
氮肥和连续夜间低温对珙桐幼苗叶片光合特性的影响1)
2015-03-10 03:51丁坤元刘艳红
东北林业大学学报 2015年3期
丁坤元 刘艳红
(北京林业大学,北京,100083)
氮肥和连续夜间低温对珙桐幼苗叶片光合特性的影响1)
丁坤元 刘艳红
(北京林业大学,北京,100083)
通过盆栽试验,对2年生珙桐幼苗施加不同量氮肥,并对其进行连续夜间低温处理,分析珙桐幼苗净光合速率、气孔导度、蒸腾速率、叶绿素质量分数,对连续夜间低温的响应以及氮肥和低温的交互作用对上述指标的影响。结果表明:①不论施加氮肥多少,珙桐幼苗的各项指标都有一定增加,但均没有显著提高(P>0.05)。②连续夜间低温处理过程中,珙桐幼苗的净光合速率、气孔导度、蒸腾速率、总叶绿素质量分数、叶绿素b质量分数,均有显著的下降(P<0.05),且都在处理后期及恢复期出现了适应性回升,但均没有恢复到处理前水平,与处理前差异显著(P<0.05),而叶绿素a质量分数变化不明显。③除气孔导度外,氮肥和低温交互作用对珙桐幼苗光合特性影响并不显著(P>0.05)。
珙桐;氮肥;连续夜间低温;光合特性
We used different amounts of nitrogen fertilizer pot experiment and continuous nighttime low temperature treatment for two-year-old seedlings ofDavidiainvolucratato study the effect of continuous nighttime low temperature on photosynthesis ofD.involucrataseedlings, and determine whether the increase of nitrogen fertilizer could develop its cold resistance. Then, we studied the impact ofD.involucrataon net photosynthetic rate, stomatal conductance, transpiration rate and chlorophyll content continuous nighttime low temperature and cryogenic nitrogen response and interaction of these indicators. No matter how much fertilizer was applied, the indicators ofD.involucrataseedlings increased, but there was no significant increase (P>0.05). Net photosynthetic rate, stomatal conductance, transpiration rate, total chlorophyll content and chlorophyll b content ofD.involucrataseedlings in continuous nighttime low temperatures process were significantly decreased (P>0.05). These indicators rebounded in late treatment and recovery, but could not recover to the level before treatment with significant difference (P>0.05) compared to that in the pre-treatment. But the chlorophyll a content had not significant difference. Except stomatal conductance givers, nitrogenous fertilizer and low interaction on photosynthetic characteristics ofD.involucrataseedlings were not significant (P>0.05). There were some responses for the seedlings to adapt low temperature.
珙桐(DavidiainvolucrataBaill),是我国特有的珙桐科单型属珍稀濒危植物,为我国的一级重点濒危保护植物。因其花形酷似白鸽,又名“ 鸽子树 ”,已成为世界范围内的著名观赏树种[1-4]。珙桐是第三纪古热带植物区系的孑遗植物,曾于白垩纪和第三纪时期广泛分布于世界许多地区,但在第四纪冰川时期相继灭绝,目前只在我国南方的一些地区有少量幸存,因此,也有“活化石”之称[5]。由于生境变化,滥砍滥伐,幼苗破坏等自然和人为原因,目前珙桐的数量还在持续减少,分布范围也日趋缩小,面临灭绝的危险。
我国自20世纪70年代就已经开始了对珙桐的研究和保护。随着研究的发展,对珙桐的保护已不再局限于原地保护[6],北京市早在20世纪80年代就开始对珙桐进行引进迁地保护。温度、光照、湿度等环境因素对迁地保护有至关重要的影响,已有学者探究了珙桐对低温的响应情况[7],但并没有对珙桐的低温适应策略及增加其抗寒性的措施进行探讨。近几年对于珙桐北迁的研究较少,主要集中在对光强以及水分的适应研究[8-9],模拟北方大温差的夜间低温研究以及使用施肥方式增加珙桐幼苗抗寒性的实验还没有被尝试。本试验运用控制变量的方法,分析夜间连续低温以及土壤氮素对珙桐叶绿素质量分数、光合作用等的影响,为珙桐的引种保护及驯化提供科学依据,也为探讨珙桐濒危的原因提供理论基础。
1 材料与方法
1.1 材料
供试材料为2013年4月从湖南购进的2年生珙桐幼苗,栽培于北京林业大学苗圃温室,缓苗2个月。栽培容器为内径25 cm、深20 cm的塑料盆,土壤为温室苗圃土壤,每盆装土2.5 kg。缓苗过程中充分供水,缓苗期间温室湿度60%~70%,温度15~20 ℃。
1.2 试验处理
施肥处理:2013年5月下旬选取大小、长势基本一致的健壮幼苗120株进行施肥处理。施肥试验设置低氮处理组(L)、高氮处理组(H)和对照组(CK);低氮处理组施肥量每盆400 mg,高氮处理组施肥量每盆800 mg[8],对照组不施肥,每组40盆。试验用肥料为尿素。施加氮肥采用追肥法分两次进行,每次间隔15 d,两次施加等量氮肥。
夜间连续低温处理:鉴于北京地区春夏季节的昼夜温差远高于珙桐原产地,本实验设置了极端的夜间低温情况。2013年6月开始,每天18:00将三种处理,每种20盆幼苗移入人工气候箱(宁波江南仪器厂RXZ-0158)中,分别记为低温对照(D),低温低氮(DL),低温高氮(DH),设定温度4 ℃,湿度80%[9]。至第2天早上8:00将培养箱中的幼苗移回室外进行测定采样。连续重复5 d。第6天不进行低温处理,自然条件下恢复一天。每天取样后的样品,不再进行接下来的处理。
1.3 测定指标及方法
光合作用参数:光合和蒸腾速率等使用Li-COR公司的Li-6400便携式光合作用测定系统测定。测量时采用人工光源控制光强,室光照强度为600 μmol·m-2·s-1[9],共测定低温处理前,低温处理每天以及恢复1 d后的数据。测定时间为每天白天上午9:00—11:00,每个处理随机选取3盆幼苗,测量幼苗中上部无损伤、发育健康的叶片,每株测量3片叶片。
叶绿素质量分数:每次光合测定后,对每日测定的植株进行采样,采样叶片为测定光合时的3片叶片。用打孔器在所取叶片上取叶圆片,每片取10个圆片并立即测定叶绿素质量分数。叶绿素质量分数测定方法采用丙酮-分光光度计法测定[10]。
1.4 数据整理及分析
实验数据用 Excel 2007及SAS 9.0软件进行统计分析并作表。
2 结果与分析
2.1 连续夜间低温对不同处理珙桐幼苗叶绿素质量分数的影响
2.1.1 连续夜间低温期间珙桐幼苗总叶绿素质量分数变化
珙桐幼苗的总叶绿素质量分数在低温处理后开始呈明显下降趋势,且在处理后期出现了回升的趋势。尤其是施用过氮肥的处理,下降较早较快且回升趋势出现较早。低温下各处理的总叶绿素质量分数都与处理前存在显著的下降(P<0.05),并且在低温后期及缓和期也没能恢复到处理前水平,与处理前仍然存在显著差异(P<0.05)(见表1)。

表1 连续低温条件下不同处理珙桐幼苗总叶绿素质量分数 mg·g-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。
2.1.2 连续夜间低温期间珙桐幼苗叶绿素a、b质量分数的变化
由表2可知,叶绿素a的质量分数各种处理中没有显著变化;叶绿素b的质量分数下降趋势比总叶绿素质量分数较为缓和(见表3)。两个施肥低温处理在第3天才与处理前水平出现显著差异(P<0.05),而单独低温处理则在第4天才出现显著差异(P<0.05)。但施肥处理也在低温后期较快的出现了回升趋势,而单独低温处理则在低温结束后才开始恢复,三者叶绿素b的质量分数均没能恢复至处理前的水平。
2.1.3 连续夜间低温期间珙桐幼苗叶绿素a/b变化
由表4可知,由于叶绿素b的质量分数的变化,低温处理过程中珙桐幼苗叶绿素a/b也发生了明显的变化。低温处理第3天开始,两个施肥处理的比值上升至与前一天产生了显著差异(P<0.05)。而单独低温处理则在第4天才出现这种显著差异(P<0.05),该差异直至恢复期也一直存在,虽然有所恢复但并没能恢复至处理前水平。
总体看来,对珙桐使用氮肥,不论施肥量较多(H)还是较少(L),都没有对珙桐幼苗的叶绿素各项指标产生明显的影响(P>0.05)。

表2 连续低温条件下不同处理珙桐幼苗叶绿素a质量分数 mg·g-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。
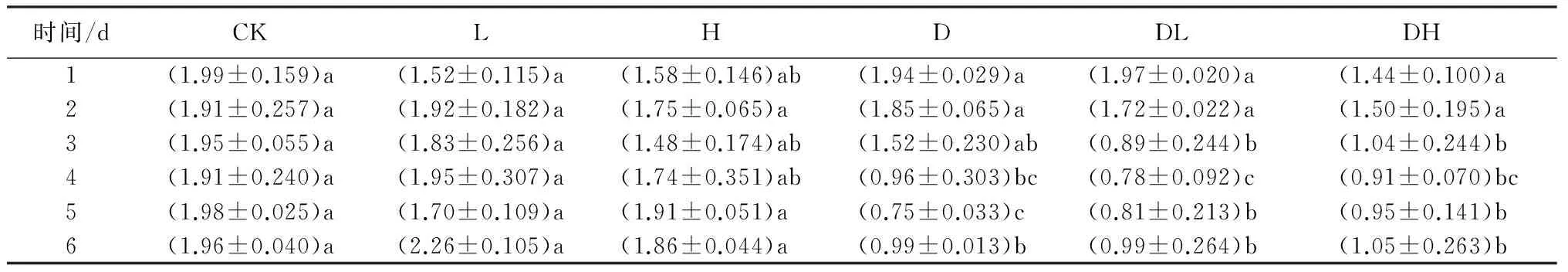
表3 连续低温条件下不同处理珙桐幼苗叶绿素b质量分数 mg·g-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。

表4 连续低温条件下不同处理珙桐幼苗叶绿素a/b值
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。
2.2 连续夜间低温对不同处理珙桐幼苗光合参数的影响
2.2.1 净光合速率变化
由表5可知,低温处理开始后,珙桐幼苗的净光合速率出现大幅度的下降,且均与处理前出现显著差异(P<0.05)。3个处理的净光合速率变化趋势较为一致,均随着夜间低温处理时间的增加而继续降低,且又与处理的第1天也存在了显著差异(P<0.05)。低温第3天,3个处理净光合速率出现了小幅度回升,之后继续下降,低温进行至最后1天,净光合速率大幅度回升,且都与处理的第2、第3和第4天呈显著差异(P<0.05)。停止低温处理后,净光合速率继续回升,数值均超过处理的第1天且差异显著(P<0.05),但没有达到处理前水平。

表5 连续低温条件下不同处理珙桐幼苗净光合速率 μmol·m-2·s-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。
2.2.2 气孔导度变化
由表6可知,连续低温处理下幼苗的气孔导度,第1天即开始下降并与处理前差异显著(P<0.05)。第2天和第3天,气孔导度继续下降,均与第1天存在显著差异。连续夜间低温处理的第4天,单独低温处理组和低氮肥处理组气孔导度开始回升,高氮肥处理在第5天气孔导度开始回升,但都没有与低温的前几天(第1天除外)呈显著差异(P>0.05)。停止低温处理后的恢复期,三组幼苗气孔导度开始继续回升,但都还保持在处理第1天水平,与处理前差异显著(P<0.05)。
2.2.3 蒸腾速率变化
由表7可知,低温过程中幼苗的蒸腾速率变化也呈下降趋势。单独低温处理在低温的第1天即与处理前出现了显著差异(P<0.05),第2天速率继续下降,以第1天的差异显著(P<0.05),第3天有所回升,基本达到第1天的水平,而第4天又下降回第2天水平。处理的最后1天蒸腾速率再次回升,其数值保持至处理结束后恢复期,但并没有恢复到处理前水平。施加氮肥的两个处理在低温过程中蒸腾速率下降幅度较大,均在第2天出现最小值,与前一天差异显著(P<0.05)。至低温处理结束及恢复期,蒸腾速率依旧保持在低温处理第3天的水平,没有再出现新的变化。

表6 连续低温条件下不同处理珙桐幼苗气孔导度 mol·m-2·s-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。

表7 连续低温条件下不同处理珙桐幼苗蒸腾速率 mmol·m-2·s-1
注:表中数值为“平均值±标准差”;同列不同小写字母表示在0.05水平差异显著,同列相同小写字母表示差异不显著(P>0.05)。
总体来看,珙桐幼苗在施加氮肥处理后,幼苗的净光合速率、气孔导度以及蒸腾速率都有了小幅度的提高,但并没有对照组呈现显著地差异(P>0.05)。施加少量氮肥的处理(L)和施加大量氮肥的处理(H)在这三个指标上也没有出现明显的区别。
2.3 低温与氮肥交互作用对珙桐幼苗叶绿素质量分数及光合特效的影响
由表8可知,连续低温和氮肥条件下,各处理的净光合速率、蒸腾速率、总叶绿素质量分数、叶绿素a质量分数、叶绿素b质量分数都没有显著的交互作用(P<0.05),仅气孔导度一项指标在处理的第2天(气孔导度下降过程)和处理结束的恢复期(气孔导度回升过程)出现了显著的交互作用(P<0.05)。

表8 低温与氮肥交互作用对珙桐幼苗的影响
注:表中的数值为该指标当天低温及肥料交互作用的F值,*表示交互作用显著(P<0.05)。
低温处理过程中,交互作用比较明显的是气孔导度和蒸腾速率两个指标。在处理的第1天和第4天,蒸腾指标的交互作用虽然没有达到显著的水平,但数值上已经与其他几天有较大差距;而气孔导度在第2天和恢复期则达到了显著水平。其他指标则是距离显著地交互作用差距极大。
3 讨论
3.1 氮肥对珙桐幼苗光合参数及叶绿素质量分数的影响
氮素是植物生长重要的化学元素。叶片对土壤氮素质量分数的变化会有直接的反应,叶片中的叶绿素和部分酶类活性都会因氮素质量分数变化受到直接的影响,从而影响叶片的光合效率。已有大量的实验证明,碳3植物和碳4植物的叶氮质量分数与其净光合速率呈正相关关系[11-14],这与本实验结果吻合。但在幼苗施加氮肥的过程中,施加量增加并没有显著提高叶片净光合速率,可能是因为氮素以叶绿体中叶绿素的形式存在于植物中,过多的氮肥同时增加了叶片中叶绿素的质量分数,也同时提升了氮素的代谢。光合中ATP和NADPH的消耗以及呼吸产物的消耗,这些因素共同限制了净光合速率的提升。叶片中氮素质量分数与叶绿体中叶绿素质量分数也有一定的相关性[11]。氮元素缺乏对叶绿体的组成有很大的影响,与充分供氮相比,缺氮植株中叶绿体较小,且组成叶绿体的基粒跟囊体所占的范围小,基质大,进而光合作用的效果会受到影响[15]。当幼苗受到连续低温胁迫时,施加氮肥的处理较对照组更加迅速的出现了反应,净光合速率以及叶绿素质量分数在下降后更早的出现了适应性恢复,这种现象可能是因为叶片中充足的氮素,保证了低温下叶绿素受损后能更快的合成,从而更快的提升净光合速率。另一方面也可能是充足的氮素可以使低温酶活性降低的情况下,为叶片提供更多的酶合成原料,使光合作用中重要的限制因子——酶在数量上得到一定保证。有研究表明,在一定范围内,核酮糖-1,5-二磷酸梭化酶/加氧酶质量分数与叶片含氮量之间相关,核酮糖-1,5-二磷酸梭化酶/加氧酶的质量分数随叶片中氮的质量分数增加而增加相[16]。由此推断,通过酶的变化,幼苗低温条件下更快的出现了净光合速率及叶绿素质量分数的恢复。施加氮肥虽并不能很有效的提高幼苗的抗寒性,但对幼苗抗寒有一定帮助。
3.2 低温对珙桐幼苗光合参数及叶绿素的影响
珙桐对夜间低温的反应较为敏感,出现如芒果等植物的迟滞性[17]。处理的第2天光合指标及叶绿素质量分数与对照及实验前显著降低。由于处理及恢复期间净光合速率、气孔导度及蒸腾速率变化规律大体一致,推测珙桐受夜间低温胁迫后,限制气孔增大,导致日间气孔导度降低,气孔对CO2更加敏感并且引起质量分数的变化,从而导致净光合速率降低,蒸腾速率下降。而这种调剂气孔减少水分蒸腾,维持水分平衡的变化常存在于耐低温植物中[18-19],由此也可以推断珙桐可能是一种耐低温的植物。此外,有研究发现低温对植物非气孔限制也有极大影响[20]。夜间低温能干扰蔗糖磷酸合成酶和硝酸还原酶活性和叶绿素结合蛋白及核酮糖1,5-二磷酸羧化酶活化酶基因表达的昼夜节律性[21-22],且能抑制淀粉的转移[23],从而减少细胞质返回到叶绿体的无机磷[24],最终引起光合作用终产物的反馈抑制。且低温会明显降低耐低温植物叶片中的叶绿素尤其是叶绿素b的质量分数[25-26]。就本实验而言,连续低温条件下,珙桐幼苗叶绿素质量分数的迅速下降与上述研究结果较为一致,净光合速率的下降是气孔变化与光合反馈抑制的共同作用结果。
3.3 夜间低温及氮肥交互作用
在本次实验的过程中,除了叶片的气孔导度外,低温和氮肥并没有在幼苗叶片的其他光合及叶绿素指标上表现出显著的交互作用。施加氮肥其实并不能很好的提高珙桐幼苗的瞬时抗寒性,而只是在幼苗生长的过程中有一定作用。另一方面,珙桐如果可以被定义为一种耐低温植物,在遭受到低温胁迫的时候,幼苗能够迅速的做出生理响应,施加氮肥可以在一定程度上加速幼苗的保护反应以及适应性恢复,这也在一定程度上有利于珙桐的抗寒性。已有研究表明珙桐在低温胁迫条件下其脯氨酸质量分数会有一定累计以提高自身抗寒能力[7]。而大量实验也证明,施加氮肥可以提高植物体内游离脯氨酸的质量分数[27-28],并在一定程度上提高植物的抗逆性及适应性[29]。
夜间低温严重影响珙桐幼苗的光合特性,但幼苗又对连续低温存在一定适应性恢复。通过生理指标的进一步测定,可以更深入的研究氮肥对珙桐抗寒性的影响。施加氮肥对珙桐幼苗抗寒性的影响有待进一步研究和探讨。
[1] 胡进耀,苏智先,黎云祥.珙桐生物学研究进展[J].中国野生植物资源,2003,22(4):15-19.
[2] 肖开煌,苏智先,黎云祥,等.珙桐群落中珙桐地位及其土壤特征的相关性[J].西华师范大学学报:自然科学版,2006,27(4):403-407.
[3] 张亚爽,苏智先,胡进耀.四川卧龙自然保护区珙桐种群的空间分布格局[J].云南植物研究,2005,27(4):395-402.
[4] 周良,苏智先,胡进耀,等.卧龙自然保护区白泥岗珙桐群落的物种多样性研究[J].四川林业科技,2004,25(1):31-35.
[5] 张家勋,李俊清,周保顺,等.珙桐的天然分布和人工引种分析[J].北京林业大学学报,1995,17(1):25-30.
[6] 吴刚,肖寒,李静,等.珍稀濒危植物珙桐的生存与人为活动的关系[J].应用生态学报,2000,11(4):493-496.
[7] 彭红丽,苏智先.低温胁迫对珙桐幼苗的抗寒性生理生化指标的影响[J].汉中师范学院学报:自然科学版,2004,22(2):50-53.
[8] 宋天宇,刘艳红,姚毅.土壤氮素与光照对盆栽珙桐幼苗光合参数的影响[J].生态科学,2011,30(3):243-249.
[9] 王宁宁,胡增辉,沈应柏.珙桐苗木叶片光合特性对土壤干旱胁迫的响应[J].西北植物学报,2011,31(1):101-108.
[10] 李合生.植物生理生化实验原理与技术[M].北京:高等教育出版社,2000.
[11] Evans J R. Photosynthesis and nitrogen relationships in leaves of C3 plants[J]. Oecologia,1989,78(1):9-19.
[12] Poorter H, Van der Werf A, Lambers H, et al. Inherent variation in plant growth Physiological mechanisms and Ecological Consequences[M]. Leiden: Backhuys Publishers,1998.
[13] Lauerer M, Saftic D, Quick W P, et al. Decreased ribulose-1,5-bisphosphate carboxylase-oxygenase in transgenic tobacco transformed with “antisense” rbcS[J]. Planta,1993,190(1):1-9.
[14] Hiremath A J. Photosynthetic nutrient-use efficiency in three fast-growing tropical trees with differing leaf longevities[J]. Tree physiology,2000,20(14):937-944.
[15] Lawlor D W. Carbon and nitrogen assimilation in relation to yield: mechanisms are the key to understanding production systems[J]. Journal of Experimental Botany,2002,53:773-787.
[16] Osaki M, Morikawa K, Matsumoto M, et al. Productivity of high-yielding-crops[J]. Soil Science and Plant Nutrition,1993,39(3):399-408.
[17] Allen D J, Ratner K, Giller Y E, et al. An overnight chill induces a delayed inhibition of photosynthesis at midday in mango (MangiferaindicaL.)[J]. Journal of Experimental Botany,2000,51:1893-1902.
[18] Pardossi A, Vernieri P, Tognoni F. The involvement of ABA in ameliorating plant water status of Phaseolus vulgaris during chilling[J]. Plant Physiology,1992,100(3):1243-1250.
[19] LEE T M, Lur H S, Chu C. Role of abscisic acid in chilling tolerance of rice (OryzasativaL.) seedlings. I. Endogenous abscisic acid levels[J]. Plant, Cell & Environment,1993,16(5):481-490.
[20] Flexas J, Badger M, Chow W S, et al. Analysis of the relative increase in photosynthetic O2uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress[J]. Plant Physiology,1999,121(2):675-684.
[21] Jones T L, Tucker D E, Ort D R. Chilling delays circadian pattern of sucrose phosphate synthase and nitrate reductase activity in tomato[J]. Plant Physiology,1998,118(1):149-158.
[22] Martino-Catt S, Ort D R. Low temperature interrupts circadian regulation of transcriptional activity in chilling-sensitive plants[J]. Proceedings of the National Academy of Sciences,1992,89(9):3731-3735.
[23] Leegood R C, Edwards G E. Carbon metabolism and photorespiration: temperature dependence in relation to other environmental factors[M]. The Netherlands: Kluwer Academic Publishers,1996.
[24] Stitt M. Rising CO2levels and their potential significance for carbon flow in photosynthetic cells[J]. Plant, Cell & Environment,1991,14(8):741-762.
[25] 孙治强,张强,张惠梅.低温弱光对番茄叶绿素含量变化的影响[J].华北农学报,2005,20(1):82-85.
[26] 曾韶西,王以柔,刘鸿先.低温光照下与黄瓜子叶叶绿素降低有关的酶促反应[J].植物生理学报,1991,17(2):177-182.
[27] 庄飞.沙棘和刺槐叶片对土壤水分和氮素有效性的生理生化响应[D].杨凌:西北农林科技大学,2013.
[28] 邓斌,曾德慧.添加氮肥对沙地樟子松幼苗生物量分配与叶片生理特性的影响[J].生态学杂志,2006,25(11):1312-1317.
[29] 刘永安.在不同氮、钾施用量和遮阴条件下麻疯树幼苗对干旱的生理生态响应[D].雅安:四川农业大学,2011.
Effects of Nitrogen and Continuous Nighttime Low Temperature on Photosynthetic Characteristics ofDavidiainvolucrataLeaves
Ding Kunyuan, Liu Yanhong(Beijing Forestry University, Beijing 100083, P. R. China)/Journal of Northeast Forestry University,2015,43(3):56-61.
Davidiainvolucrata; Nitrogen; Continuous nighttime low temperatures; Photosynthetic characteristics
丁坤元,女,1989年4月生,北京林业大学林学院,硕士研究生。E-mail:dingkunyuan@163.com。
刘艳红,北京林业大学林学院,教授。E-mail:liuyh@bjfu.edu.cn。
2014年10月7日。
Q948.112
1)北京市重点学科项目“生态学”(2013-2014年度)。
责任编辑:王广建。
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
花卉(2020年16期)2020-08-22
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
现代园艺(2018年1期)2018-03-15
新课程·下旬(2017年7期)2017-08-14
中国新技术新产品(2015年22期)2015-12-21
中国铸造装备与技术(2015年5期)2015-12-10
西北林学院学报(2015年5期)2015-01-03
中国西部(2014年2期)2014-02-02
