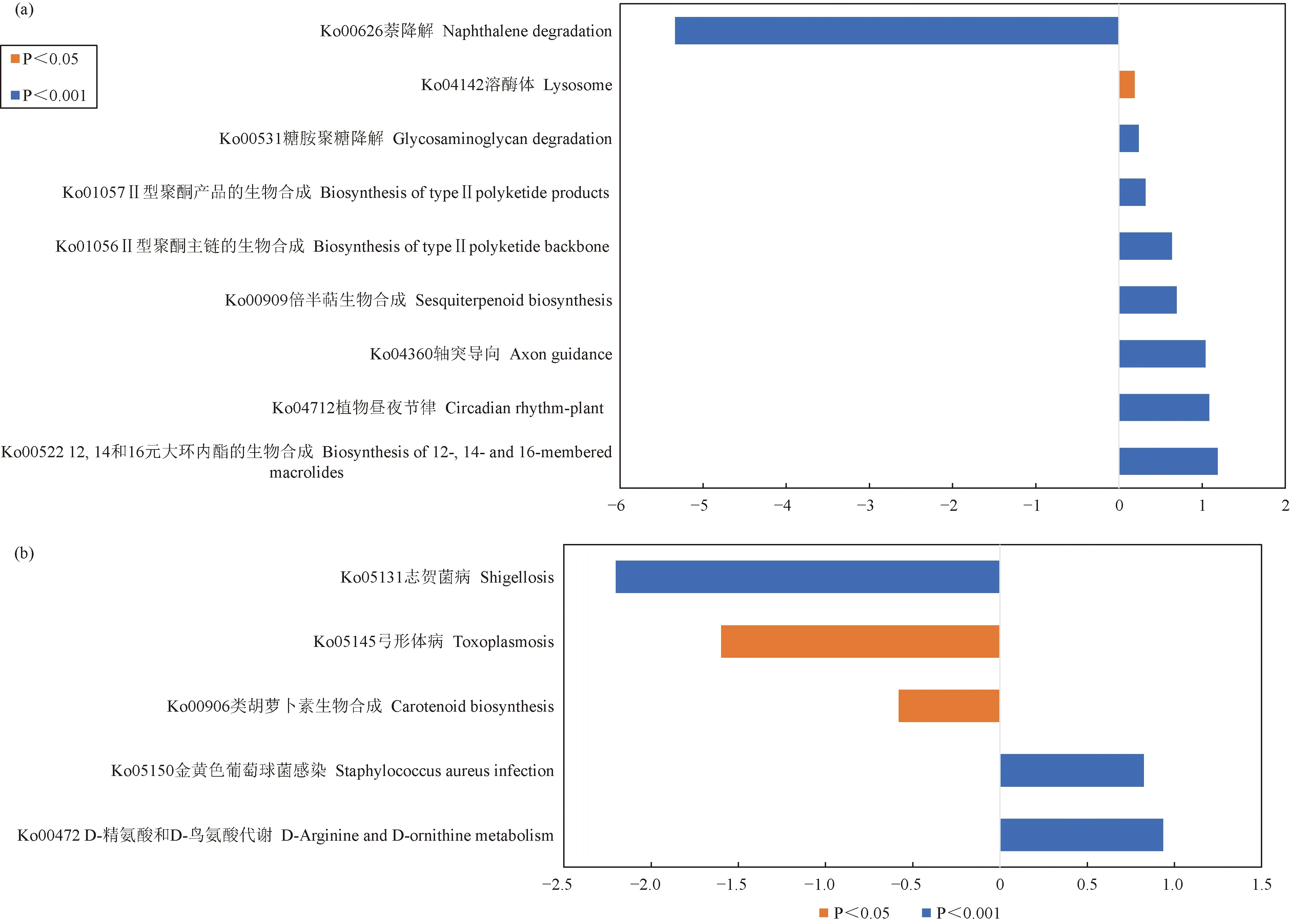
图8 两两组间代谢通路存在显著差异的功能分类
3 讨论
3.1 健康和感病山桐子各器官区细菌群落结构特征
本研究采用Illumina miseq高通量测序技术分析健康和感病山桐子土壤、树皮、叶片细菌群落结构和多样性的差异。结果表明,山桐子不同器官区细菌群落结构存在明显差异,OTU数量由高到低为土壤>树皮>叶片,土壤相较于植物组织含有较多的OTU数量,这与其他林木研究结果相似(刘海燕等, 2019; 杨多等, 2022),一定程度上也反映了不同器官区环境的特性。进一步的物种分析注释发现,山桐子不同器官区细菌群落的主要成分存在差异,在分类学上不同种类细菌的相对丰度发生变化。山桐子根际土壤细菌群落主要由变形菌门、放线菌门和酸杆菌门构成,这些优势菌门在根际环境中富集,与其较强的环境适应性有关(王安宁等, 2019; Lietal., 2020)。其中变形菌门包括多种病原菌和固氮菌,因为其外膜主要由脂多糖组成可以保护其内部遗传物质不受外部干扰,可以在大多数环境中生存并繁衍成为优势菌(葛艺等, 2021; 翟婉璐等, 2017)。山桐子树皮和叶片细菌群落主要由蓝细菌门和变形菌门构成,这一结果与前人的研究发现相似(暴家兵等, 2020; 李慧娟等, 2021)。蓝细菌门在山桐子叶际细菌中处于优势地位,相对丰度范围为84.58%~88.42%,产氧光细菌纲是其主要的细菌类群,有研究表明,蓝细菌是地球上广泛存在的一种自养型细菌,种类繁多,有些种类能够进行产氧的光合作用,有些种类在暗环境下也能生长,营养要求低,对极端环境适应性很强(王文晓等, 2020)。山桐子树皮和叶片中的微生物群落与土壤微生物群落差异较大,可能与不同器官区域的周边环境有关,蓝细菌能够为山桐子树皮、叶片区域提供更适宜生长的微环境,促进山桐子的生长。
3.2 健康和感病山桐子各器官区细菌群落差异菌属
健康和感病山桐子各器官区细菌在属水平有显著差异。本研究中鞘氨醇单胞菌属是健康山桐子各器官区共有的优势菌属,不动杆菌属是健康山桐子树皮、叶片的优势菌属,与抑病能力密切相关。鞘氨醇单胞菌属具有植物促生特性(陈兆进等, 2021),鞘氨醇单胞菌在降解有毒物质、抵抗病原菌(秦立金等, 2018)、促进植物生长(葛艺等, 2021)等方面的功能。不动杆菌属能产生生物表面活性剂和一些酶类物质,具有较高的促生、抗菌和降解有机污染物性能(程浅, 2020; 顾美英等, 2021)。从核桃根际土壤中分离到的2种不动杆菌属菌株,经研究发现对核桃细菌性疫病具有明显拮抗作用,可以抑制病原菌生长,进而提高植物的抗病性(涂昌等, 2020)。本研究从感病山桐子树皮中筛选出的优势菌属为肠杆菌科的Brenneria和Hafnia-Obesumbacterium,在感病山桐子树皮中的相对丰度均显著高于健康山桐子树皮。Brenneria是重要的林木病原细菌,该属细菌侵染林木树皮后会引起不规则的溃疡病斑,对经济林木生长有严重的威胁(Frutos, 2010)。国内外研究表明,Brenneria细菌会引起许多林木病害,例如Brennerianigrifluens是核桃树皮溃疡病的病原菌(Morettietal., 2007);Brenneriasalicis是柳树水纹病的病原菌(Martineetal., 2009);Brenneriaquercina和B.corticissp. nov.是中国欧美杨溃疡病的致病菌(杨莉, 2013; Lietal., 2019);Brenneriarubrifaciens可引起核桃深层树皮溃疡病(Amirsardarietal., 2017)等。Brenneria可能对山桐子树皮区域病害的发生有协同或伴生作用,但其是否为山桐子感病的致病菌仍需进一步的研究。Hafnia-Obesumbacterium近年来已经被证明某些细菌具有较高的耐盐性(王改萍等, 2021),而Qing等(2020)研究发现Hafnia-Obesumbacterium在感病林蛙肠道菌群中的相对丰度较高,可能与动物腹泻相关。但是,对于Hafnia-Obesumbacterium在植物致病性方面的报道还较少,有待进一步探究。
3.3 健康和感病山桐子各器官区细菌群落多样性
本研究对不同器官区山桐子细菌群落多样性分析可知,山桐子各器官区细菌群落多样性和丰富度由高到低均表现为土壤>树皮>叶片,这与Wu等(2020)的研究结果类似,表明山桐子不同器官区细菌群落在多样性和丰富度方面存在显著差异。本研究中,感病山桐子树皮细菌多样性高于健康植株,树皮细菌丰富度低于健康植株,这与林马水等(2019)以健康和干腐病山核桃植株为研究对象,得出感病山核桃(Caryacathayensis)树皮多样性及丰富度指数均低于健康植株的结果不同,这可能是由于病菌在寄主体内存在时间的长短,对树体生态平衡造成的影响不同; 同时,植株发病时间的长短也可能是造成感病山桐子树皮组内细菌多样性差异较大的主要原因。
3.4 健康和感病山桐子各器官区细菌功能特征
菌群代谢功能预测结果表明,山桐子中的细菌主要涉及代谢、遗传信息处理等6条一级代谢通路和34种子功能,表现出功能上的丰富度。山桐子不同器官区样本中所含的功能基因种类大致相同,说明山桐子相关细菌群的功能基因具有一定相似性。对山桐子不同器官区样本细菌功能基因相对丰度进行分析,发现代谢通路在山桐子不同器官区中相对丰度均最大。在代谢通路中,代谢功能注释相对丰度大多表现为土壤微生物组含有更多的碳水化合物代谢、氨基酸代谢、萜类化合物和聚酮类化合物的代谢、外源生物降解与代谢和脂质代谢,这可能与土壤益生菌的含量有关(马欣等, 2021),其中碳水化合物代谢调控着生物体内碳水化合物的代谢形成、分解和互相转化,有益于植物的氮、磷循环(马欣等, 2021; 丁钰珮等, 2021); 氨基酸代谢与碳代谢和氮代谢相关(南镇武等, 2021); 萜类化合物代谢与植物抗病性和抗逆性有关(丛微等, 2021),间接说明土壤细菌代谢功能活跃有利于植物生长发育。而辅助因子和维生素的代谢、能量代谢在叶片微生物组中注释丰度明显高于土壤和树皮,这2种代谢功能可能在维持植物叶际环境中发挥重要功能。通过代谢通路差异分析可知,感病山桐子根际土壤和树皮中相关生物合成与氨基酸代谢的功能基因富集,说明感病后山桐子感病区域微生物群落的结构及其功能已做出了有益于生长发育的响应。而感病山桐子树皮中金黄色葡萄球菌感染功能基因富集,造成代谢紊乱,降低山桐子抗病性,进而导致山桐子病害的发生。因PICRUSt功能预测分析只是初步预测相关细菌的功能,未来的研究将需要进一步验证山桐子的致病菌,明晰引起山桐子病害的机制,更好地为山桐子病害防治及高质量经营提供理论支撑。
4 结论
健康和感病山桐子植株根际、树皮及叶片细菌结构组成、多样性和功能存在差异。山桐子土壤主要优势菌门为变形菌门、放线菌门和酸杆菌门,树皮和叶片主要优势菌门为蓝细菌门和变形菌门。健康山桐子各器官区中主要的优势菌属为鞘氨醇单胞菌属和不动杆菌属,感病后群落结构发生改变,健康树皮与感病树皮细菌群落结构差异显著,感病山桐子树皮的主要优势菌属为肠杆菌科的Brenneria和Hafnia-Obesumbacterium。功能预测表明山桐子各器官区细菌生态功能较为丰富,健康和感病山桐子的土壤及树皮区域代谢功能有明显差异。
猜你喜欢
童话王国·文学大师班(2022年3期)2022-04-19
中国林业产业(2020年4期)2020-06-02
中国油脂(2019年7期)2019-08-22
湖北农业科学(2019年11期)2019-07-22
小哥白尼(野生动物)(2018年4期)2018-08-01
中国糖料(2018年4期)2018-07-31
长江大学学报(自科版)(2018年14期)2018-07-17
中国野生植物资源(2018年6期)2018-01-24
中国果业信息(2015年2期)2015-01-23
东北林业大学学报(2014年11期)2014-03-08
