
百山祖自然保护区小型兽类的群落分析
2012-07-30 10:01陈德良吴友贵夏家天鲍毅新
浙江林业科技 2012年3期
陈德良,吴友贵,夏家天,张 旭,鲍毅新*
(1. 浙江凤阳山—百山祖国家级自然保护区百山祖管理处,浙江 庆元 323808;2. 浙江师范大学生态研究所,浙江 金华 321004)
与大、中型兽类相比,小型兽类种类多,数量大,是生态系统中的重要组成部分,在食物链中为第一级消费者,对整个生态系统的物质和能量流动[1]以及维持生态系统结构功能的完整发挥着重要作用[2]。小型兽类的群落结构是生态环境的重要标志,反映了物种的多样性[3]和生态系统的特征,可以作为生态系统演化状况的指标之一[4]。关于小型兽类群落结构及其与环境关系的研究已有不少报道[5~6]。但因自然状况的不同,以及小型兽类具有较高的环境依赖性,各地区的小型兽类群落结构表现出较大差异[7]。目前,百山祖自然保护区小型兽类群落结构研究还未见报道,为此,笔者于2010年7月和11月以百山祖自然保护区为研究区域,研究其小型兽类群落结构及主要影响因子,丰富该地域内小型兽类资料。另外,由于在自然生态系统中,小型兽类群落结构的变化既反映了群落内不同物种之间的关系,同时也反映了群落与其外界环境的联系,因而本研究结果可以为该地区其他物种群落格局研究提供依据,进而为更好的保护栖息地环境提供决策依据。
1 研究区概况和研究方法
1.1 研究区概况
百山祖自然保护区位于浙江省庆元县境内(27° 37′ 45″ ~ 27° 50′ 30″ N,119° 7′ 45″ ~ 119° 19′ 20″ E),总面积108.8 km2,地貌类型为深切割中山,以侵蚀地貌为主,山地陡峭,坡度大多在30°以上,最高峰海拔1856.7 m,是浙江省第二高峰;具有山地立体气候特征,表现为年均温低、降水充沛、相对湿度大、雾日多。年均温12.8℃,年降水量2341.8 mm,相对湿度84%,≥10℃的年积温3653.3℃,极端高温32℃和极端低温-13℃。
保护区植物区系复杂,热带成分与温带成分几乎相等,呈明显过渡性,是华南与华东植物区的连接地带,植被随海拔变化形成有序的垂直谱带,大体可分为3个分布带,海拔1000 m以下为常绿阔叶林和暖性针叶林带,1000 ~ 1500 m为常绿落叶阔叶混交林和温性针叶林带,1500 m以上为山地矮林、灌草丛带[8]。
1.2 研究方法
1.2.1 野外调查 根据保护区的地形和植被特点,可划分为阔叶林、针阔叶混交林、针叶林、灌木林、竹林、农田6种生境[8]。
阔叶林:乔木层主要以甜槠(Castanopsis eyrei)、青冈(Cyclobalanopsis glauca)、木荷(Schima superba)为主;灌木层种类复杂,以隔药柃(Eurya muricata)、麂角杜鹃(Rhododendron latoucheae)、猴头杜鹃(R. simiarum)为主;草本层高5 ~ 60 cm,盖度10% ~ 20 %。
针阔混交林:乔木层主要有柳杉(Cryptomeria fortunei)、福建柏(Fokienia hodginsii)、杉木(Cunninghamia lanceolata)、木荷、甜槠等;灌木层主要有麂角杜鹃、猴头杜鹃、尖叶山茶(Camellia cuspidata)等;草本层高10 ~ 60 cm,盖度20 %~ 40 %。
针叶林:以黄山松(Pinus taiwanensis)占优势,伴生少量木荷、甜槠等;灌木层有映山红(Rhododendron simsii)、麂角杜鹃、马银花(R. ovatum)等;草本层高50 ~ 60 cm,盖度30% ~ 40 %。
灌木林:以杜鹃花科、山茶科、冬青科、山矾科等常绿树种为主;草本层高20 ~ 80 cm,盖度60% ~ 80%。
竹林、农田:生境简单,面积较小,与以上4种生境相邻。农田主要种植蔬菜或粮食作物,与人类居住地有一定距离,基本不从事精细农耕活动,人类活动干扰轻;竹林主要为人工毛竹林。
2010年7月和11月选择五岭坑(海拔约900 m)、万里林(海拔约1100 m)、栗洋村(海拔约1300 m)、百山祖保护站(海拔约1500 m)作为主要调查地点,在其附近的不同生境中以铗日法对啮齿目和食虫目等小型兽类进行调查,同一生境中连续3d依不同路线置中号鼠铗,铗距约5 m,以新鲜花生米为诱饵,对捕获的个体进行种类鉴定、常规测量和剖检。
1.2.2 数据处理 用种类(S)、多样性指数(H')、均匀性指数(E)、优势度指数(D)、相似性指数(I)5个指标进行统计分析对比。探讨各群落的组成和数量与栖息环境之间的相互关系。
群落多样性指数(H')采用Shannan-Weiner公式计算:
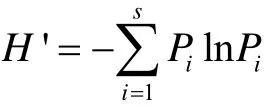
式中,Pi为第i型兽类种个体占群落的比例。
群落均匀度指数采用Pielou(1969)公式计算:
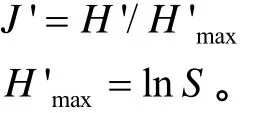
式中,H'max为最大均匀性条件下的物种多样性。
群落优势度指数采用Hurlbert(1978)公式计算:

群落相似性指数(I)采用Whittaker公式计算:
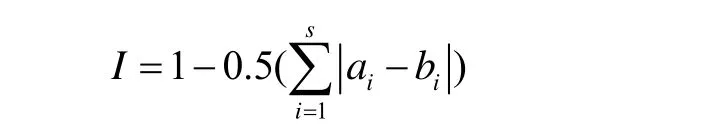
式中,ai和bi分别表示物种i的个体数在群落A和B中所占的比例。
2 研究结果
2.1 群落组成
本次调查共捕获小型兽类101只,隶属于啮齿目和食虫目2目,3科10种。按生境的不同将其划分为6个小型兽类群落,依据各群落物种的个体数量分布来确定群落的优势种(表1),并结合夏武平的群落命名方法[9],命名如下:
2.1.1 群落 A(针阔混交林) 社鼠+刺毛鼠群落。其捕获率在所有群落中最低,有啮齿目动物 2种,社鼠(Niviventer confucianus)为绝对优势种,占75%。从动物地理区划看[10],刺毛鼠(针毛鼠,N. fulvescens)属南中国型,社鼠为广布种。
2.1.2 群落B(针叶林) 社鼠+刺毛鼠+中华姬鼠群落。共有6种小型兽类,社鼠、刺毛鼠和中华姬鼠(Apodemus draco)分别占38.5%、23.1%和23.1%,小家鼠(Mus musculus)、灰麝鼩(Crocidura attenuate)和山东小麝鼩(小麝鼩C. shantungensis)数量稀少。本群落中属南中国型的有刺毛鼠、中华姬鼠和灰麝鼩,属北方型的有小家鼠,社鼠和山东小麝鼩为广布种。
2.2.3 群落C(阔叶林) 刺毛鼠群落。本群落小型兽类物种单纯,仅此一种,刺毛鼠为南中国型。
2.1.4 群落D(灌木林) 黑线姬鼠+社鼠+刺毛鼠群落。小型兽类物种最丰富,为8种。黑线姬鼠(Apodemus agrarius)占30.0%,社鼠为26.7%,刺毛鼠为16.7%,中华姬鼠、黄毛鼠(Rattus losea)、山东小麝鼩均为6.7%,黑腹绒鼠(Eothenomys melanogaster)和灰麝鼩数量最少。本群落中属南中国型的有刺毛鼠、中华姬鼠、黑腹绒鼠和灰麝鼩,属北方型的有黑线姬鼠,属东洋型的有黄毛鼠,社鼠和山东小麝鼩为广布种。
2.1.5 群落E(竹林) 社鼠+小家鼠+刺毛鼠群落。共有6种小型兽类,社鼠、小家鼠和刺毛鼠分别占42.9%、25.0%和17.8%,黑线姬鼠占7.1%,黄毛鼠和山东小麝鼩均为3.6%。群落中刺毛鼠为南中国型,小家鼠和黑线姬鼠为北方型,黄毛鼠为东洋型,社鼠和山东小麝鼩为广布种。
2.1.6 群落F(农田) 社鼠+小家鼠群落。捕获率最高,达2.12%。共有5种小型兽类,优势种明显,社鼠和小家鼠分别占45.5%和27.3%,黑线姬鼠、青毛硕鼠(青毛鼠Berylmys bowersi)和黑腹绒鼠数量较少。群落中属北方型的有小家鼠和黑线姬鼠,属南中国型的有黑腹绒鼠,属东洋型的有青毛硕鼠,社鼠为广布种。
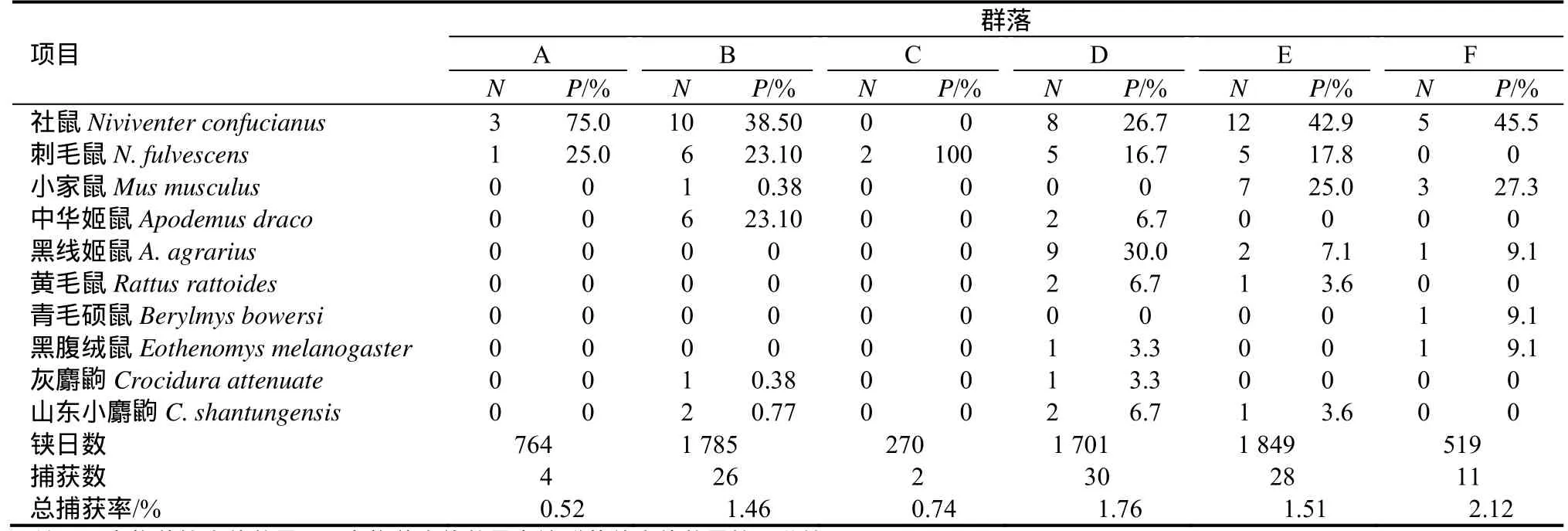
表1 各动物群落的物种组成及数量Table 1 Composition and quantity of the small mammal communities
2.2 群落多样性
物种多样性指数是群落多样性高低的定量指标,在一定程度上反映了群落的复杂程度[11~13]。根据不同群落所捕获动物的种类和个体数量,分别计算各群落的多样性指数、均匀度指数和优势度指数等,结果见表2。

表2 不同小型兽类群落多样性、均匀度指数及优势度指数Table 2 Diversity, evenness and dominance index of each small mammal community
群落多样性指数由高到低依次为:群落D(灌木林) > 群落E(竹林) > 群落F(农田) > 群落B(针叶林) > 群落A(针阔混交林) > 群落C(阔叶林)。均匀度指数除了群落B(针叶林)较低,其他5个群落均匀度差异不大,均在0.800 ~ 0.850。优势度指数群落A与群落C较高,其次依次为群落F、群落E、群落B、群落D。
2.3 群落相似性
不同小型兽类群落的相似性指数见表3。
由表3可见,群落B和D的相似性指数最大(I= 0.881),其次是群落E和群落F(I= 0.750),群落C与其他5个群落的相似性均普遍较低。
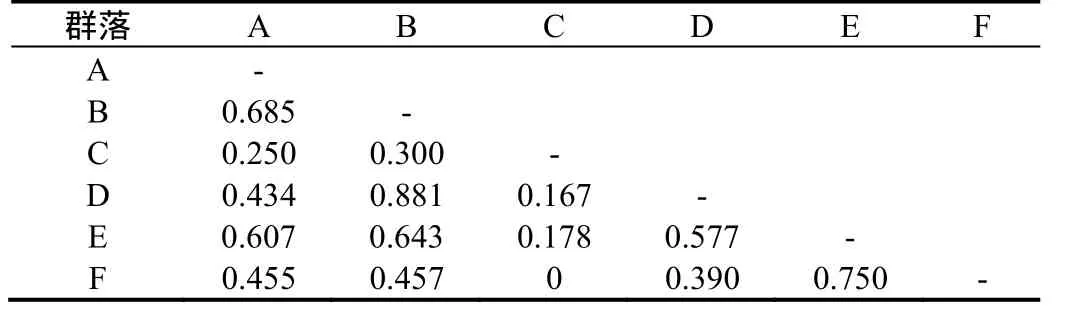
表3 不同小型兽类群落的相似性指数Table 3 Similarity index of different small mammal communities in the tested area
3 讨论
小型兽类依赖环境为其提供食物和隐蔽条件等生存的基本要素,所以其群落的物种组成与所处的生境之间存在密切联系[14]。
草本植物的作用,一方面是为小型兽类提供食物,另一方面其覆盖度的高低将在很大程度上决定小型兽类的被捕食风险。灌木林草本植物最为茂盛,加上地形的多样化,生境的食物资源与隐蔽条件最好,为更多物种的共存创造了良好的微环境,为多种小型兽类所利用,这与鲍毅新和诸葛阳[15]对金华北山啮齿类的群落研究相同,因此群落 D(灌木林)的多样性指数最高。阔叶林林冠郁闭度高,林下光线较弱,必然会影响到草本植物的生长,而针叶林和针阔混交林的郁闭度相对较低,草本植物的生长相对比较茂盛,因而对小型兽类的生存比较有利[16]。因此,群落B(针叶林)和群落A(针阔混交林)的多样性指数要大于群落C(阔叶林)。
一般情况下,农田生境相对于森林、灌丛等生境的植物群落结构单一,复杂性降低,所以多样性降低[17]。但是百山祖自然保护区内,单个农田和竹林生境面积很小且均是林缘农田和竹林。Bider[18]和Sekgororoane等[19]的研究表明森林与农田交界处,小型兽类的多样性较相邻的任一栖息地都要高。边缘效应的作用可能就是导致本研究中农田和竹林生境的群落多样性指数要高于阔叶林、针叶林以及针阔混交林的主要原因。
优势度指数反映了各物种种群数量变化状况,指数越大,说明群落内物种数量的分布越不均匀,优势种的地位越突出[20]。由于群落A和群落C(阔叶林)捕获物种数均没有超过2种,因此导致优势度较大,无太大意义。其他4种生境中,群落F(农田)的优势度最大(D= 0.306),从表1也可看出,该群落中社鼠的分布数量最多。从均匀性指数和优势度指数的定义及计算公式来看,二者之间存在着负相关,即均匀性指数越小的群落,优势度越大。但本研究中群落A的优势度指数最大(D= 0.625),但其均匀性指数却并不是最小的(E= 0.811),出现这种结果的原因可能与调查时放置鼠铗的强度不同有关[21]。
群落B和群落D的相似性指数最大(I= 0.881),这也表明生境中草本植物越丰富,其群落也越相似;其次是群落E(竹林)和群落F(I= 0.750),可能是这两种生境均受边缘效应的影响而导致群落结构方面相接近。群落C与其他5个群落的相似性均普遍较低,显然这是由于群落C仅捕获到1个物种所导致的。
不同生境中植被类型不仅影响小型兽类的组成和分布,而且也与啮齿动物的密度有关。在灌木林、农田生境中小型兽类密度较高,而针叶林、针阔混交林、阔叶林等森林生境中小型兽类密度较低。显然,在结构复杂的生态系统中,小型兽类作为重要的组成成分之一必然要受到诸多因子的制约,因此其数量会较低。
百山祖自然保护区与相邻地区的天目山自然保护区[22]、金华北山[15]的小型兽类组成有明显的差异。这可能跟研究地点的海拔不同有关,本研究的海拔主要在1000 ~ 1500 m,天目山的研究在400 ~ 1400 m,金华北山的研究在500 ~ 1150 m,海拔高度的不同造成的气候的差异,可能是导致物种组成差异的主要原因。然而社鼠在这3个地点均是优势种,有研究表明其组成百分比一般在40%以上,有的高达100%[23],这可能跟其作为广布种适应能力较强有关。
[1]郑雄,李波,蒋光权,等. 四川九顶山自然保护区的小型兽类群落结构[J]. 四川林业科技,2009,30(5):69-85.
[2]王淯,王小明,胡锦矗,等. 唐家河自然保护区小型兽类群落结构[J]. 兽类学报,2003,23(1):39-44.
[3]刘清君,胡锦矗,吴攀文. 南充高坪区嘉陵江畔鼠型小兽多样性调查[J]. 西华师范大学学报(自然科学版),2008,29(4):413-416.
[4]涂云飞. 四川夹金山小型兽类区系及群落多样性研究[D]. 雅安:四川农业大学,2010.
[5]阿不都热合曼·吐尔逊,余亮,艾尼瓦尔·吐米尔,等. 新疆北部干旱地区鼠类群落空间结构差异的比较研究[J]. 新疆大学学报(自然科学版),2008,25(2):211-218.
[6]赵庆洋,鲍毅新,孙波,等. 千岛湖岛屿小型兽类群落分布格局及其影响因素[J]. 动物学研究,2009,30(6):671-678.
[7]傅必谦,陈卫,高武. 百花山鼠类群落结构及其与环境的关系[J]. 北京师范学院学报(自然科学版),1992,13(4):69-77.
[8]余久华,姚丰平,陈小荣,等. 百山祖自然保护区主要植被类型概述[J]. 热带亚热带植物学报,2003,11(2):93-98.
[9]夏武平. 谈谈草原啮齿动物的一些生态学问题[J]. 动物学杂志,1964,6(6):299-302.
[10]张荣祖. 试论中国陆栖脊椎动物地理特征—以哺乳动物为主[J]. 地理学报,1978,33(2):85-101.
[11]Hodkinson I D, Hodkinson E. Pondering the imponderable: a probability based approach to estimating insect diversity from repeat faunal samples [J]. Entomology,1993,18(1):91-92.
[12]Scheiner S M. Measuring pattern diversity [J]. Ecology,1992,73(5):1860-1867.
[13]Wilson C C, Hebert P D N. The maintenance of taxon diversity in an asexual assemblage: an experimental analysis [J]. Ecology, 1992, 73(4):1402-1422.
[14]肖治术,王玉山,张知彬,等. 都江堰地区小型哺乳动物群落与生境类型关系的初步研究[J]. 生物多样性,2002,10(2):163-169.
[15]鲍毅新,诸葛阳. 金华北山啮齿类的生态研究[J]. 兽类学报,1987,7(4):266-274.
[16]杨春文. 东北主要林区森林五种啮齿动物共存机制研究[D]. 哈尔滨:东北林业大学,2007
[17]张美文,王凯荣,王勇,等. 洞庭湖区鼠类群落的物种多样性分析[J]. 生态学报,2003,23(11):2260-2270.
[18]Bider A. Animal activity in uncontrolled terrestrial communities as determined by a sand transect technique[J]. Ecol Monog, 1963(38):269-308.
[19]Sekgorroane G B, Dilworth T G. Relative abundance, richness and diversity of small mammals at induced forest edges[J]. Can J Zool,1995,28(3):1432-1437.
[20]张云智,龚正达,冯锡光,等. 云南白岭草鼠形小兽群落结构及垂直分布[J]. 动物学杂志,2002,37(2):63-66.
[21]王艳妮,周材权,张君,等. 唐家河国家级自然保护区夏季啮齿动物的群落分析[J]. 西华师范大学学报(自然科学版),2005,26(3):247-251.
[22]鲍毅新,诸葛阳. 天目山自然保护区啮齿类的研究[J]. 兽类学报,1984,4(3):197-205.
[23]鲍毅新. 社鼠的研究概要[J]. 浙江师范大学学报(自然科学版),1993,16(2):50-54.
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
林业与环境科学(2021年1期)2021-04-12
落叶果树(2021年6期)2021-02-12
河南科学(2020年3期)2020-06-02
安徽农学通报(2018年8期)2018-05-30
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
