
天南星科植物叶绿体基因组结构特征及系统发育分析:以天南星等20个物种为例
2024-02-26 03:07许祎珂寇佩雯刘长乐李爽孙晓春黄文静
中国野生植物资源 2024年1期
许祎珂,寇佩雯,刘长乐,李爽,孙晓春,黄文静
(陕西中医药大学 陕西中药资源产业化省部共建协同创新中心/秦药特色资源研究开发国家重点实验室(培育),陕西 咸阳 712083)
叶绿体是绿色植物光合作用的主要细胞器,具有合成淀粉、色素及蛋白质等功能[1]。在大多数被子植物中,叶绿体遗传信息来源于母系,所以其基因组相对稳定且基因较保守[2]。1962年,Ris和Plaut[3]在观察衣藻时发现了叶绿体DNA。1986年,Shinozaki等[4]获得了第一条(烟草)叶绿体基因组序列。同年,Ohyama等[5]获得了地钱的叶绿体基因组的完整序列,借助于测序技术的发展,叶绿体基因组的研究愈来愈多。叶绿体基因组大约由130个基因组成(核糖体RNA、转运RNA、蛋白编码基因),大小在107 ~ 218 kb之间,这些叶绿体基因组均具有典型的四分体结构,包括一个大单拷贝区(Large single copy,LSC)、一个小单拷贝区(Small single copy,SSC)和两个反向重复区(Inverted repeats a,IRa;Inverted repeats b,IRb)[6]。近缘植物的叶绿体基因大小、含量可能会因为IR区的收缩扩张而变化,基因突变、插入、倒置、重排等均在叶绿体基因组中被发现[7-8]。由于叶绿体基因组较小并且相对保守,因此其已经成为进化和比较基因组研究的理想模型,可为揭示植物类群的系统位置和遗传发育关系提供较多的理论依据[9]。
天南星科(Araceae)隶属于单子叶植物纲的泽泻目,该家族有110属3 500余种,主要分布于热带地区及部分温带地区[10]。该科植物大部分具有重要的药用价值,常用的中药材有天南星(Arisaema heterophyllumBlume)、半夏[Pinellia ternata(Thunb.) Breit.]和千年健[Homalomena occulta(Lour.) Schott]等,其中天南星可燥湿化痰、祛风止痉、散结消肿[11];半夏可燥湿化痰、降逆止呕[12];千年健具有祛风除湿,强健筋骨等功效[13]。近些年随着分子生物学和测序技术的发展,关于叶绿体基因组系统发育的研究逐渐增多,如菊科、木犀科和豆科等植物的叶绿体基因组特征及其系统发育关系已被系统报道[14-16]。在天南星科叶绿体基因组研究中,前人分析了白鹤芋[Spathiphyllum lanceifolium(Jacq.) Schott]、绿萝[Epipremnum aureum(Linden et Andre) Bunting]和滴水珠(Pinellia cordataN. E.Brown)的叶绿体基因组以及近缘物种的关系[17-19],但这些研究均侧重于某个物种的叶绿体基因组特征以及部分属的系统发育。目前,数据库中的天南星科植物信息不断增加与更新,亟需开展对该科植物叶绿体基因组全面系统的研究。本研究基于GenBank数据库中相关叶绿体全基因组序列,针对天南星科物种的亲缘关系、遗传多样性和系统发育展开分析,以期为该科植物的遗传学和系统发育进化等研究提供理论依据。
1 数据与方法
1.1 基因组信息
通过NCBI(https://www.ncbi.nlm.nih.gov/)下载GenBank数据库中现有的天南星科20个物种的叶绿体全基因组序列,并以泽泻科慈姑属利川慈姑(Sagittaria lichuanensisJ. K. Chen)和野慈姑(Sagittaria trifoliaL.)的叶绿体全基因组序列为外类群参照,相关物种信息见表1。
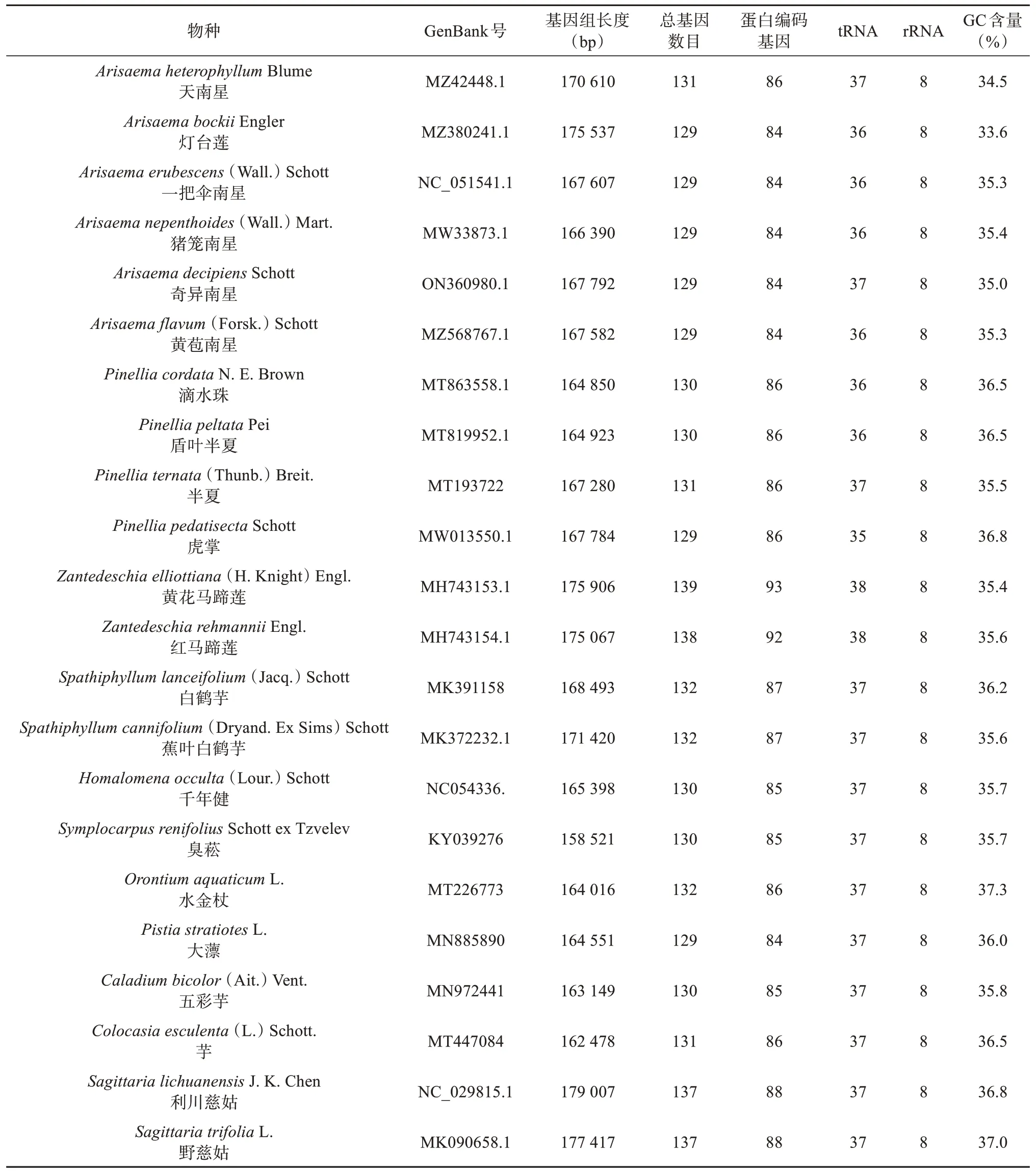
表1 叶绿体全基因组信息Tab.1 Chloroplast genome information
1.2 重复序列和SSR分析
使用在线软件RePuter(https://bibiserv.cebitec.uni-bielefeld.de/reputer)检索串联重复序列,参数设置:最小长度为30 bp,最多碱基错配为3。利用MISA(https://webblast.ipk-gatersleben.de/misa/index.php)检索叶绿体基因组中SSR重复序列,分别将单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸的最小重复设置为8、4、4、3、3和3。
1.3 全基因组分析与共线性分析
以天南星为参照基因组,利用mVISTA(https://genome.lbl.gov/vista/mvista/about.shtml)中的Shuffle-LAGAN模式进行全基因组比较分析,并用Mauve软件进行共线性分析,检测基因组中的重排或倒位现象。
1.4 IR边界收缩、扩张分析
叶绿体基因组中的IR区具有高度保守性,但在不同物种中仍存在微小变化,因此被子植物叶绿体基因组大小的差异与IR边界的收缩和扩张有很大关系。本研究使用IRscope(https://irscope.shinyapps.io/irapp/)软件分析IR边界。
1.5 系统发育分析
以与天南星科同目的泽泻目下泽泻科慈姑属中的利川慈姑和野慈姑作为外类群,利用最大似然法(Maximum likelihood, ML)将天南星科20个物种构建系统发育分析。采用Geneious 8.0.2软件的MAFFT插件进行序列比对,并使用PhyML(http://www.atgc-montpellier.fr/phyml/)软件构建系统发育树,模型为GTR + Gamma,Bootstrap设为1 000。
2 结果与分析
2.1 叶绿体基因组特征
所有的天南星科植物叶绿体基因组皆具有完整的四分体结构(LSC、SSC、IRa和IRb)(表1)。全基因组长度介于158 521(C. esculenta) ~ 175 906 bp(Z. elliottiana)之间,大部分物种的总基因数目介于129 ~ 132个之间。其中,Z. elliottiana和Z. rehmannii的基因数目达到138和139个,基因数目的差异主要是由于蛋白编码基因数目不一致。天南星科植物叶绿体基因组的rRNA基因最为保守,皆为8个基因。碱基GC含量分析表明,不同天南星科物种叶绿体全基因组的GC含量在33.6% ~ 37.3%之间,同属的物种GC含量差异很小,而在不同属之间差异较大。
2.2 重复序列和SSR
采用REPuter对叶绿体基因组中的重复序列进行分析(图1),所有物种均检索出50条重复序列。多数物种正向重复(F)、反向重复(R)、互补重复(C)和回文重复(P)4种类型均检索出,且正向重复和反向重复为主要重复类型;A. heterophyllum、P. cordata、P. peltata、P. pedatisecta、Z. elliottiana、Z. rehmannii和C. bicolor这7个物种中只有正向重复、反向重复、及回文重复;其中Z. elliottiana和Z. rehmannii中只有正向重复和回文重复2种类型。P. cordata、P.peltata、P. pedatisecta、Z. elliottiana和Z. rehmannii这5个物种的50条重复序列长度均大于60 bp,其他物种的重复序列长度以30 ~ 39 bp和40 ~ 49 bp居多(图2)。

图1 天南星科植物叶绿体基因组重复序列种类及数量Fig.1 Species and number of chloroplast genome repeats in Araceae
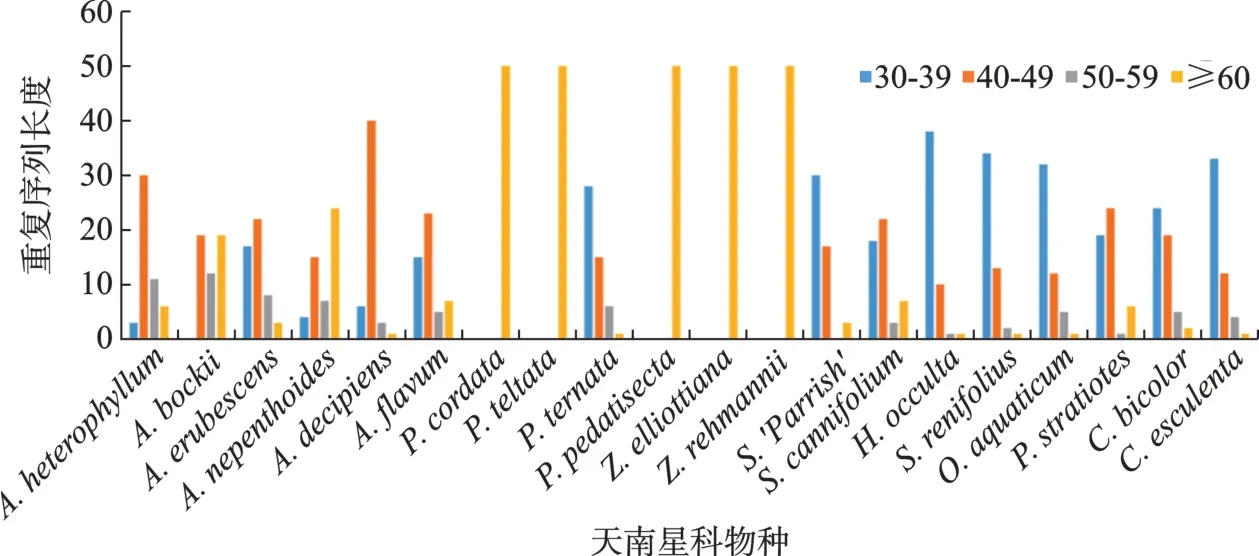
图2 天南星科植物叶绿体基因组重复序列类型及长度分布Fig.2 Type and length distribution of chloroplast genome repeats in Araceae
利用MISA分析天南星科20个物种的SSR。结果表明(图3),叶绿体基因组SSR位点数介于253 ~482个之间,除S. renifolius、C. bicolor和C. esculenta这3个物种没有六碱基重复外,其余物种均有从单碱基重复到六碱基重复6种类型。在所有的SSR重复类型中,单碱基重复和二碱基重复比例较高,六碱基重复最少。单核苷酸重复主要为A/T重复,二核苷酸以AT/AT为主要重复类型,而三、四、五、六核苷酸相对偏少,但是均富含较高含量的A和T碱基。

图3 天南星科植物叶绿体基因组SSR分布特征Fig.3 SSR distribution of chloroplast genomes in Araceae
2.3 全基因组比较与共线性分析
以天南星为参照,我们使用mVISTA软件进一步分析了20种天南星科植物叶绿体全基因组序列的差异性。结果表明(图4),高度可变的区域主要集中在非编码区中,而较为保守的序列多分布在非翻译区,并发现3个高度可变区accD、ycf2和ycf1。Mauve共线性分析可知,天南星科10个属20个物种中,19个物种的叶绿体基因组序列均未检测到大片段基因重排,说明相对保守,只有S. cannifolium中出现基因重排或倒位现象(图5)。

图4 天南星科叶绿体基因组比对Fig.4 Alignment of chloroplast genomes in Araceae

图5 天南星科20种植物叶绿体基因组共线性分析Fig.5 Synteny analysis of chloroplast genomes of 20 species of Araceae
2.4 IR边界
通过比较天南星科物种IR边界分布情况,分析其进化过程中的IR边界收缩扩张现象。天南星科10个属的叶绿体基因组中,基因ndhF都位于SSC区(图6)。除Z. elliottiana的rpl22基因横跨LSC区和IRb区外,其他属的植物rpl22基因都完整位于LSC区;除Z. elliottiana外,其他属植物的rpl2基因均分布在2个IR区;并且大部分植物的rps19基因都完整位于LSC区,唯有Z. elliottiana的rps19基因出现在IR区,这一变异使得rps19基因在Z. elliottiana中为双拷贝。Zantedeschia属的植物与其他属的差异较大,这一结果可能是其IR区过大导致。

图6 天南星科10个属的IR边界比较Fig.6 IR boundary comparison of 10 genera in Araceae
2.5 系统发育分析
基于20个天南星科物种和两个慈姑属物种的叶绿体全基因组序列,构建了天南星科的系统发育关系树(图7)。利川慈姑和野慈姑作为外类群被聚类到天南星科类群外,其余的20个物种中自展值只有一个低于80%,剩余的自展值均为100%,表明结果可信度较高。在进化树中,O.aquaticum和S. renifolius并没有形成单系群,最先从基部聚为一支;其次是S. lanceifolium和S. can-nifolium聚为一支;Z. elliottiana和Z. rehmannii聚为一支,H. occulta作为一个单支与这2个物种聚在一起;P. stratiotes、C. bicolor和C. esculenta这3个物种各为一支;最后是天南星属中6个物种(A. heterophyllum、A. bockii、A. erubescens、A. nepenthoides、A. decipiens和A. flavum)与半夏属中4个物种(P. cordata、P. peltata、P. ternata和P. pedatisecta)各聚为一支,并且天南星属和半夏属关系最近聚为一支。

图7 基于天南星科20个物种的叶绿体全基因组序列构建的系统发育树Fig.7 Phylogenetic trees constructed based on the complete chloroplast genome sequences of 20 Araceae species
3 讨论
3.1 重复序列和SSR分析
在物种进化中,重复序列扮演了重要的角色,因其会影响物种内基因的遗传和变异[20]。本研究发现20个天南星科植物的重复类型主要表现为正向重复和反向重复,少数植物也发现了回文重复和互补重复,特别在黄花马蹄莲和红马蹄莲中仅检索到了正向重复和回文重复。不同植物间重复序列的分布差异可反映植物进化时的遗传速率基因突变速率的差异[21],这表明天南星科植物在突变频率和进化速率上的差异可能与重复序列有关,因此可根据重复序列的差异大小推测物种之间的亲缘关系远近。同时,本研究检索到的重复序列主要分布在非编码区,与大部分被子植物的研究结果相似[22-24]。已有研究表明,在其他物种中大部分重复序列也都大多分布在非编码区,这可能与物种在进化过程中通过保留较少遗传信息来提高遗传效率有关[20]。
SSR分子标记共显性、多态性高,且分布广泛[25],常用于遗传多样性分析、物种鉴定、群体结构研究、遗传图谱构建和QTL定位等研究[26-27]。基于叶绿体基因组独特的母系遗传优势,植物体叶绿体SSR逐步广泛应用于遗传学。本研究发现天南星科物种的叶绿体基因组多以单核苷酸和二核苷酸为主,其他类型的较少。以往研究表明,在所有的SSR类型中,均可发现A和T碱基含量远高于GC含量,这与天南星科植物的叶绿体基因组的结果相似[28]。
3.2 全基因组对比分析
mVISTA全局比对分析可知变异主要集中在非编码区,并发现accD、ycf2和ycf1三个高度可变区,与Ren等[22]研究的Cyperus esculentusLinnaeus和Fu等[29]研究的Dracocephalum heterophyllumBenth.结果相似。这些高度可变区域可用于DNA条形码研究,从而用于天南星科植物中的物种鉴别和遗传图谱分析。
3.3 IR边界分析
IR区的收缩和扩张是植物进化中的普遍现象,同时也是导致叶绿体基因组大小差异的主要原因[30]。本研究中,在天南星科10个属中,rps19基因、rpl12基因、ndhF基因和trnH基因所居位置高度相似,说明天南星科IR区有一定的保守性。Z.rehmannii与其他属相比,差异较大,可能因其IR区扩张导致。在被子植物中,假基因ycf1和rps19基因源于IR区的收缩和扩张[31]。在物种H. occulta中,ycf1基因从IR区开始并整合进入SSC区,导致假基因的形成,功能拷贝基因则出现在IRa/SSC的交界处,而其他物种的ycf1基因主要位于SSC区,并且不会导致假基因的形成。基因rps19通常位于LSC/IR和SSC/IR区,在天南星科植物中,rps19基于主要位于LSC区,这个结果与Liang等[32]研究的壳斗科类似。
3.4 系统发育分析
本研究基于天南星科20个物种的叶绿体全基因组序列构建系统发育树,该科各属的关系得到较好处理,10个属各聚为一支,且节点支持率较高。目前,基于叶绿体基因组测序技术的快速发展,对天南星科中少部分物种的种间关系和属间关系已有研究报导。Zhang等[33]对A. erubescens和Cai等[34]对P. ternata的研究中,天南星属植物和半夏属植物关系最近,与本研究结果一致,并且在本研究中通过mVISTA、Mauve等分析也发现天南星属植物和半夏属植物的叶绿体基因组序列相似度最高。在罗须等[19]的研究中,大薸(P. stratiotes)和芋(C.esculenta)与天南星属和半夏属聚为一支,与本研究中大薸和芋的系统位置相似,说明在天南星科中大薸和芋与天南星属和半夏属植物关系较近。对于天南星科的其他物种,由于目前数据缺少,尚不能进一步探讨。
本研究基于天南星科10个属20个物种的叶绿体全基因组序列,分析了基因组的大小、长度、基因数量、序列变异和基因边界变化等特征,并基于叶绿体全基因组序列构建了系统发育树,且支持率较高,可为天南星科的系统分类和进化关系提供有力的证据。随着高通量测序技术的发展,系统发育关系研究逐渐迈入基因组水平。然而,对于天南星科家族而言,当前该科物种的叶绿体基因组信息数量偏少,因此仍需更进一步开展测序与分析研究,为该科属间关系和种间关系的研究提供科学依据。
猜你喜欢
长春中医药大学学报(2022年7期)2022-11-21
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
中国民族民间医药(2021年9期)2021-06-17
山东畜牧兽医(2020年11期)2020-12-30
Digital Chinese Medicine(2020年3期)2020-11-03
广西林业科学(2016年1期)2016-03-20
癌变·畸变·突变(2014年2期)2014-03-01
食品科学(2013年6期)2013-03-11
