
秸秆纤维素降解真菌QSH3-3的筛选及其特性研究
2012-08-31 02:15刘爽范丙全
植物营养与肥料学报 2012年1期
刘爽,范丙全
(中国农业科学院农业资源与农业区划研究所,北京100081)
农作物秸秆是我国农业上最丰富的有机质资源,同时也是我国年产量最多的农业副产品,据不完全统计,我国每年农作物秸秆产出量达到7~8亿吨。农作物秸秆结构复杂,短期内不易自然腐解,利用效率低,尤其在种植业为主的地区,作物秸秆已经成为生产中的障碍。加快秸秆的腐熟,解决农业主产区的秸秆问题成为国内外科学家的研究热点,而秸秆纤维素降解菌在加快秸秆腐解方面发挥着重要作用[1]。
自然界中能分解纤维素的有细菌、放线菌和真菌。文献报道的细菌主要有纤维粘菌(Cytophaga)和纤维杆菌(Cellulomonas)等[2],细菌所产纤维素酶酶系单一,主要是内切酶,多为胞内酶,产量也不高,且多数结合在细胞膜上,菌体细胞需吸附在纤维素上才能起作用,使用很不方便,酶的分离提取也较困难[3]。具有降解纤维素能力的放线菌主要有纤维放线菌(Acidothermus cellulolyticus)、诺卡氏菌属(Ncardia)和链霉菌属(Streptomyces),但放线菌产量极低,所以研究很少[4-6]。真菌由于具有较高的胞外酶活性且其酶种类比较全面使其有很强的纤维素降解能力,因此对其的研究报道比较多。其中主要以 木 霉 属 (Trichoderma)[7-9]、曲 霉 属 (Aspergillus)[10-11]、青霉(Penicillium)[12-13]为主,木霉属所产纤维素酶的组分比较齐全,但存在β-葡萄糖苷酶活性较低,产酶速度慢,作用pH范围窄,耐低温性差等问题[14];黑曲霉是优良的产β-葡萄糖苷酶菌株,但酶系组分较为单一[15];青霉属(Penicillium)中的一些种类不仅能分泌组成齐全、酶活较高的纤维素降解酶系,而且具有易培养、生长快的优势[16],但由于之前研究较少,对其所产纤维素酶系需要更全面、深入的认识。因而,目前研究的纤维素降解真菌菌株存在产酶组分系不全、活性不稳定、产酶速度较慢、作用pH范围窄、特别是产酶温度要求高等问题,限制了其在田间,特别是北方低温地区对秸秆的促熟作用,获取高效降解秸秆纤维素的微生物菌株是解决秸秆还田的关键因素。本文拟从低温高腐殖质环境中筛选在常温、低温下具有产纤维素酶能力的青霉属真菌菌株,并对其产酶特性和酶学性质进行研究,以期为常温、低温条件下农作物秸秆的还田利用提供菌种资源和理论依据。
1 材料和方法
1.1 实验材料
1.1.1 土壤样品的采集 土壤、腐殖质落叶层样品取自小兴安岭原始林区及黑龙江农垦集团多年秸秆还田的土壤。
1.1.2 培养基[17](1)赫奇逊氏(Huchinson)无机盐培养基:KH2PO41.0 g,NaCl 0.1 g,MgSO4·7H2O 0.3 g,NaNO32.5 g,FeCl30.01 g,CaCl20.1 g,水1000 mL,pH 7.2左右。(2)滤纸培养基:赫奇逊氏无机盐培养基中添加圆形滤纸片。(3)羧甲基纤维素钠(CMc-Na)平板培养基:CMc-Na 10.0 g,200 g去皮马铃薯汁1000 mL,琼脂18 g,加蒸馏水至pH 7.2左右。(4)种子培养基:采用马铃薯–蔗糖琼脂(PDA)培养基。(5)液体发酵培养基:KH2PO41.0 g,NaCl 0.1 g,MgSO4·7H2O 0.3 g,NaNO32.5 g,FeCl30.01 g,CaCl20.1 g,秸秆木质纤维素50 g或0.5% 的其他碳源,水1000 mL,pH为7.2~7.4。
上述培养基均在121℃,1.01 ×105Pa条件下灭菌30 min备用。
1.1.3 秸秆纤维素的制备 将玉米秸秆剪成2~3 cm小段,烘干后粉碎至0.25 mm,加入2 mol/L的氢氧化钠溶液,在室温浸泡12 h,水洗至pH为中性,烘干备用。
1.1.4 主要试剂及仪器 试剂:DNS试剂,柠檬酸缓冲液,燕麦木聚糖(Sigma公司),羧甲基纤维素钠(国药集团化学试剂有限公司);新华定性滤纸。仪器:分光光度计、分析天平、恒温摇床、恒温水浴箱、离心机。
1.2 测定项目与方法
1.2.1 秸秆降解菌的筛选 直接接种腐殖土粒及腐殖落叶于赫奇逊氏培养基上(培养基凝固后在琼脂表面放一张灭菌无淀粉滤纸)[8]富集培养,15℃培养20 d后,挑取生长旺盛且使得滤纸变薄或透明的真菌菌落进行纯化和保种。再将纯化过的菌株分别点接到CMc-Na初筛培养基上培养3 d,采用刚果红染色法染色,挑取有透明圈的菌株接入到液体滤纸培养基中进行复筛,选取24 h内可以将滤纸消解的菌株进行液态发酵产酶试验,选取产酶量高的菌株。
1.2.2 秸秆降解菌的鉴定形态鉴定:将菌株接种到PDA固体培养基上,25℃培养7 d后,观察菌落形态并镜检其产孢结构和菌丝形态,根据《真菌鉴定手册》[18]和《中国真菌志》[19]进行鉴定。分子生物学鉴定:采用rDNA ITS基因序列分析[20]。取培养24 h 菌丝体,提取 DNA[21-22]后,扩增引物 ITS1(5’-GGAAGTAAAAGTCGTAACAAGG -3’)和 ITS4(5’-TCCTCCGCTTATTGATATGC -3’),进行 18S rDNA的扩增,PCR扩增产物经上海生工生物技术有限公司测序后,将获得的序列与GenBank中已知真菌的18S rDNA序列进行比对。并利用MEGA4.1进行系统发育树的构建。PCR反应体系为50 μL,反应条件为 94℃ 5 min,94℃ 30 s,57℃ 50 s,72℃50 s,循环30 次;72℃ 10 min。
1.2.3 秸秆降解菌产酶特性分析 将筛选到的菌种在PDA上进行纯培养7 d,用无菌水将孢子洗下,制成孢子悬液(1×107/mL)。然后按1%的接种量将孢子悬液加入到100 mL(300 mL的三角瓶)pH为7的赫奇逊氏液体培养基中,碳源为0.5%碱处理的玉米秸秆粉,25℃、170 r/min培养,分别在第1、2、3、4、5、6、7 和 8 d 取样测定粗酶液中的内切酶活力(CMCase)、滤纸酶活力(FPase)、和木聚糖酶活力(Xylanase)。根据菌株产酶时间变化趋势,接种1%的孢子悬液到100 mL(300 mL的三角瓶)pH为7的赫奇逊氏液体培养基中,研究不同碳源对产酶的影响时,其中碳源为不同结构的纤维素[0.5%羧甲基纤维素(CMC)、微晶纤维素(MCC)、玉米秸秆粉(CS)以及碱处理的玉米秸秆粉(APCS)];研究不同氮源对产酶的影响时,以碱处理的玉米秸秆粉为碳源,以0.2% 蛋白胨,(NH4)2SO4,(NH2)2CO和NH4NO3替换NaNO3作为单一氮源;在最佳碳源、氮源条件下,研究不同起始 pH(3.0、4.0、5.0、6.0、7.0、8.0、9.0 和10.0)、接种量(1%、3%、5%、7%和9%)和温度(10℃、15℃、20℃、25℃、30℃、35℃、40℃、45℃)对产酶的影响,每个处理设3个重复,25℃、170 r/min培养4 d,离心,取上清液测定内切酶、滤纸酶和木聚糖酶。
1.2.4 酶学稳定性分析 接种1%的孢子悬液到100 mL(300 mL的三角瓶)pH为7的赫奇逊氏液体培养基中,以碱处理的玉米秸秆粉为碳源,25℃、170 r/min培养4 d,离心,取上清获得粗酶液。将0.5 mL 粗酶液加入到 pH 值为 3.0、4.0、5.0、6.0、7.0、8.0、9.0和10.0的1.5 mL缓冲液中,45℃保温1 h,测定滤纸酶、内切酶和木聚糖酶。将0.5 mL粗酶液加入到pH值5.0的1.5 mL缓冲液中,在40℃、45℃、50℃、55℃、60℃、65℃ 各温度下保温 1 h后,测定内切酶、滤纸酶和木聚糖酶。
1.2.5 纤维素酶、木聚糖酶活力的测定 发酵液离心10 min(4000 r/min),取上清液作为粗酶液。用DNS法测定粗酶液中内切酶、滤纸酶和木聚糖酶活力[23-26]。内切酶和滤纸酶单位定义为每mL每min产生1 μg的葡萄糖所需要的酶量定义为一个酶活单位(U)[27],木聚糖酶单位定义为每mL每min产生1 μg的木糖所需要的酶量定义为一个酶活单位(U)[28]。
试验数据采用Microsoft Excel(2003)和SPSS V17.0统计软件进行统计分析,LSD法检验差异显著性。
2 结果与分析
2.1 纤维素降解低温菌的筛选
从东北地区的低温土壤样品中,经富集、初筛得到12株真菌,能够在15℃条件下以滤纸为唯一碳源的培养基上生长良好,而且将滤纸完全降解。再将菌株接种到以CMC为唯一碳源加入刚果红的无机盐培养基上,以透明圈直径大小为标准进行复筛,获得效果最好菌株QSH3-3。
2.2 产纤维素酶菌株的鉴定
2.2.1 形态特征显示 QSH3-3菌株所形成菌落在PDA平板上先以白色絮状轮生,待长出青绿色孢子后,孢子会向四面散射,在平板上形成许多菌落。随培养时间延长,孢子变成暗绿色。菌落平坦,质地绒状。菌落背面呈黄绿色。挑取孢子与菌丝体在显微镜下观察,分生孢子梗帚状分支,最末级的瓶状小枝上生出成串的分生孢子。孢子透明,呈卵圆形,光滑,约4 μm ×5 μm。
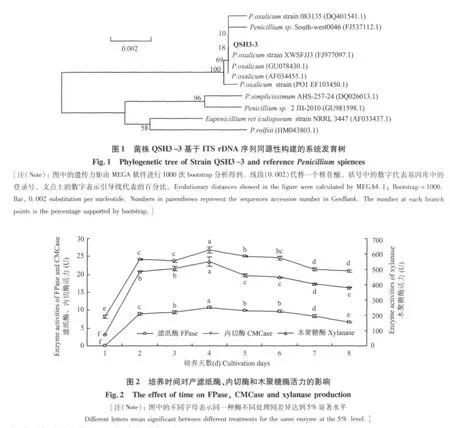
2.2.2 ITS序列分析 提取菌株QSH3-3 DNA后,利用ITS通用引物进行PCR扩增,将扩增并验证得到的片段测序,然后经过DNASTAR软件校正拼接,获得572 bp的核苷酸序列。将获得的序列在Gen-Bank上进行同源性比较,发现该序列与青霉菌的序列同源性>98%,利用MEGA4.1的Neighbor-Joining进行系统发育树的构建,结果如图1,遗传距离显示菌株QSH3-3与青霉遗传距离最近。结合形态观察,初步确定菌株QSH3-3属于青霉属中的草酸青霉(Penicillium oxalicum)。
2.3 不同条件对菌株QSH3-3产酶效果的影响
2.3.1 培养时间对产酶的影响 将菌株QSH3-3按1%接种量接入到液体培养基中发酵培养(170 r/min,25℃),每隔1 d取样测定滤纸酶、内切酶、木聚糖酶。结果显示,滤纸酶、内切酶、木聚糖酶在培养2 d时酶活力分别达到9 U,24 U,487 U,到4 d时均达到最高,分别为11 U,27 U,550 U,之后酶活缓慢下降。菌株在培养2~6 d,残余酶活力达80%以上(图2)。在之后考察各种因子对产酶影响效果时,均测定发酵培养4 d后粗酶液的酶活力。
2.3.2 不同碳源对产酶的影响 以不同结构的纤维素[0.5% 羧甲基纤维素(CMC)、微晶纤维素(MCC)、玉米秸秆粉(CS)以及碱处理的玉米秸秆粉(APCS)]为底物,研究碳源对菌株QSH3-3产酶的影响(表1)。碱处理玉米秸秆粉处理的产酶效果最佳,滤纸酶、内切酶、木聚糖酶分别达到11 U、27 U、550 U,其次为天然的玉米秸秆粉,羧甲基纤维素和微晶纤维素产酶效果较差。以碱处理的玉米秸秆粉比玉米秸秆粉为底物的滤纸酶、内切酶和木聚糖酶分别提高57.1%、28.6%和62.7%;比以MCC为底物的分别提高36.3%、46.1%和136.1%;比以CMC为底物的分别提高了22.2%、92.9%和957.7%。
2.3.3 不同氮源对产酶的影响 发酵液体培养基中分别添加0.2%的蛋白胨,硫酸铵,脲,硝酸铵作为单一氮源,碳源均为碱处理的玉米秸秆粉,接种量为1%,发酵培养(170 r/min,25℃),4 d后测定滤纸酶、内切酶、木聚糖酶。结果显示,蛋白胨为氮源时,滤纸酶、内切酶、木聚糖酶酶活力最大,分别达到11.2 U、26.6 U和582.8 U(表2);硫酸铵作为氮源时,滤纸酶、内切酶、木聚糖酶三种酶活性与蛋白胨作为氮源的处理没有明显差异,但与脲作氮源相比,滤纸酶、内切酶、木聚糖酶分别高了 28.6%、62.3%、178.7%,差异显著;与硝酸钠作氮源相比分别高了约3.8%、1.2%和4.1%。考虑成本因素,本研究选择硫酸铵为草酸青霉QSH3-3产酶培养的氮源。

表1 同碳源对产滤纸酶、内切酶和木聚糖酶活力的影响(U)Table 1 The effect of carbon sources on FPase,CMCase and xylanase production

表2 不同氮源对产滤纸酶、内切酶和木聚糖酶活力的影响(U)Table 2 The effect of nitrogen sources on FPase,CMCase and xylanase production
2.3.4 不同起始pH对产酶的影响 将菌株QSH3-3按1% 接种量接入到4~9的不同起始pH的液体培养基中发酵培养(170 r/min,25℃),结果表明,初始pH值在4~9的液态发酵条件下,菌株都能很好的生长,而且滤纸酶、内切酶和木聚糖酶三种酶活力表现相同规律,随着起始pH值升高酶活力升高,到pH值为7.0时,达到最大,其后逐渐下降(图3)。在初始pH值为7的条件下,滤纸酶、内切酶、木聚糖酶分别约为11 U、28 U、576 U,比初始pH值为4时分别高约30.2%、24.0%、22.8%;比初始pH值为9时分别高约19.0%、18.1%、18.2%。初始pH值在4~9范围内,三种酶活力均可达到最适起始pH值为7时酶活力的70%以上。

图3 起始pH对产滤纸酶、内切酶和木聚糖酶活力的影响Fig.3 The effect of initial pH on FPase,CMCase and xylanase production
2.3.5 不同接种量对产酶的影响 将菌株按1%、3%、5%、7%和9% 的接种量(107/mL菌悬液)分别接入到液体培养基中发酵培养(pH 7,170 r/min,25℃)。结果显示,滤纸酶、内切酶、木聚糖酶在接种量分别为5%、5% ~7%和5%时,酶活达到最大(表 3),分别达到11.8U、28.2U、593.0 U,与接种量为1%时相比,滤纸酶、内切酶、木聚糖酶分别高出9.3%、8.0%和4.3%,比接种量为9%时分别高出37.2%、16.1%和4.9%。接种量小于5%或者大于5%,菌株所产三种酶的酶活均不能达到最高值。只有接种量在一定范围内才能保证菌株的最大产酶量。本试验以5%接种量最佳,在此条件下,滤纸酶、内切酶、木聚糖酶酶活力达到最大。
2.3.6 不同培养温度对产酶的影响 温度是影响产纤维素酶菌株活力的重要因素之一,培养温度对产酶有较大的影响。将菌株QSH3-3按5%接种量接入到液体培养基中,于 10℃、15℃、20℃、25℃、30℃、35℃、40℃、45℃摇瓶发酵培养,4 d后测定滤纸酶、内切酶、木聚糖酶。结果表明,产酶温度在10℃~30℃范围内,酶活力随温度升高酶活逐渐增大,产酶温度在30℃ ~45℃范围内,酶活力随温度升高迅速下降,当产酶温度为30℃时对菌株产酶最为有利,滤纸酶、内切酶、木聚糖酶分别达到12 U、30 U、605 U,且该菌株最大优势是在15℃条件下滤纸酶、内切酶、木聚糖酶即可达到最大酶活力的73.9%、78.9%和80.6%(图4)。该研究结果显示了该菌株在低温条件下对秸秆纤维素类物质的降解潜力。

表3 接种量对滤纸酶、内切酶和木聚糖酶产酶活力的影响(U)Table 3 The effect of inoculation volume on FPase,CMCase and xylanase production
2.4 酶学稳定性
2.4.1 酶的pH稳定性 将菌株QSH3-3在最佳产酶条件下的粗酶液分别置于pH值在3~10范围内的缓冲液中,室温保持1 h后,测定内切酶、滤纸酶和木聚糖酶。结果显示,在pH 4~9范围内,酶活力保持稳定,内切酶残余酶活力达到70%以上;滤纸酶和木聚糖酶残余酶活力达到85%以上(图5)。该菌株所产的内切酶、滤纸酶和木聚糖酶具有较强的pH稳定性。

2.4.2 酶的热稳定性将菌株QSH3-3在最佳产酶条件下的粗酶液分别在 40℃、45℃、50℃、55℃、60℃、65℃的不同温度下保温1 h后,测定内切酶、滤纸酶和木聚糖酶。由图6可以看出,三种酶的酶活力规律一致,随着温度的升高其活性逐渐降低。内切酶、滤纸酶和木聚糖酶在65℃时,分别只有45℃条件下的40%、53%和13%(图6)。内切酶和滤纸酶在40~50℃之间酶活保持稳定,在50℃以上,酶活力迅速下降,木聚糖酶在40~45℃之间酶活保持稳定,45℃以上,酶活力迅速下降。该菌株所产内切酶、滤纸酶和木聚糖酶的耐热性较差。

图6 不同温度条件下滤纸酶、内切酶和木聚糖酶活力的稳定性Fig.6 Thermal stability of FPase,CMCase and xylanase activities
3 讨论
秸秆成分复杂,在高温堆腐条件下,较易腐解,而秸秆的应用难题在于农田大规模秸秆的降解,其腐解温度低,需要筛选常温或低温条件下能够产生高活性的纤维素、半纤维素酶菌株加快秸秆腐熟,已报道的产纤维素酶菌株对其产酶适宜温度的研究大多都在25℃以上,且具有较高产酶能力的温度范围一般都在 25 ~35℃范围内[29-30],低温条件下产酶能力相当较弱,不利于在田间温度较低条件下发挥作用。王希国等[29]研究了不同培养温度对梅林青霉下的产纤维素酶的影响,在24℃时,残余纤维素酶活力约为40% ~60%,而本实验筛得的菌株QSH3-3在15℃时即可产生的酶活力较高,残余酶活力可以达到70%以上,这对在地温较低的田间秸秆原位还田具有重要意义。另外,田间环境中酸碱差异较大,南方土壤一般呈偏酸性,北方土壤一般呈偏碱性,菌株不能保持在中性环境下进行生长,而目前筛得的菌株产酶的最适pH条件一般均为偏酸性,王仪明等[30]研究表明绿色木霉产纤维素酶的适宜pH在5~6.5范围内,培养基起始pH 7.5时纤维素酶的产量大大下降。史玉英等[31]分离的纤维素分解菌群,对滤纸、膨化稻草等自然纤维素表现出较强的分解能力,但培养4 d后由于pH失调而反应终止。菌株产酶的pH范围窄限制了其在田间的推广应用。所筛菌株QSH3-3在pH 4~9范围内,均可保持较强的产纤维素酶的能力,且该菌株所产纤维素酶、木聚糖酶均具有较强的pH耐受力。因而菌株QSH3-3该菌株在田间温差大、土壤偏碱性等复杂条件下对秸秆纤维素类物质的降解具有较高的应用潜力。
4 结论
本研究在15℃下筛选到1株产纤维素酶真菌QSH3-3,经鉴定为草酸青霉Penicillium oxalicum。该菌株以碱处理过的玉米秸秆粉为碳源,硫酸铵为氮源,起始pH为7,接种量5%,30℃下培养4 d,滤纸酶、内切酶、木聚糖酶分别达到约12 U、30 U、605 U,其中木聚糖酶是已报道的最大酶活为216 U的长梗木霉E3菌株[32]的2.8倍,最大酶活387 U赭绿青霉Sp1[33]的1.56倍,最大酶活136.9 U的粗糙脉孢菌[34]的4.4倍,滤纸酶、内切酶与目前报道的酶活力一致。
[1]赵明文,史玉英,李玉祥,娄无忌.纤维分解菌群对水稻秸秆田间腐熟效果的研究[J].江苏农业科学,2000,1:51-53.
Zhao M W,Shi Y Y,Li Y X,Lou W J.Effect of cellulose de-composing bacteria population on ecomposition of rice straw in fields[J].Jiangsu Agric.Sci.,2000,1:51 -53.
[2]刘娣.秸秆纤维素高效降解真菌的筛选、鉴定及其纤维素酶基因克隆[D].北京:中国农业科学院硕士学位论文,2008.
Liu D.Screening,identification and cellulase gene cloning of high effective straw cellulose-degrading fungus strains[D].Beijing:MS thesis,Chinese Academy of Agricultural Sciences,2008.
[3]陈红歌,张东升,刘亮伟.纤维素酶菌种选育研究进展[J].河南农业科学,2008(8):5-7.
Chen H G,Zhang D S,Liu L W.Recent advances of breeding strains of cellulose[J].J.Henan Agric.Sci.,2008(8):5 -7.
[4]Sakon J,Adney W S,Himmel M Eet al.Crystal structure of thermostable family 5 endocellulase E1 fromAcidothermus cellulolyticusin complex with cellotetraose[J].Biochem.J.,1996,35(33):10648-10660.
[5]刘东波,高培基.纤维素诺卡氏菌的一个新菌株[J].微生物学报,1990,30(1):70-72.
Liu D B,Gao P J.A new stain ofNcardia.Cellulolyticus[J].Acta Microbiol.Sin.,1990,30(1):70 -72.
[6]Li X.Streptomyces cellulolyticus sp.nov.,a new cellulolytic member of the genus Streptomyces[J].Int.J.Syst.Bacteriol.,1997,47(2):443-445.
[7]李振红,陆贻通.高效纤维素降解菌的筛选[J].环境污染与防治,2003,25(3):133-136.
Li Z H,Lu Y T.Screening on the cellulose-decomposing microorganisms[J].Environ.Pull.Control,2003,25(3):133 -136.
[8]张毅民,吕学斌,万先凯,等.一株纤维素分解菌的分离及其粗酶性质研究[J].华南农业大学学报,2005,26(2):69-72.
Zhang Y M,Lü X B,Wang X Ket al.Isolation of a cellulose-decomposing microorganisms and investigation of its crude enzyme properties[J].J.South China Agric.Univ.,2005,26(2):69-72.
[9]孟会生,刘卫星,洪坚平.纤维素分解菌群的筛选组建与羧甲基纤维素酶活初探[J].山西农业大学学报,2006,26(1):27-31.
Meng H S,Liu W X,Hong J P.Research on screening of cellulose-decomposing strains and activity of CMCase [J].J.Shanxi Agric.Univ.,2006,26(1):27 -31.
[10]徐昶,龙敏南,邬小兵,等.高产纤维素酶菌株的筛选及产酶条件研究[J].厦门大学学报,2005,44(1):107-110.
Xu C,Long M N,Wu X Bet al.Screening and characterization of a high cellulase producing[J].J.Xiamen Univ.,2005,44(1):107-110.
[11]蔡凤.黑曲霉纤维素酶的纯化及酶学性质研究[D].南京:南京理工大学硕士学位论文,2006.
Cai F.Purification and properties of cellulase from Aspergillus[D].Nanjing:MS thesis,Nanjing University of Science and Technology,2006.
[12]王洪媛,范丙全.三株高效秸秆纤维素降解真菌的筛选及其降解效果研究[J].微生物学报,2010,50(7):870-875.
Wang H Y,Fang B Q.Screening of three straw-cellulose degrading microorganism[J].Acta Microbiol.Sin.,2010,50(7):870-875.
[13]殷中伟,范丙全,任萍.纤维素降解真菌Y5的筛选及其对小麦秸秆降解效果[J].环境科学,2011,32(1):247-252.
Yin Z W,Fan B Q,Ren P.Isolation and identification of a cellulose degrading fungus Y5 and its capability of degradating wheat straw[J].Environ Sci.,2011,32(1):247 -252.
[14]孙宪昀,陈家楠.纤维素化学的现状与发展趋势[J].纤维素科学与技术,1995,3(1):1-10.
Sun X Y,Chen J N.The situation and development trend of Cellulose chemistry[J].J.Cell.Sci.Technol.,1995,3(1):1 -10.
[15]Rashid M H,Rajoka M I,Siddiqui K Set al.Kinetic properties of chemically modified β-glucosidase from Aspergillus niger280[J].Pakistan J.Zool.,1997,29:354 -363.
[16]曲音波,刘自勇.青霉木质纤维素降解酶系研究进展[J].应用与环境生物学报,2007,13(5):736-740.
Qu Y B,Liu Z Y.Progress in research of lignocellulose-degrading enzymes from penicillium [J].Chin.J .Appl.Environ.Biol.,2007,13(5):736 -740.
[17]中国科学院南京土壤研究所微生物室.土壤微生物研究法[M].北京:科学出版社,1985.
Academy of Science Nanjing Institute Microorganism Room.Methodology of research soil microorganism Chinese[M].Beijing:Science Press,1985.
[18]魏景超.真菌鉴定手册(第一版)[M].上海:上海科学技术出版社,1979.501-512.
Wei J C.Fungal identification manual.First section[M].Shanghai:Shanghai Scientific and Technical Publishers,1979.501-512.
[19]孔华忠.中国真菌志(第三十五卷)—青霉属及其相关有性型属(第一版)[M].北京:科学出版社,2007.197-199.
Kong H Z.Flora fungorum sinicorum(vol.35)– penicillium et teleomorphicognati.First section[M].Beijing:Science Press,2007.197-199.
[20]White T J,Bruns T,Lee Set al.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics:A guide to methods and application[M].San Diego:Academic Press,1990.315 -322.
[21]Raeder U,Broda P.Rapid preparation of DNA from filamentous fungi[J].Lett.Appl.Microbiol.,1985,1:17 -20.
[22]吴志红,汪天虹,黄卫.简便易行的丝状真菌染色体DNA提取法[J].菌物系统,2001,20(4):575-577.
Wu Z H,Wang T H,Huang W.A simplified method for chromosome DNA preparation from filamentous fungi[J].Mycosystema,2001,20(4):575-577.
[23]Ghose T K.Measurement of cellulose activities[J].Pure Appl.Chem.,1987,59:257-268.
[24]Wyk V,Mohulatsi M.Biodegradation of wastepaper by cellulase fromTrichooderma viride[J].Biores.Technol.,2003,86(1):21-23.
[25]Bailey M J,Biel Y P,Poutanen K J.Znterlatoratoratory Testing of methods for assay of xylamase [J].J.Biotechnol.,1992,23:257-270.
[26]Laura P,Wendicuti Castro.Themostable xylanases produced at 37℃ and 45℃ by a themotolerantAspergillusstrains[J].FFMS Microbiol.Lett.,1997,146:97 -102.
[27]刘世利,汪天虹,秦楠,等.产适冷木聚糖酶的海洋产青霉的筛选和诱变[J].菌物系统,2003,22(2),272-276.
Liu S L,Wang T H,Qin Net al.Isolation and mutagenesis of cold-adaptive xylanase-producing penicillium strain[J].Mycosystema,2003,22(2):272-276,
[28]侯运华.适冷海洋青霉纤维素酶和半纤维素酶的分子生物学研究[D].济南:山东大学博士学位论文,2006.
Hou Y H.Molecular research on the cellulase and hemicellulase of a novel cold-adaptive marine Pencillium FS010[D].Jinan:PhD dissertation,Shandong University,2006.
[29]王希国,杨谦,燕红,张鲁进.梅林青霉液态发酵生产纤维素酶条件的研究[J].农业环境科学学报,2007,26:97-100.
Wang X G,Yang Q,Yan H,Zhang L J.Cellulase production byPenicillium meliniiinliquid state fermentation [J].J.Agro-Environ.Sci.,2007,26:97 -100.
[30]王仪明,张宗舟,蔺海明,等.绿色木霉固态发酵产纤维素酶活力的研究[J].草业科学,2009,26(5):123-127.
Wang Y M,Zhang Z Z,Lin H Met al.Study on cellulase activity produced by solid-state fermentation ofTrichoderma viride[J].Pratac.Sci.,2009,26(5):123 -127.
[31]史玉英,沈其荣,娄无忌,等.纤维素分解菌群的分离和筛选[J],南京农业大学学报,1996,19(3):59-62.
Shi Y Y,Shen Q R,Lou W Jet al.Isolation and screening of cellulose decomposing mixing strains [J].J.Nanjing Agric.Univ.,1996,19(3):59-62.
[32]申轲.木聚糖酶产生菌的筛选及产酶条件的研究[D].广西:广西大学硕士学位论文,2010.
Shen K.Screening of xylanase producing strain and study on its producing condition[D].Guangxi:MS thesis,Guangxi University,2010.
[33]冯蕾,宋彦波,徐晓立,等.赭绿青霉Sp1菌株木聚糖酶最佳产酶条件及部分酶学特性研究[J].新疆农业科学,2010,47(8):1606-1614.
Feng L,Song Y B,Xun X Let al.Fermentation and characteristics of a xylanase producing strainPenicillium ochrochloronSp1[J].Xinjiang Agric.Sci.,2010,47(8):1606 -1614.
[34]赵峰,聂小琦,余丽娟,等.木聚糖酶高产菌株的筛选及其产酶营养条件研究[J].酿酒科技,2008,9:36-38.
Zhao F,Nie X S,Yu L Jet al.Screening of fungal strain with high yield of xylanase and study of its nutritional condition for enzyme production[J].Liquor-Mak.Sci.Technol.,2008,9:36-38.
猜你喜欢
林产化学与工业(2022年4期)2022-09-13
玻璃纤维(2022年1期)2022-03-11
食品安全导刊(2021年29期)2021-11-10
粮食加工(2021年5期)2021-11-01
湖南饲料(2019年5期)2019-10-15
新农业(2019年16期)2019-09-05
中学化学(2019年2期)2019-07-08
天津造纸(2018年2期)2019-01-21
山东工业技术(2016年15期)2016-12-01
中国医科大学学报(2015年10期)2015-03-01
