
白洋淀2种枝角类的摄食生态
2013-10-09 11:52王军霞刘存歧张晶张跃伟王亚斌
河北大学学报(自然科学版) 2013年6期
王军霞,刘存歧,张晶,张跃伟,王亚斌
(1.河北大学生命科学学院,河北保定 071002;2.重庆大学资源及环境科学学院,重庆 400030;3.保定市环境保护局环境保护监测站,河北保定 071002)
浮游动物在水生态系统中起着重要的调控作用.初级生产者(浮游植物)如不迅速被次级生产者(浮游动物)利用将产生水华[1].浮游动物对环境变化比较敏感,与水体质量关系密切,很多种类对污染物有一定的积累和代谢作用.在控藻生物链中,最重要的、最可能压低浮游植物数量的因素是大型浮游动物[2],最有效的为水体中重要的滤食性浮游动物大型枝角类[3],因此枝角类同时也是经典生物操纵生物调控的关键生物类群.但要使控藻效果达到最佳,就必须保证大型浮游动物的牧食力,保持水体中浮游动物的生物量及优势地位.
白洋淀是中国华北平原最大和最典型的淡水湖泊湿地,近年来白洋淀流域内水体富营养化问题严重,各种理化方法和工程措施的实行虽耗资巨大但效果均不理想,白洋淀春夏季藻类生物量仍然较高,生物链调控湖泊富营养化技术因其经济、低耗、收效显著日益受到人们重视[4].作为水生食物链中不可缺少的组成成分[5],枝角类在水生态系统生物量方面也占据较大优势[6],因此本文选择白洋淀春、夏季主要浮游动物蚤状溞(Daphniapulex)和老年低额溞(Simocephalusvetulus)作为消费者,以导致白洋淀富营养化的优势藻类蛋白核小球藻(Chlorellapyrenoidosa)和四尾栅藻(Scenedesmusquadricanda)为生产者,研究枝角类对藻类的摄食作用,为生态系统生物链调控富营养化的理论及技术体系研究奠定基础,以充分发挥水生态系统对内源营养负荷和富营养化的综合调控功能.
1 材料与方法
1.1 枝角类
老年低额溞(S.vetulus)和蚤状溞(D.pulex)采自河北安新白洋淀,实验室镜检分离,空腹培养24h后用于实验.
1.2 藻类
蛋白核小球藻(C.pyrenoidosa)藻种购自中国科学院水生生物研究所淡水藻种库,锥形瓶灭菌后加入100mL SE培养基[7],接入藻种,光照培养箱(PGX-250C)中培养,温度(25±1)℃,光周期12L/12D,定期更换培养基,使藻处于对数生长期.四尾栅藻(S.quadricanda)采自白洋淀,分离后在实验室培养,培养条件同上.
1.3 实验方法
1.3.1 老年低额溞摄食单一藻类实验
取对数生长期的藻液,镜检计数藻液密度;配制不同生物量的藻液,每个实验瓶内放置10只活力较强的蚤状溞,实验分四尾栅藻、蛋白核小球藻2组,每组设4个密度梯度,每个梯度设3个重复,将实验瓶置于暗室中,温度控制在(25±1)℃;24h后,镜检直接计数各容器中的藻液密度,每个样品计数3片,镜检中测量藻类体积,参考“淡水浮游生物调查技术规范(SC/T 9402—2010)”换算为藻类生物量.
1.3.2 老年低额溞摄食混合藻类实验
由上述2种藻类按1∶1的生物量配成混合藻液,实验步骤同上.
1.3.3 蚤状溞摄食实验
蚤状溞的摄食实验,也分为单一藻类和混合藻类2部分.实验步骤同老年低额溞.
1.3.4 计算方法
选择指数的计算公式[8]
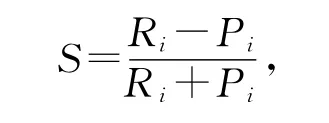
其中:S——选择指数;Pi——混合藻液中某种藻类所占的百分比;Ri——被枝角类摄食的藻类占全部藻类的比例.
摄食速度和滤水速度的计算公式[8]
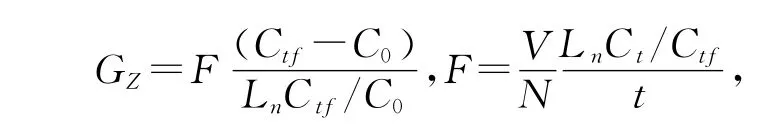
其中:Gz——摄食速度(μg/(溞·h));F——滤水速度(mL/(溞·h));N——放入该水体中的枝角类总数;V——实验瓶水的体积;C0——藻类密度;Ctf——t时投放枝角类的藻类密度;Ct——t时后未投放枝角类的藻类密度.
1.4 数据分析
采用SPSS11.5统计软件对数据进行单因素方差分析(One-Way ANOVA),采用LSD多重比较检验各参数间的差异.
2 结果与分析
2.1 老年低额溞摄食藻类
2.1.1 老年低额溞对蛋白核小球藻的摄食
表1所示为老年低额溞对蛋白核小球藻的摄食结果.在藻类生物量3.34~35.25mg/L内,老年低额溞的滤水速度与藻类生物量密切相关,水中藻类生物量的增加使得老年低额溞的滤水速度显著降低,但老年低额溞的摄食速度则随藻类生物量增加而显著升高,当蛋白核小球藻的生物量最高时(35.25mg/L),老年低额溞的摄食速度达到最大值2.50μg/(溞·h);而老年低额溞24h内对蛋白核小球藻的去除率达到最大值70.91%时,蛋白核小球藻的生物量为6.25mg/L,之后随着藻类生物量的增加,老年低额溞24h内对蛋白核小球藻的去除率显著降低.
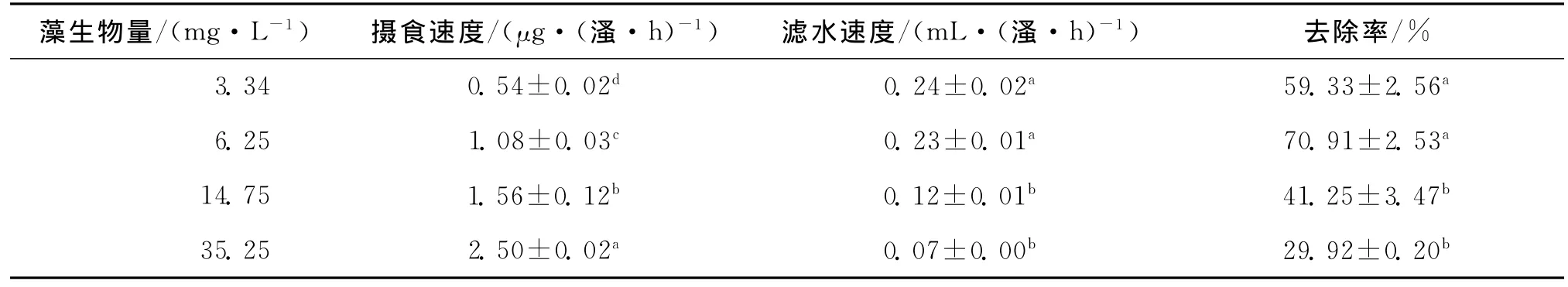
表1 老年低额溞对蛋白核小球藻的牧食力Tab.1 Grazing ability of Simocephalus vetulus on Chlorella pyrenoidosa
2.1.2 老年低额溞对四尾栅藻的摄食
表2为老年低额溞对不同生物量四尾栅藻的摄食结果,水中藻类生物量的增加降低了老年低额溞的滤水速度,而老年低额溞的摄食速度则随水中藻类生物量的升高而增加,当藻类生物量为35.00mg/L时老年低额溞的摄食速度达到最高,24h内老年低额溞对四尾栅藻的去除率随着藻类生物量的升高也由开始的最大值40.62%显著下降到12.66%.
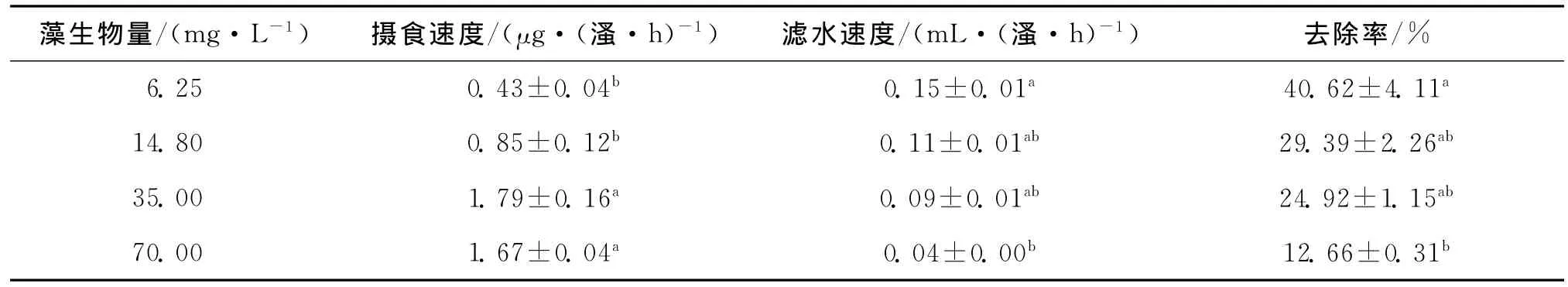
表2 老年低额溞对四尾栅藻的牧食力Tab.2 Grazing ability of Simocephallus vetulus on Scenedesmus quadricanda
2.1.3 老年低额溞对混合藻类的摄食
老年低额溞对含2种藻类的混合藻的摄食差异性如表3所示,在混合藻液中,老年低额溞对2种藻的摄食速度、滤水速度无明显差异;老年低额溞对四尾栅藻的选择指数显著大于其对蛋白核小球藻的选择指数(P<0.05),说明水中2种藻共存时老年低额溞明显优先选择四尾栅藻.

表3 老年低额溞对混合藻液中不同藻的牧食力Tab.3 Grazing ability of Simocephalus vetulus on different species of algae in mixed-species waters
2.2 蚤状溞对藻类的摄食
2.2.1 蚤状溞对蛋白核小球藻的摄食
蚤状溞对蛋白核小球藻的摄食结果如表4所示,在实验藻类生物量范围内,蛋白核小球藻生物量最低为3.34mg/L时,蚤状溞对藻类的摄食速度最低,而滤水速度和去除率最高,随着蛋白核小球藻生物量的增加,蚤状溞对藻类的滤水速度降低,摄食速度则升高,24h内蚤状溞对蛋白核小球藻的去除率由开始的最大值50.39%显著降低到8.49%.

表4 蚤状溞对蛋白核小球藻的牧食力Tab.4 Grazing ability of Daphnia pulex on Chlorella pyrenoidosa
2.2.2 蚤状溞对四尾栅藻的摄食
蚤状溞对四尾栅藻的摄食结果如表5所示,随着四尾栅藻生物量的增加,蚤状溞对四尾栅藻的滤水速度和去除率均显著降低,当四尾栅藻的生物量为6.25mg/L时,蚤状溞的滤水速度达到最大0.11mL/(溞·h),同时24h内对四尾栅藻的去除率也达到最大29.23%;四尾栅藻的生物量达到70.00mg/L时,蚤状溞的去除率和滤水速度显著下降为8.75%和0.03mL/(溞·h);蚤状溞对四尾栅藻的摄食速度则随着藻类生物量的增加显著增大.

表5 蚤状溞对四尾栅藻的牧食力Tab.5 Grazing ability of Daphnia pulex on Scenedesmus quadricanda
2.2.3 蚤状溞对混合藻类的摄食
蚤状溞对蛋白核小球藻和四尾栅藻的混合藻摄食差异性如表6所示,在混合藻液中,蚤状溞对四尾栅藻的摄食速度大于蛋白核小球藻,但其滤水速度则小于蛋白核小球藻,两者之间的差异均不显著(P>0.05);蚤状溞对四尾栅藻的选择指数显著大于其对蛋白核小球藻的选择指数(P<0.05),说明相对于蛋白核小球藻,蚤状溞更优先摄食四尾栅藻.

表6 蚤状溞对混合藻液中不同藻类的牧食力Tab.6 Grazing ability of Daphnia pulex on different species of algae in mixed-species waters
3 讨论
3.1 不同枝角类对蛋白核小球藻的摄食
藻类水华的控制以生物操纵最具前景,而浮游动物的摄食作用也是藻类水华控制的最常用手段.作为水域生态系统重要组成部分的枝角类,在水生生态系统生物生产力中占有重要的位置[8].不同种类的枝角类对藻类的控制作用具有明显差异,同时也与水体中藻类生物量密切相关.
本实验中,老年低额溞对于蛋白核小球藻的滤水速度、摄食速度和去除率均大于蚤状溞.2种枝角类的滤水速度均随蛋白核小球藻生物量的上升而显著降低,当蛋白核小球藻的生物量达到一定水平后蚤状溞的摄食速度开始下降,说明蚤状溞对蛋白核小球藻的摄食具有一定的饱和密度,韩士群等[9]研究发现当水体中藻类生物量超过一定值时,长肢秀体溞(Diaphanosamaleuchtenbergianum)的摄食速度趋于下降,而程汉良等[10]对多刺裸腹溞摄食强度的研究也具有相似结论;老年低额溞的摄食速度则随藻生物量的增加而上升,这与唐洪玉等[11]关于长刺溞(Diaphanosamalongispina)摄食的研究结果相似.蛋白核小球藻生物量较低时老年低额溞和蚤状溞对藻类的去除率较高,随着藻类生物量的增加去除率下降,但2种溞的摄食量却随藻生物量的增加而增加,说明枝角类取食能力的高低与其对藻类的去除率大小之间相关性不明显.综上所述,老年低额溞对于蛋白核小球藻生物量的控制效果优于蚤状溞.
3.2 不同枝角类对四尾栅藻的摄食
老年低额溞对于四尾栅藻的摄食速度、滤水速度和去除率均高于蚤状溞.随四尾栅藻生物量的上升,2种溞对四尾栅藻的摄食速度、滤水速度和去除率具有相同的变化趋势,去除率和滤水速度均降低,摄食速度则随四尾栅藻的生物量的增加先上升后下降,说明2种溞摄食四尾栅藻具有饱和密度,这与蛋白核小球藻不同,老年低额溞对蛋白核小球藻的摄食速度在实验藻类浓度范围内始终增加,说明老年低额溞对不同藻类的控制作用不同,同理,其去除率降低,也并不代表枝角类取食能力的降低.相同的藻类生物量水平下,老年低额溞的摄食速度、藻类去除率均高于蚤状溞,因此对于富营养化水体中的四尾栅藻利用老年低额溞进行生物调控更有效果.
3.3 不同枝角类对混合藻类的摄食
在单一藻类实验中,老年低额溞对蛋白核小球藻的摄食速度、滤水速度和去除率均高于四尾栅藻,但四尾栅藻和蛋白核小球藻共存条件下,老年低额溞对蛋白核小球藻和四尾栅藻的摄食速度、滤水速度无明显差异,但老年低额溞对四尾栅藻的选择指数显著大于蛋白核小球藻(P<0.05),说明对于以蛋白核小球藻和四尾栅藻为优势藻类的富营养化水体中,老年低额溞可能会优先利用四尾栅藻.蚤状溞对蛋白核小球藻和四尾栅藻的混合藻摄食差异性与老年低额溞结果相似,与蛋白核小球藻相比蚤状溞更倾向于选择四尾栅藻.白洋淀春季富营养化水体中蛋白核小球藻和四尾栅藻均为优势藻类,老年低额溞和蚤状溞对于富营养化水体藻类均有控制作用,但具体的控藻效果与藻类生物量、藻类种类组成差异等因素密切相关,应用生物链控藻治理白洋淀水体富营养化尚需进一步的理论探索与实践.
4 结论
1)老年低额溞和蚤状溞对四尾栅藻和蛋白核小球藻有一定的牧食力,而蚤状溞的去除率及摄食速度均低于老年低额溞,说明在白洋淀水体富营养化治理中,可以考虑利用这2种枝角类调控水体中藻类生物量,水体中藻类优势种为四尾栅藻和蛋白核小球藻时,老年低额溞的控藻效果要优于蚤状溞.
2)2种枝角类牧食力的大小受到藻类生物量的影响,老年低额溞对蛋白核小球藻的摄食速度随藻类生物量的增加而增加,实验中2种枝角类对四尾栅藻摄食均有饱和生物量,超过饱和生物量,枝角类的摄食速度下降.说明利用这2种枝角类进行生物控藻时,控藻效果受富营养化水体中藻类生物量的影响.
[1] 全为民,沈新强,严力蛟.富营养化水体生物净化效应的研究进展[J].应用生态学报,2003,14(11):2057-2061.QUAN Weimin,SHEN Xinqiang,YAN Lijiao.Advance in research of biological purification of eutrophic water body[J].Chinese Journal of Applied Ecology,2003,14(11):2057-2061.
[2] PACE M L.Zooplankton community structure,but not biomass,influence the phosphorus chlorophylla relationship[J].Can J Fish Aquat Sci,1984,41:1089-1096.
[3] BROOKS J L,DODSON S I.Predation,body size and the composition of plankton[J].Science,1965,150:28-35.
[4] 王军霞,张亚娟,刘存歧,等.生物链调控技术防治湖泊富营养化研究进展[J].湖北农业科学,2011,50(9):1736-1739.WANG Junxia,ZHANG Yajuan,LIU Cunqi,et al.Progress of biological chain regulation on lake eutrophication[J].Hubei Agricultural Science,2011,50(9):1736-1739.
[5] DOWNING J A,RIGLER F H.A manual on methods for the assessment of secondary production in fresh waters IBP Hand Book 17[M].Oxford:Blackwell Sci Publications,1984:336-412.
[6] 邓道贵,靳英,柳明,等.温度和食物浓度对老年低额溞生长与生殖的影响[J].水生生物学报,2006,30(3):298-303.DENG Daogui,JIN Ying,LIU Ming,et al.Effect of temperature and food concentration on the growth and reproduction ofSimocephalusvetulus[J].Acta Hydrobiologica Sinaca,2006,30(3):298-303.
[7] 柴民伟,石福臣,马妍,等.药用植物浸提液抑制蛋白核小球藻生长的化感效应[J].生态学报,2010,30(18):4960-4966.CHAI Minwei,SHI Fuchen,MA Yan,et al.The allelopathic and inhibitive effects of extracts from medicinal plants on the growth ofChlorellapyrenoidosa[J].Acta Ecologica Sinica,2010,30(18):4960-4966.
[8] DRENNER R W,HAMBRIGHT K D.Biomanipulation of fish assemblages as a lake restoration technique[J].Arch Hydrobiol,1999,146(2):129-165.
[9] 韩士群,严少华,范成新,等.长肢秀体溞对富营养化水体藻类的生物操纵[J].江苏农业学报,2006,22(1):81-85.HAN Shiqun,YAN Shaohua,FAN Chenxin,et al.Biomanipulation of algae in eutrophicated waters byDiaphanosoma leuchtenbergianumfischer[J].Jiangsu Journal of Agricultural Sciences,2006,22(1):81-85.
[10] 程汉良,王策.多刺裸腹藻摄食强度的研究[J].中国水产科学,1999,6(3):104-106.CHENG Hanliang,WANG Ce.A study on filtering intensity ofMoinamacrocoapa[J].Journal of Fishery Sciences of China,1999,6(3):104-106.
[11] 唐洪玉,周小河,姚维志.长刺溞对两种单细胞藻类牧食力研究[J].西南农业大学学报,2001,23(4):329-332.TANG Hongyu,ZHOU Xiaohe,YAO Weizhi.A study on the grazing ability ofDaphnialongispinaon chlorella and navicula[J].Journal of Southwest Agricultural University,2001,23(4):329-332.
猜你喜欢
西南农业学报(2022年8期)2022-10-28
中国油脂(2020年8期)2020-08-12
生态学报(2020年19期)2020-02-20
陶瓷学报(2019年5期)2019-01-12
绿色科技(2018年20期)2018-12-19
吉林农业(2018年24期)2018-12-18
生物医学工程研究(2018年1期)2018-10-18
中国水产(2018年9期)2018-09-28
河北渔业(2018年8期)2018-01-17
环境污染与防治(2016年9期)2016-03-13
