
主养草鱼高密度池塘溶氧收支平衡的研究
2013-04-19 08:49龚望宝余德光王广军张敬旺李志斐郁二蒙王海英
水生生物学报 2013年2期
龚望宝 余德光 王广军 谢 骏 张敬旺李志斐 郁二蒙 王海英
(中国水产科学研究院珠江水产研究所, 农业部热带亚热带水产资源利用与养殖重点实验室, 广州 510380)
主养草鱼高密度池塘溶氧收支平衡的研究
龚望宝 余德光 王广军 谢 骏 张敬旺李志斐 郁二蒙 王海英
(中国水产科学研究院珠江水产研究所, 农业部热带亚热带水产资源利用与养殖重点实验室, 广州 510380)
采用原位生态学的方法测定广东省中山市9口主养草鱼高密度池塘中浮游植物光合作用产氧量、水柱呼吸耗氧量、底泥呼吸耗氧量和鱼呼吸耗氧量, 并用数学模型计算增氧机增氧量及用差减法计算大气扩散作用引起的得氧或失氧, 对高密度养殖池塘中溶氧收支平衡状况进行了研究。结果显示: 在水深为1.5—2.0 m的主养草鱼高密度池塘中, 光合作用产氧量随着水深的增加而显著降低(P<0.05), 底层出现负值呈现氧债现象。水呼吸耗氧量在表层、中层和底层之间没有显著差异(P>0.05)。表层水光合作用产氧量显著大于水呼吸耗氧量(P<0.05), 而中层和底层水光合作用产氧量却显著小于水呼吸耗氧量(P<0.05)。在主养草鱼高密度池塘溶氧的收入中, 浮游植物光合作用产氧量、增氧机增氧量和大气扩散溶入氧量分别占总溶氧来源的44.7%、42.3%和13.0%, 机械增氧作用已接近光合作用, 成为溶氧来源的主要贡献者; 在池塘溶氧的支出中,水呼吸、鱼呼吸和底泥呼吸耗氧量分别占总耗氧量的45.9%、45.0%和9.1%, 鱼呼吸耗氧与水呼吸耗氧相当,成为水体中氧气的主要消耗者。结果表明在草鱼高密度养殖过程中, 合理使用机械增氧是池塘溶氧管理的有效措施。
草鱼; 高密度; 池塘养殖; 溶氧收支
高密度精养是目前珠江流域池塘养殖的主要生产方式[1]。由于大量投饵, 高密度精养造成池塘底部残饵和养殖动物粪便等物质大量积累, 池塘底部水体缺氧现象严重[2,3], 溶氧有效管理对池塘养殖动物健康维护和水环境调控日显重要[4]。国内外在载鱼量为150 kg/km2以下的斑点叉尾(Ictalurus punctatus)、日本鳗鲡(Anguilla japonica)和“四大家鱼”等池塘开展了溶氧变化规律及模型预测研究[5—9],表明光合作用产氧是池塘溶氧的主要来源和水呼吸是主要的耗氧因子, 这一结果为中低密度养殖池塘溶氧管理提供了重要的参考依据。但是, 在高密度精养池塘, 增氧机的高强度运行使池塘溶氧变化特征发生了改变[10], 溶氧是池塘水质指标中的一项重要变量, 亦是高密度精养池塘人工调控的重要因子之一[11], 而对高密度池塘溶氧收入和支出平衡的研究是高产精准管理的关键。
草鱼(Ctenopharyngodon idella)是我国大宗淡水养殖的重要鱼类之一, 主养草鱼高密度池塘的载鱼量可达300 kg/km2以上[12], 但有关主养草鱼高密度池塘溶氧收支的研究还未见报道。本研究采用原位生态学的方法测定广东省中山市9口主养草鱼高密度池塘中浮游植物光合作用产氧量、水柱呼吸耗氧量、底泥呼吸耗氧量和鱼呼吸耗氧量, 并用数学模型计算增氧机增氧量及用差减法计算大气扩散作用引起的得氧或失氧, 研究主养草鱼高密度池塘养殖期间增氧与耗氧因子的变化及溶氧收支, 以期为草鱼高密度养殖池塘的溶氧管理提供基础数据。
1 材料与方法
1.1 研究地点及草鱼养殖条件
实验于2011年6月至9月在广东省中山食品水产进出口集团有限公司坦洲养殖基地进行。选取9口主养草鱼池塘作为测试对象, 鱼池平均载鱼量为311.26 kg/km2, 草鱼占总载鱼量的80%左右, 规格为2.0—3.5 kg, 混养少量鳙(Aristichthys nobilis)和鲮(Cirrhina molitorella)。水源来自西江支流, 实验期间的水温在29.4—32.2℃, pH在7.47—8.62, 溶氧在4.70—10.43 mg/L。实验塘的基本情况见表1。
1.2 测定与计算方法
浮游植物光合作用产氧和水柱呼吸作用耗氧
采用黑白瓶测氧法测定浮游植物光合作用产氧和水柱呼吸作用耗氧。黑白瓶分表层、中层和底层进行悬挂, 表层悬挂在距水面下10 cm, 中层为水深的中间, 底层距池塘底部10 cm, 24h后取瓶用Winkler碘量法测溶解氧含量。取水样同时测水温。白瓶与黑瓶溶氧量之差为光合作用产氧量, 初始含氧量与黑瓶溶氧量之差为水呼吸耗氧量。养殖池塘日平均光合作用产氧量或水柱呼吸耗氧量按下式计算[13]: Q = [(Q表+Q中)/2×H中+(Q中+Q底)/2×(H−H中)]/H (1)式中, Q表、Q中、Q底为表、中、底层水每日每升水体的光合作用产氧量或水呼吸耗氧量(mg/L), H 为水深(m), H中为中层黑白瓶挂取深度(m), Q为平均光合作用产氧量或水柱呼吸耗氧量[mg/(L·d)]。
全水柱池水的光合作用产氧量或水呼吸耗氧量则为Q×相应塘的平均水深。
底泥呼吸耗氧 底泥呼吸耗氧用底泥呼吸器现场原位测定, 底泥呼吸器参照李德尚等[14]描述的进行制作。以6h为时间间隔测定底泥呼吸耗氧的周日节律, 取样测定方法和计算方法采用张敬旺等[15]所用的方法。底泥耗氧率按下式计算:
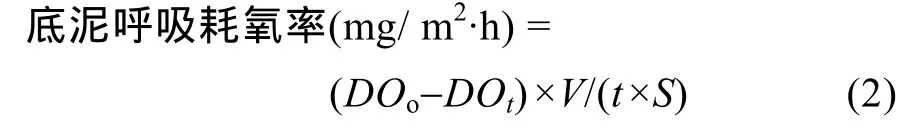
式中, DOo为初始溶氧浓度(mg/L), DOt为t小时后溶氧浓度(mg/L), t为取样间隔时间6h, V 为底泥呼吸器体积29.5 L, S为底泥呼吸器底面积 0.152 m2。
池塘底泥耗氧量[g/ (m2·d)]则为单位时间内单位面积的底泥呼吸耗氧率×24h。
鱼的呼吸耗氧 草鱼呼吸耗氧率测定于取样的同时在该基地进行。实验用水为充分曝气的自来水, 在自然温度下采用静水式装置(黑色圆形塑料桶,容积为1000 L)测得。取与实验池塘中相同规格的单个实验鱼[(2.20 ± 0.05) kg, N =6]放入每个装置中,用塑料薄膜封口, 共3个重复, 同时设定空白对照组, 以测定水呼吸耗氧量。容器内置一个额定功率为12W的小水泵进行搅水, 保证容器内的溶氧分布均匀, 每隔3h用虹吸法在每个装置的3个不同位置采集水样。采用Winkler碘量法测定溶解氧, 计算平均值。当溶解氧降低至2.0 mg/L左右停止实验, 并观察实验鱼活动正常。实验结束后, 称其体重。草鱼的单位鱼体重耗氧率按下式计算[16]:
单位鱼体重耗氧率[mg/ (g·h)] =
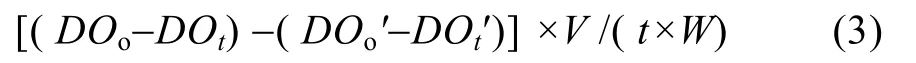
式中, DOo、DOt分别为初始溶氧浓度和t 小时后溶氧浓度(mg/L), DOo′、DOt′分别为对照组初始溶氧浓度和t 小时后溶氧浓度(mg/L), V为呼吸室内水体体积(L), W为草鱼体重(g), t为实验时间(h)。
池塘鱼呼吸耗氧量[g/(m2·d)]则为单位时间内单位鱼体重的耗氧率×池塘载鱼量×24h再除以池塘面积。

表1 主养草鱼高密度池塘基本情况Tab.1 Basic conditions of C.idella-farming ponds
机械增氧 9口池塘均安装1.5 kW的叶轮式增氧机, 各池塘的增氧机数量(表1)。实验期间, 每天从晚上22:00开至凌晨06:00, 共8h。根据国家渔业机械仪器质量监督检验中心实验室实测数据, 1.5 kW叶轮增氧机的标准氧传递速率在2.16—2.59 kg/h,本实验参考顾海涛等[17]取2.30 kg/h, 实验池塘水温下的氧传递速率(Oxygen transfer rate, OTR)根据文献[18]按照下式进行计算:
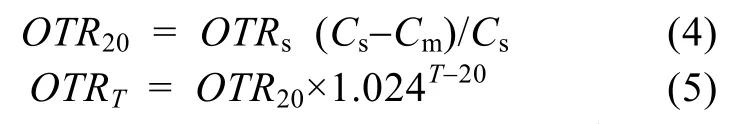
式中, Cs为水温20℃和1个标准大气压时氧的饱和浓度(mg/L), SC/T 6009-1999推荐为9.17 mg/L; Cm为最低耐受氧浓度2.0 mg/L; T为实验池塘水温; OTRs为标准氧传递速率, 即2.30 kg/h; OTR20为水温20℃时的氧传递速率(kg/h); OTRT为实验池塘水温下的氧传递速率(kg/h)。
水体与大气的氧交换(溶入和释放) 水体与大气的氧交换量用差减法[19]求得, 即溶氧的总消耗量减去浮游植物光合作用产氧量和增氧机增氧量。
1.3 数据分析
采用Statistical 6.0对所有数据进行单因子方差分析(One-way ANOVA)。当差异显著时, 再用Duncan’s多重比较对平均数进行比较。表中数据表示为平均数±标准差, 显著水平为P<0.05。
2 结果
2.1 草鱼养殖池塘光合作用产氧量与水呼吸耗氧量
实验池塘各水层光合作用产氧量与水呼吸耗氧量(表2)。各池的透明度在15—29 cm, 平均为(21± 4) cm。从表2可以看出, 池塘表层、中层和底层的光合作用产氧量存在明显差异, 随着水深的增加, 光合作用产氧量显著降低(P<0.05)。表层产氧量[(16.85 ± 3.49) mg/(L·d)]显著大于中层[(2.03 ± 1.77) mg/(L·d)]和底层[(−0.12 ± 0.40) mg/(L·d)](P<0.05),底层出现负值呈现氧债现象。表层、中层和底层的水呼吸耗氧量分别为(5.76 ± 2.01)、(5.50 ± 1.63)和(4.78 ± 1.53) mg/(L·d), 各水层之间没有显著差异(P>0.05)。表层水光合作用产氧量显著大于表层水呼吸耗氧量(P<0.05), 而中层和底层的光合作用产氧量均显著小于中层和底层的水呼吸耗氧量(P<0.05)。
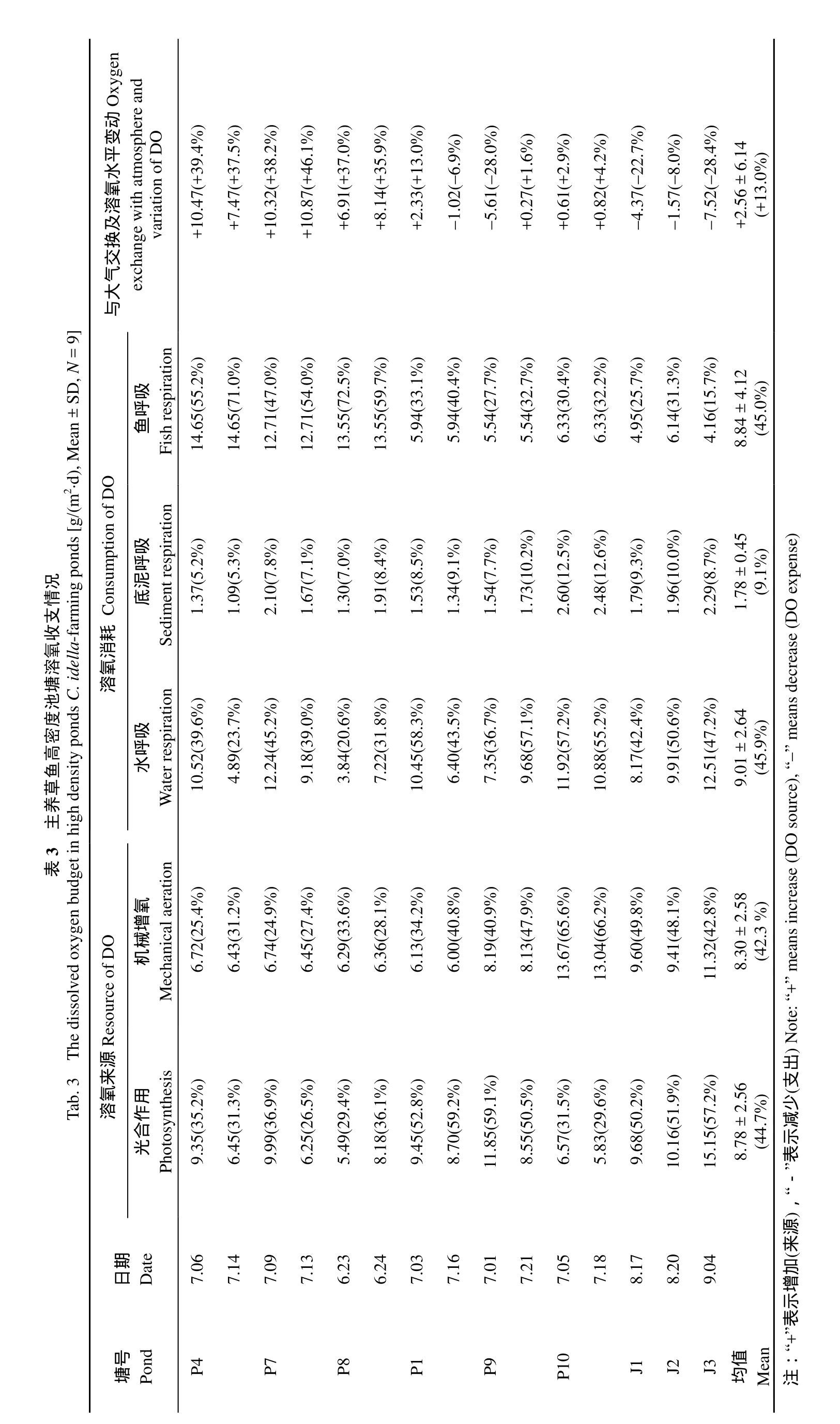
实验池塘日平均光合作用产氧量与水柱呼吸耗氧量(表3)。从表3可以看出, 主养草鱼高密度池塘光合作用产氧量在5.49—15.15 g/(m2·d), 平均为8.78 ± 2.56 g/(m2·d); 水柱呼吸耗氧量在3.84—12.51 g/(m2·d), 平均为(9.01 ± 2.64) g/(m2·d)。
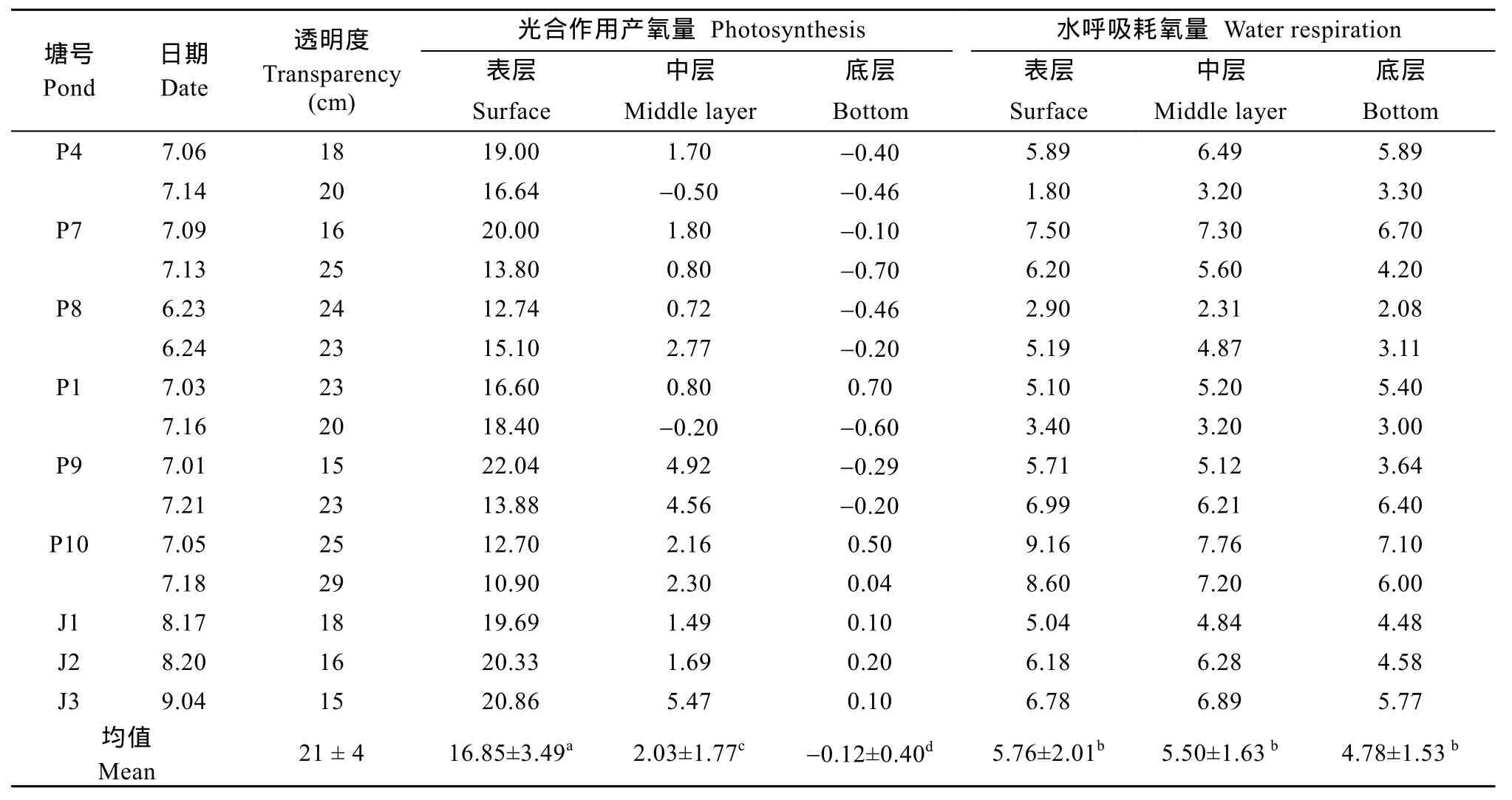
表2 主养草鱼高密度池塘光合作用产氧量和水呼吸耗氧量Tab.2 Oxygen production of photosynthesis and oxygen consumption of water respiration in high density C.idella-farming ponds (mg/L·d) (Mean ± SD, N = 9)
2.2 增氧机增氧
根据实验池塘配置增氧机的数量、功率、开机持续时间及氧传递速率, 按照公式(4)和(5)计算出增氧机的增氧量(表3)。实验池塘增氧机增氧量在6.00—13.67 g/(m2·d), 平均为(8.30 ± 2.58) g/(m2·d)。
2.3 底泥耗氧
实验池塘的底泥耗氧率(表3、表4)。从表3可以看出, 主养草鱼高密度池塘底泥耗氧量在1.09—2.60 g/(m2·d), 平均为1.78 ± 0.45 g/(m2·d)。从表4可以看出, 底泥耗氧率在0.042—0.095 g/(m2·h), 有明显的日变化趋势。10:00和16:00的底泥耗氧率显著高于04:00的底泥耗氧率(P<0.05)。
2.4 鱼呼吸耗氧
草鱼(2.2 ± 0.05) kg/ind呼吸耗氧率和各池塘鱼呼吸耗氧量(表3、表5)。从表3可以看出, 草鱼呼吸耗氧量在4.16—14.65 g/(m2·d), 平均为(8.84 ± 4.12) g/(m2·d)。从表5可以看出, 草鱼呼吸耗氧率昼夜波动较大, 耗氧率的高峰出现在白天, 低谷出现在夜间。9:00、12:00、15:00和18:00点的耗氧率显著高于03:00和06:00的耗氧率。草鱼的平均耗氧率为0.110 mg/(g·h)。
2.5 池塘溶氧的收支平衡
池塘溶氧的收入和支出决定着池塘的溶氧平衡状况。各池塘的收支结果(表3)。在溶氧的来源中, 光合作用产氧量和增氧机增氧量分别占溶氧收入的44.7%和42.3%。增氧机增氧作用接近光合作用产氧作用, 成为池塘溶氧收入的主要贡献者。
在溶氧的消耗中, 水呼吸、鱼呼吸和底泥呼吸耗氧分别占溶氧支出的45.9%、45.0% 和9.1%。从结果可以看出, 鱼呼吸耗氧与水呼吸耗氧的比例相当, 成为池塘溶氧的主要耗氧因子, 底泥呼吸耗氧所占比例最小。
各池塘水体与大气交换的结果(表3)。当交换值>0时, 表明池塘通过大气溶解而得氧; 当交换值<0时, 表明池塘通过大气扩散而逸出氧。从表中可以看出, 各池的数值波动较大, 平均值为13.0%, 表明草鱼池塘通过大气扩散作用得氧。
3 讨论
早于20世纪80年代就有研究表明[20], 传统鱼池(载鱼量为84 kg/km2)的溶氧收入中, 浮游植物光合作用占90.3%; 支出中, 水呼吸、鱼呼吸和底泥呼吸分别占总耗氧量的72.9%、16.1%和0.6%。本研究结果显示, 在主养草鱼高密度池塘(平均载鱼量为311.26 kg/km2) 的溶氧收入中, 浮游植物光合作用产氧量和增氧机增氧量分别占总溶氧来源的44.7%和42.3%; 在溶氧的支出中, 水呼吸、鱼呼吸和底泥呼吸耗氧量分别占总耗氧量的45.9%、45.0%和9.1%。与传统鱼池溶氧收支相比, 主养草鱼高密度池塘的溶氧收支发生了很大改变, 增氧机增氧作用接近浮游植物光合作用, 为溶氧主要来源; 鱼类呼吸耗氧比例上升, 与水呼吸耗氧比例相当, 为主要的耗氧因子; 底泥呼吸耗氧比例亦有所增加。
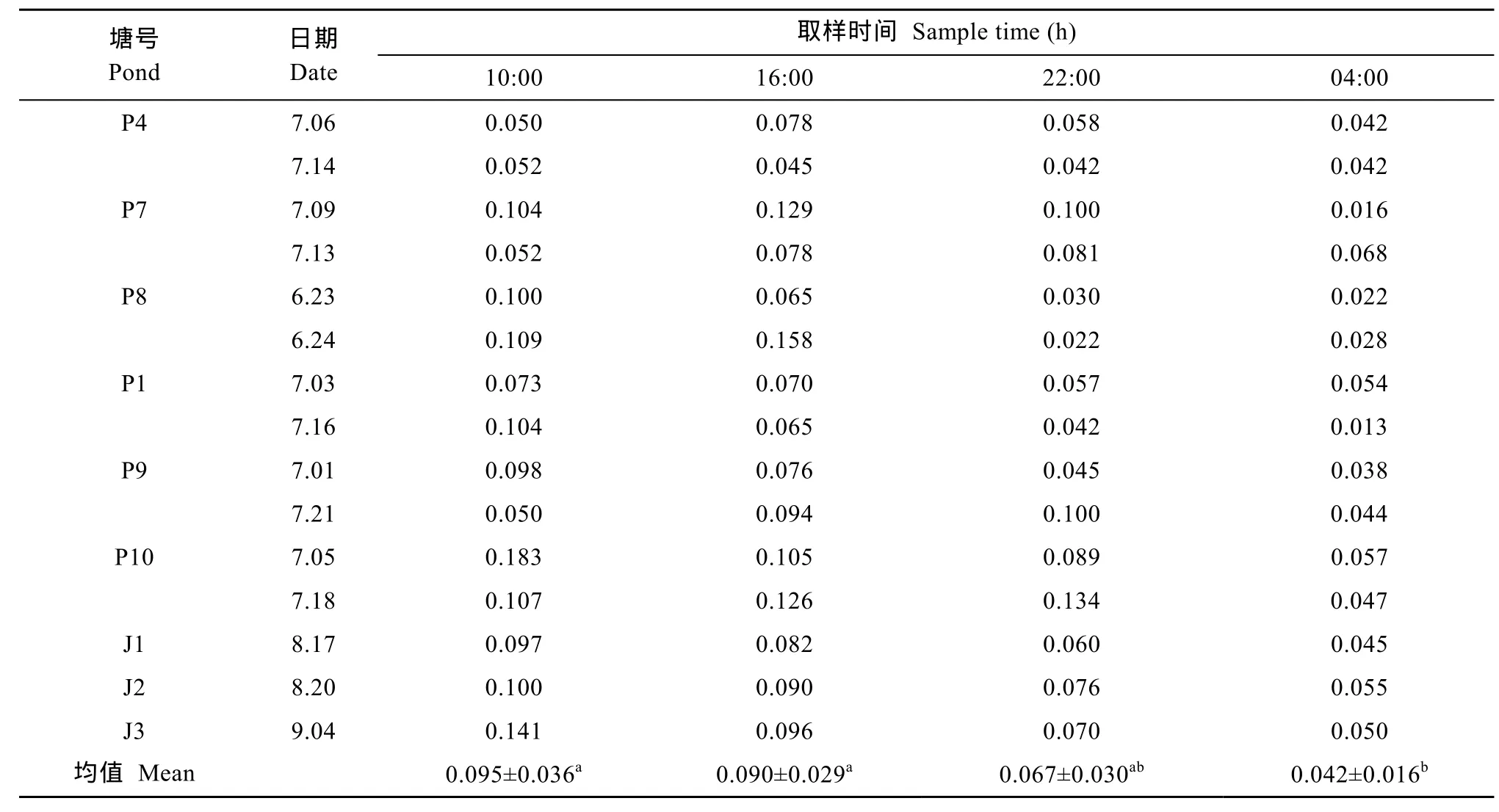
表4 主养草鱼高密度池塘底泥耗氧率的日变化Tab.4 Diurnal variation of sediment respiration rates g/(m2·h) in high density C.idella-farming ponds (Mean ± SD, N = 9)
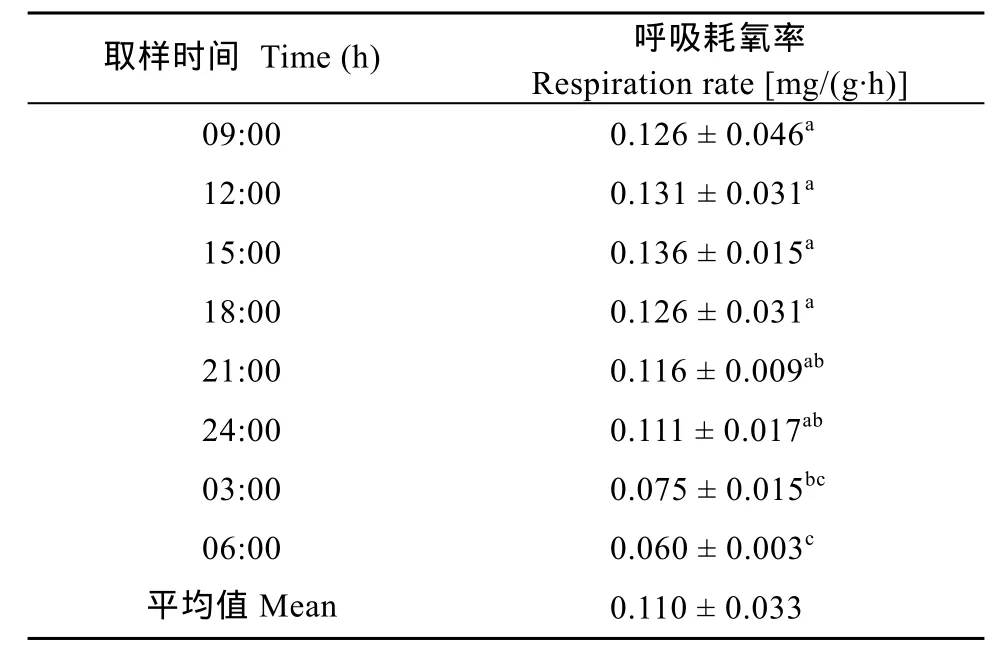
表5 草鱼耗氧率的日变化Tab.5 Diurnal variation of respiration rates [mg/(g·h)] of adult C.idella(Means ± SD, N = 3)
增氧机是主养草鱼高密度池塘的重要设备, 其增氧量占总溶氧来源的42.3%。周劲风等[21]对主养加州鲈(Micropterus salmoides)精养池塘水质模型的研究表明, 人工增氧量略大于浮游植物光合作用产氧量, 增氧机增氧已是精养池塘重要增氧措施, 精养养殖就是增氧养殖。在中国对虾(Penaeus chinensis)养殖池塘中[22], 虾池溶解氧处于支出大于收入的不平衡状态, 48%的溶氧需要由机械增氧与空气溶解补给。机械增氧成为高密度养殖池塘溶氧的主要来源, 不仅解决了鱼虾的氧气需求, 而且还对于残饵和粪便等有机物在池塘底部大量沉积起分解促进作用。由于底部阳光缺乏, 少有藻类光合作用, 增氧机通过搅拌和曝气, 充分混合上层氧盈水及下层氧债水, 促进养殖水体得氧, 提高池塘底部的溶氧水平,从而提高底部的氧化还原电位, 降解底泥有害物质的积累[23,24]。
在载鱼量为74.63—150 kg/ km2的传统养鱼池中, 养殖鱼类的呼吸耗氧量一般占总耗氧量的5—22%[25]。在本研究中, 鱼呼吸耗氧量占总耗氧量的比例高达45%, 成为主要的耗氧因子。这种耗氧方式主要随放养密度的变化而变化。本文中的平均载鱼量为311.26 kg/km2, 是传统养殖池塘载鱼量的2—3倍, 养殖密度的提高导致鱼呼吸耗氧量的显著增加。在对虾池[26]中也有相似的研究结果, 放养密度为20 kg/km2的中国对虾池, 其对虾耗氧量仅占总耗氧量的3.7%; 而放养密度为200 kg/km2的南美白对虾池, 对虾耗氧所占比例达到72.7%, 成为主要的耗氧因子。
养鱼池塘底泥原位测定耗氧速率一般为0.33—1.60 g/(m2·d)[6,19,27]。本文采用底泥呼吸器原位测定的底泥耗氧率在1.09—2.60 g/(m2·d), 平均为1.78 g/(m2·d)。研究结果比通常的鱼池底泥耗氧率较高,这可能与高密度养殖有关。在高密度养殖中, 残饵和粪便等有机物在池塘底泥中沉积, 而有机物的分解消耗大量的氧气, 池塘底泥耗氧量随着有机物含量的增加而增加[28]。Madenjian[29]报道, 在夏威夷载虾量为28 kg/km2的对虾养殖池塘中, 底泥耗氧量为7.15 g/(m2·d)。Steeby, et al.[30]报道, 在美国密西西比州西北部的斑点叉尾养殖池塘中, 载鱼量仅为56 kg/km2, 但其底泥原位测定耗氧率在1.49—23.09 g/(m2·d), 平均为11.47 g/(m2·d)。本研究中的池塘载鱼量为311.26 kg/km2, 但是底泥耗氧率却明显要低,一方面与主养草鱼池塘投喂较低的能量饲料导致底泥有机质含量低有关[20]。不同饵料在投喂过程中产生的有机废物在颗粒大小、有机质溶出速率、沉降速率等存在较大差异[31], 导致沉积物中有机物含量有很大差异[32], 进而影响沉积物耗氧。另一方面与套养鳙和鲮等可消耗部分有机物有关[33]。
实验结果显示主养草鱼高密度池塘表层浮游植物光合作用产氧量显著高于水呼吸耗氧量, 而中层和底层的光合作用产氧量均显著小于其水呼吸耗氧量。同时, 本研究中的池塘平均透明度为21 cm, 说明池水中浮游生物和有机物丰富。类似的研究结果也出现在养鳗池[6]、“四大家鱼”池[8]和虾池[12,21], 表明自1 m水深以下, 光合作用极弱, 呈现氧债现象。这主要是由于养殖水体中大量有机质分解以及浮游动物和细菌等耗氧所导致的。姚宏禄[9]认为水体光合作用产氧量(P)和水呼吸耗氧量(R)及其比率(P/R)是高产鱼池溶氧变化的主要矛盾, 这种矛盾与放养密度、营养盐含量和管理技术等有关。
综上所述, 与传统鱼类养殖池塘相比, 主养草鱼高密度池塘的溶氧收支平衡已发生了很大改变,机械增氧作用接近光合作用, 成为主要的溶氧来源;鱼呼吸成为主要的耗氧因子, 底泥呼吸耗氧量增加。在养殖过程中, 合理采取机械增氧措施是高密度养殖池塘水质管理的有效措施。本研究虽然通过原位生态学的方法对草鱼高密度池塘养殖期间增氧与耗氧因子的变化及溶氧收支有一个较全面的认识,但下一步需要对该类型养殖池塘溶氧的昼夜和季节变化规律进行系统研究。
[1] Zhou J F, Wen Y M, Liang Z Q.Nutrient budget of intensive fish pond in Pearl River Delta [J].Fisheries Science, 2004, 23(9): 11—15 [周劲风, 温琰茂, 梁志谦.珠江三角洲密养池塘营养物质收支的研究.水产科学, 2004, 23(9): 11—15]
[2] Avnimelech Y, Mozes N, Diab S, et al.Rates of organic carbon and nitrogen degradation in intensive fish ponds [J].Aquaculture, 1995, 134: 211—216
[3] Jing X J, Miao W M, Yuan X H, et al.Review on the biological purification techniques of intensive ponds water quality [J].Chinese Agricultural Science Bulletin, 2008, 24(9): 490—495 [敬小军, 缪为民, 袁新华, 等.精养池塘水质生物净化技术研究综述.中国农学通报, 2008, 24(9): 490—495]
[4] Boyd C E.Water Quality Management for Pond Fish Culture [M].Elsevier, Amsterdam.1982, 318
[5] Boyd C E, Romaire R P, Johnston E.Predicting early morning dissolved oxygen concentration in channel catfish ponds [J].Transactions of the American Fisheries Society, 1978, 107(3): 484—492
[6] Lu M X, Huang Z H, Wu R Q, et al.Primary productivity and energy conversion efficiency in eel pond [J].Journal of Fisheries of China, 2000, 24(1): 37—40 [卢迈新, 黄樟翰,吴锐全, 等.养鳗池塘的初级生产力和能量转换效率.水产学报, 2000, 24(1): 37—40]
[7] Chang W Y B, Ouyang H.Dynamics of dissolved oxygen and vertical circulation in fish ponds [J].Aquaculture, 1988, 74: 263—276
[8] Lei Y Z, Yu S M, Xu J.Studies on the water quality of the high-yield fishponds in He Lie Commune, Wuxi Part 1.Chemistry of fishpond water and primary productivity [J].Journal of Fisheries of China, 1983, 7(3): 185—199 [雷衍之,于淑敏, 徐捷.无锡市河埒口高产鱼池水质研究 I.水化学和初级生产力.水产学报, 1983, 7(3): 185—199]
[9] Yao H L.Fluctuations of dissolved oxygen in integrated fish culture ponds [J].Acta Hydrobiologica Sinica, 1988, 12(3): 199—211 [姚宏禄.综合养鱼高产池塘的溶氧变化周期.水生生物学报, 1988, 12(3): 199—211]
[10] Chen F, Sun Z, Wang Y B.A study for variation of dissolved oxygen in different breeding mode ponds of Protunus trituberculatus in summer [J].Journal of Zhejiang Ocean University (Natural Science), 2011, 30(3): 221—225 [陈芳, 孙忠, 王跃斌.三疣梭子蟹不同养殖模式池塘夏季溶解氧变化特征的研究.浙江海洋学院学报(自然科学版), 2011, 30(3): 221—225]
[11] Ruiz-Velazco J M J, Hernández-Llamas A, Gomez-Muñoz V M.Management of stocking density, pond size, starting time of aeration, and duration of cultivation for intensive commercial production of shrimp Litopenaeus vannamei [J].Aquaculture Engineering, 2010, 43(3): 114—119
[12] Lu Z C, Li F, Zhang C S.High yield test in intensive grass carp ponds [J].Current Fisheries, 2010, (6): 68—69 [卢正臣,李峰, 张春山.池塘精养草鱼种高产试验.当代水产, 2010, (6): 68—69]
[13] Wang W D, Zang W L, Dai X L, et al.The incomings and outgoings budget of dissolved oxygen in Penaeus monodon desalting culture ponds in estuarine area [J].Journal of Shanghai Fisheries University, 2000, 9(2): 97—102 [王为东,臧维玲, 戴习林, 等.河口区斑节对虾淡化养殖塘溶氧收支平衡状况.上海水产大学学报, 2000, 9(2): 97—102]
[14] Li D S, Lu J R, Xu N, et al.A device of in situ sediment respiration determing apparatus used in pond [J].Journal of Ocean University of Qingdao, 1998, 28(2): 205—209 [李德尚, 卢敬让, 徐宁, 等.一种用于现场测定的池塘底泥呼吸器.青岛海洋大学学报, 1998, 28(2): 205—209]
[15] Zhang J W, Xie J, Li Z F, et al.The pond’s sediment oxygen demand and the relationship with physical and chemical factors [J].Freshwater Fisheries, 2012, 42(3): 37—43 [张敬旺,谢骏, 李志斐, 等.家鱼池塘底泥耗氧率与理化因子的相关性分析.淡水渔业, 2012, 42(3): 37—43]
[16] Lei S J.Effect of salinity and body weight on the oxygen consumption of Oreochromis niloticus × O.mossambicus [J].Chinese Journal of Applied Ecological, 2002, 13(6): 739—742 [雷思佳.盐度与体重对台湾红罗非鱼耗氧率的影响.应用生态学报, 2002, 13(6): 739—742]
[17] Gu H T, He Y P, Men T, et al.Study on aerobic performance of impeller aerator [J].Fishery Modernization, 2011, 38(4): 41—44 [顾海涛, 何雅萍, 门涛, 等.叶轮式增氧机性能研究.渔业现代化, 2011, 38(4): 41—44]
[18] Santa K D, Vinatea L.Evaluation of respiration rates and mechanical aeration requirements in semi-intensive shrimp Litopenaeus vannamei culture ponds [J].Aquaculture Engineering, 2007, 36: 73—80
[19] Xu N, Li D S, Dong S L.Diel balance of DO in mariculture ponds [J].Journal of Fishery Sciences of China, 1999, 6(1): 69—74 [徐宁, 李德尚, 董双林.海水养殖池塘溶氧平衡的实验研究.中国水产科学, 1999, 6(1): 69—74]
[20] Wang W.Culture and Enhancement of Fish [M].Beijing: China Agriculture Press.2000, 53—76 [王武.鱼类增养殖学.北京: 中国农业出版社.2000, 53—76]
[21] Zhou J F, Wen Y M, Li Y C.Establishment of water quality model of farming ponds in the Pearl River delta and itsapplication [J].Acta Scientiarum Naturalium Universitatis Sunyatseni (Nature Science), 2004, 43(5): 103—106 [周劲风,温琰茂, 李耀初.珠三角养殖池塘水质模型建立及其应用研究.中山大学学报: 自然科学版, 2004, 43(5): 103—106]
[22] Zang W L, Dai X L, Zhu Z G, et al.The budget of dissolved oxygen in shrimp culture ponds [J].Acta Oceanologica Sinica, 1995, 17(4): 137—141 [臧维玲, 戴习林, 朱正国,等.中国对虾池溶解氧的收支平衡状态.海洋学报(中文版), 1995, 17(4): 137—141]
[23] Zhou J F, Wen Y M, Liang Z Q.Water quality characteristics of intensive fish pond in Pearl River Delta [J].Fisheries Science, 2004, 23(12): 13—17 [周劲风, 温琰茂, 梁志谦.珠江三角洲密养池塘的水质特点分析.水产科学, 2004, 23(12): 13—17]
[24] Zhou J F, Wen Y M, Li Y C.Nutrients diffusion at the water-sediment interface of farming ponds by indoor simulation experiment: ⅠDifferent of nitrogenous compounds [J].Journal of Ago-Environment Science, 2006, 25(3): 786—791 [周劲风, 温琰茂, 李耀初.养殖池塘底泥-水界面营养盐扩散的室内模拟研究: Ⅰ氮的扩散.农业环境科学学报, 2006, 25(3): 786—791]
[25] Liu H Y, Qu K M, Ma S S.Survey of both the variation and the absorption and consumption budget of dissolved oxygen in culture ponds [J].Marine Fisheries Research, 2005, 26(2): 79—84 [刘海英, 曲克明, 马绍赛.养殖水体中溶解氧的变化及收支平衡研究概况.海洋水产研究, 2005, 26(2): 79—84]
[26] Liu H Y, Qu K M, Zhang Q Q, et al.Study on dissolved oxygen consumption both in industrial and pond culture of shrimps [J].Marine Fisheries Research, 2005, 26(5): 52—56 [刘海英, 曲克明, 张前前, 等.对虾工厂化养殖和土池养殖溶解氧消耗研究.海洋水产研究, 2005, 26(5): 52—56]
[27] Lei Y Z, Piao W H, Bai L J, et al.The oxygen consumption rate in mud of fish ponds [J].Journal Dalian Fisheries College, 1992, 6(3—4): 6—13 [雷衍之, 朴文豪, 白禄君, 等.养鱼池底泥耗氧速率的研究.大连水产学院学报, 1992, 6(3—4): 6—13]
[28] Steeby J A, Hargreaves J A, Tucker C S.Factors affecting sediment oxygen demand in commercial Channel catfish ponds [J].Journal of the World Aquaculture Society, 2004a, 35(3): 322—334
[29] Madenjian C P.Patterns of oxygen production and consumption in intensively managed marine shrimp ponds [J].Aquaculture and Fisheries Management, 1990, 21(4): 407—417
[30] Steeby J A, Hargreaves J A, Tucker C S, et al.Modeling industry-wide sediment oxygen demand and estimation of the contribution of sediment to total respiration in commercial channel catfish ponds [J].Aquaculture Engineering, 2004b, 31(3-4): 247—262
[31] Sun Y, Yang Q F, Cui Y, et al.Study of oxygen demand of newborn feed remains and its change during shrimp culture [J].Journal of Fishery Sciences of China, 1996, 3(4): 53—59 [孙耀, 杨勤芳, 崔毅, 等.对虾养殖中新生残饵耗氧动态及其规律的研究.中国水产科学, 1996, 3(4): 53—59]
[32] Jiang Z J, Fang J G, Mao Y Z, et al.Identification of aquaculture-derived organic matter in the sediment associated with coastal fish farming [J].Journal of Fishery Sciences of China, 2012, 19(2): 348—354 [蒋增杰, 方建光, 毛玉泽,等.海水鱼类网箱养殖水域沉积物有机质的来源甄别.中国水产科学, 2012, 19(2): 348—354]
[33] Zhao X B, Yu E M, Wang G J, et al.A comparative study on two polyculture models of grass carp bases on ecological niche and water quality factors [J].Chinese Journal of Ecology, 2010, 29(11): 2187—2191 [赵旭斌, 郁二蒙, 王广军,等.基于生态位和水质因子的草鱼两种混养模式的比较.生态学杂志, 2010, 29(11): 2187—2191]
THE BUDGET OF DISSOLVED OXYGEN IN HIGH DENSITY PONDS MAINLY STOCKED WITH CTENOPHARYNGODON IDELLA
GONG Wang-Bao, YU De-Guang, WANG Guang-Jun, XIE Jun, ZHANG Jing-Wang, LI Zhi-Fei, YU Er-Meng and WANG Hai-Ying
(Key Laboratory of Tropical & Subtropical Fishery Resource Application & Cultivation, Ministry of Agriculture, Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510380, China)
To increase the efficiency of dissolved oxygen (DO) management in high density ponds mainly stocked with grass carp (Ctenopharyngodon idella), the present research aimed to characterize the budget of dissolved oxygen in nine fish ponds in Zhongshan, Guangdong province, China.The water depth was 1.5—2.0 m.Photosynthesis of phytoplankton and water respiration rate (WR) were determined by the clear and dark bottles method.The sediment respiration rate (SR) was measured byin siturespirometry.The fish respiration rate was determined by the respiratory chamber method.The mechanical aeration was calculated by predicting models.Atmosphere diffusion was estimated by the subtraction method.The results showed the gross oxygen production (GOP) of photosynthesis showed a decreasing trend as the water depth increased, and the value in the bottom was negative, which was the common oxygen debt phenomenon.There was no significant difference between the WR under the surface, middle layer and bottom (P>0.05).The GOP of photosynthesis in the surface was significantly higher than theWR(P<0.05), but the GOP of photosynthesis was significantly lower than theWRboth in the middle layer and in the bottom (P<0.05).The photosynthesis of phytoplankton and the mechanical aeration were the main contributors to the total oxygen production in high density ponds mainly stocked with C.idella, which accounted for 44.7% and 42.3% of the total oxygen production, respectively, but the contribution of atmosphere diffusion was the lowest and accounted for 13.0%.Among the oxygen consuming factors, the WR was the largest oxygen consumer, which accounted for 45.9% of the total oxygen consumption of water column, followed by fish respiration rate which accounted for 45.0% of the total oxygen consumption, and the proportion ofSRwas the lowest which accounted for 9.1%.
Ctenopharyngodon idella; High density; Pond culture; Dissolved oxygen budget
S963
A
1000-3207(2013)02-0208-09
10.7541/2013.6
2012-10-16;
2012-12-29
国家科技支撑计划课题“珠江流域高产精养池塘健康养殖技术集成与示范 No.2012BAD25B04”; 现代农业产业技术体系建设专项资金 nycytx-49-13; 公益性行业(农业)科研专项“池塘复合净化技术研究与示范 201203083” 资助
龚望宝(1979—), 女, 湖北咸宁人; 博士, 助理研究员; 主要从事水产健康养殖研究。E-mail: gwbli@163.com
余德光, 男, 副研究员; E-mail: gzyudeguang@163.com
猜你喜欢
家庭科学·新健康(2020年7期)2020-08-06
南方农业·中旬(2020年2期)2020-05-28
当代水产(2019年7期)2019-09-03
航天器环境工程(2019年3期)2019-07-01
福建质量管理(2019年2期)2019-01-22
发酵科技通讯(2018年3期)2018-10-16
农家之友(2018年8期)2018-08-24
医药前沿(2018年18期)2018-06-21
海峡科学(2017年4期)2017-07-12
中国海洋大学学报(自然科学版)(2017年7期)2017-06-05
