
日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫BALB/c小鼠诱导免疫应答的动态研究
2015-05-09 01:03谭建蓉李文桂
中国人兽共患病学报 2015年9期
谭建蓉,李文桂,覃 婷
·实验研究·
日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫BALB/c小鼠诱导免疫应答的动态研究
谭建蓉1,李文桂2,覃 婷1
目的 动态观察日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫BALB/c小鼠后诱导的免疫应答,探讨该疫苗的免疫机制。方法 将日本血吸虫重组Bb(pGEX-Sj32)疫苗分别经口服(PO)和滴鼻(IN)途径免疫BALB/c小鼠,在免疫后0~20周,每隔2周每组随机剖杀4只小鼠,取脾并制备脾细胞悬液,用四甲基偶氮唑盐比色法(MTT)检测脾细胞增殖水平、流式细胞仪(FACsort)检测脾CD4+和CD8+T淋巴细胞亚群和脾细胞凋亡、双抗体夹心ELISA法检测血清及脾细胞培养上清液中细胞因子IL-10、IL-12和TNF-α水平和血清抗体IgG及其亚类、IgE和IgA的动态变化。结果 当SjAWA和ConA刺激或不刺激时,PO组和IN组脾细胞增殖水平均于免疫后6周达最高水平,分别于6周和8周达最高脾细胞凋亡发生率,脾CD4+T细胞亚群百分比分别在6周和8周达峰值,两组CD8+T细胞亚群百分比在免疫后2~20周升高不显著,PO组脾细胞因子IL-10、IL-12和TNF-α水平均分别于10周、6周和8周达峰值,IN组分别在10周、8周和8周达峰值,口服组血清抗体IgG、IgG1、IgG2a、IgG2b、IgG3和IgA水平均于8周达峰值,而IN组除IgG和IgA于8周达峰值外,其余均于12周达最高水平,IgE于14周达峰值,两组血清细胞因子IL-10、IL-12和TNF-α水平分别在免疫后(PO组)14、6和8周及(IN组)8、6和8周达最高水平。结论 该疫苗可诱导小鼠产生有效的免疫应答。
日本血吸虫;重组Bb(pGEX-Sj32)疫苗;免疫
随着对日本血吸虫基因组学研究的不断深入[1],选取日本血吸虫高免疫原性蛋白编码基因制备各种疫苗,用于防治日本血吸虫病已成为当前的研究热点。Sj(Schistosomajaponicum)32基因编码的32 kDa蛋白被证实为天冬酰胺肽链酶[2],是虫体消化宿主血红蛋白的关键酶之一,具有较好的免疫原性。已有研究表明利用该基因制备的血吸虫疫苗具有较好的免疫保护性,免疫小鼠后能得到一定的减虫减卵率[3-4]。为了克服现有日本血吸虫病疫苗的安全性差、有效性低等缺点,本文在成功构建日本血吸虫重组Bb(pGEX-Sj32)疫苗[5]的基础上,将其免疫小鼠,动态观察其诱导小鼠产生的免疫应答,旨在探索该疫苗的作用机制,为后续研究提供有意义的材料。
1 材料和方法
1.1 材料
1.1.1 实验试剂及仪器 日本血吸虫成虫抗原(SjAWA)、日本血吸虫Bb(pGEX-Sj32)疫苗由本室保存;羊抗小鼠HRP-IgG、IgG1、IgG2a、IgG2b、IgG3、IgE和IgA购自美国Southern Biotech公司;IL-10、IL-12和TNF-α检测试剂盒购自深圳欣博盛生物科技有限公司;四甲基偶氮唑盐(MTT)、二甲基亚砜(DMSO)购自重庆白萃生物;大鼠抗小鼠CD4+T和CD8+T细胞单抗购自北京邦定泰克;Annexin V-FITC试剂盒(FAK015)购自深圳欣博盛生物科技有限公司;刀豆蛋白A(ConA)购自美国Sigma;RPMI 1640培养液、10%胎牛血清、青霉素和链霉素购自上海生工;Elx800UV酶标仪(美国Bio-TEK);流式细胞仪FACSort(美国Becton Dickinson)。
1.1.2 实验动物及免疫方案 88只雌性BALB/c鼠,6~8周龄,体重(18±2) g,购自重庆医科大学实验动物中心。将其随机分为两组:口服免疫组(Per os ,PO)和滴鼻免疫组(intranasal,IN),PO组:经口服单次接种悬浮于100 μL MRS液体培养的重组Bb(pGEX-Sj32)疫苗1×108克隆形成单位(CFU);IN组:经滴鼻单次接种悬浮于10 μL MRS液体培养的重组Bb(pGEX-Sj32)疫苗1×105CFU。至实验结束无小鼠死亡。
1.2 方法
1.2.1 脾细胞分离及血清收集 在疫苗免疫后0、2、4、6、8、10、12、14、16、18和20周两组各随机剖杀4只小鼠,取眼球血,室温放置过夜,4 000 r/min离心10 min,收集血清,置-20 ℃保存备用;无菌取脾,制备成终浓度为5×109/L的脾细胞悬液,方法同文献[6]。
1.2.2 脾细胞增殖检测 将分离的脾细胞(浓度为5×109/L)接种于24孔细胞培养板,每份标本设3组:原液组(基础培养基)、SjAWA刺激组和ConA刺激组,分别加入1mL脾细胞悬液、1 mL脾细胞悬液+10 μg 的SjAWA和1 mL脾细胞悬液+10 μg 的ConA,SjAWA和ConA的终浓度均为10 g/L,最后于每孔加入10 μL的MTT(浓度为5 g/L),37 ℃、5% CO2培养18 h,弃去细胞上清液150 μL/孔,加入DMSO 150 μL/孔,吹打至甲臜充分溶解,将混匀的细胞转至96孔板(150 μL/孔),同时设3个空白孔为对照,酶标仪570 nm波长检测吸光度(A)值。
1.2.3 脾细胞凋亡检测 将制备的脾细胞悬液加入24孔细胞培养板(细胞密度为5×109/L),每份标本设2组:原液组和ConA刺激组,分别加入0.5 mL脾细胞悬液和0.5 mL脾细胞悬液加5 μg的ConA,在37 ℃、5%的CO2细胞培养箱中培养16~18 h,将培养的脾细胞转至1.5 mL无菌Ep管中,4 ℃预冷的PBS洗两次,2 500 r/min 离心5 min,弃上清,加入250 μL结合缓冲液重调细胞密度至1×1010/L,加5 μL的Annexin V-FITC及10 μg/μL的碘化丙锭溶液(PI)10 μL混匀,室温避光孵育15 min,流式细胞仪测定脾细胞的凋亡发生率。
1.2.4 脾细胞亚群检测 将细胞浓度为5×109/L的脾细胞悬液与1%的阻断液各200 μL混匀于1.5 mL EP管,置37 ℃孵育1 h;加1 mL PBS洗涤2次(方法同前),加入大鼠抗小鼠CD4+或CD8+亚群单抗5 μL,4 ℃放置30 min,加入1 mL PBS洗涤2次,离心并弃上清液,加入5 μL的兔抗大鼠FITC-IgG抗体混匀,4 ℃放置30 min,同上洗涤2次,离心后弃上清液,用FACsort流式细胞仪测定CD4+T细胞和CD8+T细胞亚群的百分比。
1.2.5 脾细胞因子的检测 将步骤1.2.1制备的脾细胞悬液加入24孔细胞培养板,每份标本设原液组(1 mL脾细胞悬液)、SjAWA刺激组(1 mL脾细胞悬液+10 μg的SjAWA)和ConA刺激组(1 mL脾细胞悬液+10 μg的ConA),37 ℃ 5%CO2培养48 h,收集脾细胞悬液至1.5 mL EP管,3 000 r/min 离心5 min,收集上清液,-20 ℃保存备用。按双抗体夹心ELISA试剂盒说明书检测TNF-α、IL-10和IL-12水平。
1.2.6 常规ELISA法检测血清抗体IgG及其亚类、IgE和IgA 棋盘滴定法确定最佳抗原包被浓度,每孔加入1 μL的SjAWA(1 μg/μL) 和200 μg抗原包被液包被96孔酶标板,置湿盒于4 ℃过夜;弃抗原液,以磷酸缓冲液-吐温20 (PBST)洗板3次,每次5 min,甩干,每孔加入100 μL封闭液,置湿盒于37 ℃封闭2 h;同上洗板并甩干后,加小鼠血清(15100稀释)100 μL,置湿盒于37℃反应1 h; 同上方法洗板3次,分别加入辣根过氧化物酶HRP (1∶5 000稀释)标记的羊抗小鼠IgG、IgG1、IgG2a、IgG2b、IgG3、IgE和IgA 100 μL,37 ℃湿浴30 min;同上洗板3次,甩干后,加入100 μL底物液(四甲基联苯胺,TMB ),37 ℃反应15 min,加入50 μL浓硫酸(2 mol/L)终止反应,用酶标仪测450 nm处吸光度(A)值。
1.2.7 双抗体夹心ELISA法检测血清IL-10、IL-12和TNF-α水平 分别按IL-10、IL-12和TNF-α试剂盒的说明书操作,检测细胞因子IL-10、IL-12和TNF-α水平;酶标仪测450 nm处吸光度(A)值;通过标准品A值制备标准曲线,据此计算各细胞因子浓度。
2 结 果
2.1 脾淋巴细胞增殖 当SjAWA和ConA刺激或不刺激时,PO组小鼠脾细胞增殖水平均在免疫后2~16周升高,IN组小鼠脾淋巴细胞增殖水平均在免疫后2~18周升高,两组均于免疫后6周达最高水平,与0周相比有统计学差异(P<0.01);同一时间点内原液组、SjAWA刺激和ConA刺激组比较,SjAWA与ConA能够明显促进脾细胞的增殖,ConA作用最强,与原液组相比具有统计学差异(P<0.01);不同免疫途径比较,IN组与PO组小鼠脾淋巴细胞增殖水平差异无统计学意义。(见表1)

表1 日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫小鼠脾细胞增殖的检测
Note: Compared with 0 week, *P<0.05, **P<0.01.
2.2 脾细胞凋亡发生率 PO组小鼠在未刺激和ConA刺激时脾细胞凋亡率均在免疫后2~14周显著升高,于6周达最高水平,在免疫后4~14周两组细胞凋亡发生率与0周相比显著升高(P<0.01);IN组小鼠在未刺激和ConA刺激时脾细胞凋亡率均在免疫后6~12周升高,于8周达峰值,与0周相比差异有统计学意义(P<0.05或P<0.01);两未刺激组脾细胞凋亡率均明显低于相应的ConA组(P<0.01);两免疫途径比较未见统计学差异。(见表2)

表2 日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫小鼠脾细胞凋亡的检测
Note: Compared with 0 week, *P<0.05, **P<0.01.
2.3 脾细胞亚群 PO组和IN组小鼠脾CD4+T细胞亚群百分比均在免疫后2~12周升高,分别在6周和8周达峰值,与0周相比差异有统计学意义(P<0.01);两组CD8+T细胞亚群百分比在免疫后2~20周缓慢升高,仅PO组6~8周、IN组2周和12周与0周相比有统计学差异(P<0.05或P<0.01),其余时间点均无统计学意义;两组间比较差异无统计学意义。(见图1)

aP<0.05,bP<0.01,compared with 0 week
图1 日本血吸虫重组Bb(pGEX-Sj32)疫苗免疫小鼠脾细胞亚群的检测
Fig.1 Splenocyte subsets from mice immunized with recombinant Bb(pGEX-Sj32) vaccine againstSchistosomajaponicum
2.4 脾细胞因子IL-10、IL-12和TNF-α水平的检测 原液、SjAWA和ConA刺激时,PO组小鼠脾细胞因子IL-10、IL-12和TNF-α水平均分别在2~18、2~14及2~14周升高,并分别于免疫后10、6和8周达最高水平,与0周相比差异有统计学意义(P<0.05或P<0.01);原液、SjAWA和ConA刺激时,IN组小鼠脾细胞因子IL-10、IL-12和TNF-α水平均分别在2~18、2~18及2~14周升高,并分别于免疫后10、8和8周达最高水平,与0周相比差异有统计学意义(P<0.05或P<0.01)。(见表3、表4)
2.5 血清抗体水平 口服免疫组小鼠血清中IgG、IgG2a、IgG2b、IgG3水平分别在免疫后4~20、2~16、2~20和2~20周升高,并均于8周达到峰值,且分别在6~16、4~12、6~12和6~10周与0周相比升高有统计学意义(P<0.05或P<0.01),IgG1、IgE和IgA水平仅分别在6~8、12~14和8~10周有统计学差异,其余时间点均未测出;滴鼻组IgG、IgG2a、IgG2b、IgG3水平分别在免疫后2~20、2~20、4~20和2~20周升高,除IgG于8周达峰值外其余均于12周达到峰值,且分别在4~14、4~14、10~14和10~12周与0周相比升高有统计学意义(P<0.05或P<0.01),IgG1、IgE和IgA水平仅分别在8~10、14~16和8~10周有统计学差异,其余时间点检测值为0。(见图2、图3)
2.6 血清细胞因子IL-10、IL-12和TNF-α水平 与0周相比,口服免疫组IL-10、IL-12和TNF-α水平均于免疫后2~20周升高,并分别在14、6和8周达最高水平,与0周相比有统计学意义(P<0.05或P<0.01);滴鼻免疫组分别于免疫后2~16、2~20和2~20周升高,并分别在8、6和8周达最高水平,与0周相比有统计学意义(P<0.05或P<0.01)。见表5。
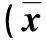
表3 日本血吸虫重组Bb(pGEX-Sj32)疫苗口服免疫小鼠脾细胞上清液IL-10、IL-12和TNF-α水平的检测
Note: Compared with 0 week,*P<0.05,**P<0.01.
表4 日本血吸虫重组Bb(pGEX-Sj32)疫苗滴鼻免疫小鼠脾细胞上清液IL-10、IL-12和TNF-α水平的检测
Note: Compared with 0 week, *P<0.05, **P<0.01.

图2 rBb(pGEX-Sj32)疫苗口服免疫小鼠血清抗体的动态变化
Fig.2 Dynamic changes on the levels of antibodies in sera of BALB/c mice orally immunized with rBb (pGEX-Sj32) vaccine

图3 rBb(pGEX-Sj32)疫苗滴鼻免疫小鼠血清抗体的动态变化
Fig.3 Dynamic changes on the levels of antibodies in sera of BALB/c mice intranasally immunized with rBb(pGEX-Sj32) vaccine
表5 rBb(pGEX-Sj32)疫苗免疫小鼠血清细胞因子IL-10、IL-12和TNF-α的动态变化
Note: Compared with 0 week, *P<0.05, **P<0.01.
3 讨 论
大量研究表明日本血吸虫疫苗经滴鼻和口服免疫小鼠后,在小鼠体内稳定表达32 kDa抗原蛋白,该抗原经免疫提呈细胞有效提呈,与MHC-II类分子结合成免疫复合物,被体内T淋巴细胞所识别,使其不断增殖分化为CD4+T淋巴细胞,而经分化成熟的CD4+T细胞进一步活化为辅助性T(Th)细胞,受抗原性质和局部免疫微环境等因素的影响,Th(Th1/Th2)细胞可分泌多种细胞因子引起和(或)加强体液及细胞免疫应答。研究表明Th1型细胞因子IL-2、TNF-α和IFN-γ具有活化T细胞和巨噬细胞的功能,介导细胞免疫应答,Th2型细胞因子IL-4、IL-5和IL-10可促进B细胞产生抗体介导体液免疫应答,两种机制共同作用加强免疫[7-9]。
机体发生免疫应答是体内各免疫细胞、特异性抗体及多种细胞因子共同调节的结果,细胞因子可调节免疫应答过程中细胞间相互作用、免疫细胞的生长和分化等,是一类重要的免疫活性分子。研究表明Th1型细胞因子IL-12和TNF-α具有重要的免疫学效应,IL-12可促进Th0细胞向Th1方向分化成熟,分泌细胞因子TNF-α和促进抗体IgG2a产生、抑制IgG1生成而形成Th1型免疫保护作用[10],Cheng等[11]研究发现IL-12能阻碍血吸虫雌虫生长,减少产卵,从而减轻由虫卵引起的病理性炎症反应和纤维化,以上研究表明IL-12是一类重要的免疫增强和保护因子。细胞因子TNF-α在血吸虫感染免疫中可通过募集多种粒细胞而引发炎症反应,能介导虫卵肉芽肿形成及杀伤童虫,大量研究表明多种血吸虫疫苗的免疫保护力都与其诱导机体产生高水平TNF-α有关[12-14]。而抗体具有凝集抗原、降低毒素水平、发挥体液免疫和辅助细胞免疫等重要作用,自然感染日本血吸虫后体内产生的IgG抗体及其亚类起到主要的抗炎杀虫作用,其机制与其增强NK及巨噬细胞吞噬能力、中和毒素、细胞毒效应和激活经典补体途径等作用相关[15]。Tu等[16]研究发现日本血吸虫pIRES-SjFABP-Sj26GST疫苗可诱导小鼠产生高水平IgG抗体,从而引发特异性体液免疫应答;Dai等[17]将日本血吸虫rAdV-SjTPI.opt疫苗分别经皮下及肌内注射免疫小鼠后检测到高水平IgG、IgG1和IgG2a抗体及多种Th1和Th2类细胞因子,认为该疫苗的保护作用与抗体介导的ADCC效应和细胞因子介导的细胞免疫应答有关;Wang等[18]对PAMAM-Lys/pJW4303-SjC23-mHSP70疫苗研究发现其能诱导产生高水平的IgG2a、IL-2和TNF而介导Th1型免疫应答发挥保护作用。本研究显示日本血吸虫重组Bb(pGEX-Sj32)疫苗可促进小鼠脾淋巴细胞早期即开始大量增殖,并向CD4+T淋巴细胞亚群优势分化,进而在免疫早中期分泌IL-12和TNF-α等Th1类细胞因子,在免疫后期则以分泌IL-10等Th2类细胞因子为主,共同发挥有效的细胞免疫应答;与此同时,某些细胞因子还可促进B细胞向浆细胞分化,加强抗体IgG及其亚类IgG1、IgG2a、IgG2b和IgG3产生,引发体液免疫应答,在抗日本血吸虫感染中发挥重要的作用。推测日本血吸虫重组Bb(pGEX-Sj32)疫苗表达的抗原分子主要与MHC-II类分子结合促进CD4+T细胞亚群升高,增殖活化为Th细胞分泌IL-12、IL-10和TNF-α等多种细胞因子,发挥免疫效应;并通过正反馈调节机制,CD4+T细胞不断增殖,使免疫效应得以强化并维持;同时某些细胞因子抑制CD8+T细胞亚群的分化,TS细胞相应减少,阻碍了其免疫下调作用,如此形成CD4+T细胞亚群分化优势;分化成熟的CD4+T细胞产生大量的Th1和Th2类细胞因子,这些细胞因子通过募集炎症细胞聚集、介导吞噬细胞吞噬等作用发挥强效的细胞免疫,同时以不同的方式(促进、抑制和类别转换)参与抗体的成熟和产生,从而发挥中和毒素、促进补体产生及连接杀伤细胞等体液免疫,如此建立细胞因子之间、细胞因子与抗体和免疫细胞之间相互作用的网络模式,形成该重组疫苗诱导的强效免疫保护机制。
研究表明诱导细胞凋亡是血吸虫产生病理损伤及免疫逃避的关键[19]。Chen等[20]研究发现曼氏血吸虫可分泌抗原物质上调T淋巴细胞表面FasL的表达,促进皮肤T淋巴细胞的凋亡,逃避机体对其产生的免疫应答,阻断和(或)干预免疫细胞凋亡有助于控制各炎症反应和病理损伤。本研究显示两组小鼠脾细胞凋亡率在免疫后早中期显著升高,分别在6周(PO组)和8周(IN组)达峰值,免疫后期细胞凋亡发生率则逐渐下降,可见rBb(pGEX-Sj32)疫苗对脾淋巴细胞凋亡具有抑制效应。推测早中期凋亡现象与疫苗诱导淋巴细胞大量增殖有关,为了提升自身免疫能力机体需要不断更新免疫细胞而启动反馈性凋亡反应,促进老的、免疫效力低下的淋巴细胞凋亡;另一方面机体需要维持稳定而高效的免疫力,增殖的效应性T细胞分泌凋亡抑制因子,抑制脾淋巴细胞凋亡。通过以上机制,疫苗的总效应表现为抑制脾淋巴细胞凋亡,使机体储备较多的免疫活性细胞,从而上调免疫应答。
[1]Chen Z, Wang SY, Han ZG. TheSchistosomajaponicumgenome reveals features of host-parasite interplay[J]. China Basic Sci, 2010, 12(3): 13-17. (in Chinese) 陈竺,王升跃,韩泽广.日本血吸虫全基因组测序完成[J].中国基础科学,2010,12(3):13-17.
[2]Ruppel A, Diesfeld HJ, Rother U. Immunoblot analysis ofSchistosomamansoniantigens with sera of schistosomiasis patients: diagnostic potential of an adult Schistosome polypeptide[J]. Clin Exp Immunol, 1985, 62(3): 499-506.
[3]Shi Y, Xie L, Li C, et al. DNA vaccine pCD-Sj32 and its efficacy of protective immunity against infection ofSchistosomajaponicum[J]. Chin Med J (Engl), 1999, 112 (8): 677-680.
[4]Shen DW, Luo JP, Peng SG, et al. Studies on immunoprotection in mice after immunization withSchistosomajaponicum31/32kDa recombinant protein[J]. Chin J Public Health, 2002, 18(2): 171-172. (in Chinese) 沈定文,罗金萍,彭胜国,等.日本血吸虫31/32kDa重组抗原对小鼠免疫保护性研究[J].中国公共卫生,2002,18(2):171-172.
[5]Tan JR, Li WG, Zhang L. Construction and identification of recombinant vaccineBifidobacteriumbifidum(pGEX-Sj32) ofSchistosomajaponicum[J]. Chin J Endemiol, 2015, 34 (1): 37-40. (in Chinese) 谭建蓉,李文桂,张丽.日本血吸虫重组两歧双歧杆菌(pGEX-Sj32)疫苗构建及鉴定[J].中华地方病学杂志,2015,34 (1): 37-40.
[6]Xiang JP, Li WG, Zhang L. Dynamic changes of proliferation, subsets and apoptosis of splenocytes from BALB/c mice immunized with recombinant Bb(pGEX-Sj26GST-Sj32) vaccine againstSchistosomajaponicum[J]. J Cellular Mol Immunol, 2013, 29(11): 1129-1132. (in Chinese) 向进平,李文桂,张丽.日本血吸虫重组Bb(pGEX-Sj26GST-Sj32)疫苗免疫小鼠脾细胞增殖、亚群及凋亡的动态变化[J].细胞与分子免疫学杂志,2013,29(11): 1129-1132.
[7]Ji MJ, Su C, Wang Y, et al. Characterization of CD4+T cell responses in mice infected withSchistosomajaponicum[J]. Acta Biochim Biophys Sin, 2006, 38(5): 327-334. DOI: 10.1111/j.1745-7270.2006.00169.x
[8]Mushtaq K, Chodisetti SB, Rai PK, et al. Decision-making critical amino acids: role in designing peptide vaccines for eliciting Th1 and Th2 immune response[J]. Amino Acids, 2014, 46(5): 1265-1274. DOI: 10.1007/s00726-014-1692-4
[9]Tu Y, Hu Y, Fan G, et al. Protective effects of membrane-anchored and aecreted DNA vaccines encoding fatty acid-binding protein and glutathione S-transferase against Schistosomajaponicum[J]. PLoS One, 2014, 9(1): e86575. DOI: 10.1371/journal.pone.0086575
[10]Fonseca CT, Brito CF, Alves JB, et al. IL-12 enhances protective immunity in mice engendered by immunization with recombinant 14 kDaSchistosomamansonifatty acid-binding protein through an IFN-gamma and TNF-alpha dependent pathway[J]. Vaccine, 2004, 22(3/4): 503-510. DOI: 10.1016/j.vaccine.2003.07.010
[11]Cheng YL, Song WJ, Liu WQ, et al. The effects of interleukin (IL)-12 and IL-4 deficiency on worm development and granuloma formation inSchistosomajaponicum-infected mice[J]. Parasitol Res, 2012, 110(1): 287-293. DOI: 10.1007/s00436-011-2487-0
[12]Cardoso FC, Macedo GC, Gava E, et al.Schistosomamansonitegument protein Sm29 is able to induce a Th1-type of immune response and protection against parasite infection[J]. PLoS Negl Trop Dis, 2008, 2(10): e308. DOI: 10.1371/journal.pntd.0000308
[13]Farias LP, Cardoso FC, Miyasato PA, et al.SchistosomamansoniStomatin like protein-2 is located in the tegument and induces partial protection against challenge infection[J]. PLoS Negl Trop Dis, 2010, 4(2): e597. DOI: 10.1371/journal.pntd.0000597[14]Wang X, Dong L, Ni H, et al. Combined TLR7/8 and TLR9 ligands potentiate the activity of aSchistosomajaponicumDNA vaccine[J]. PLoS Negl Trop Dis, 2013, 7(4): e2164. DOI: 10.1371/journal.pntd.0002164
[15]Cardoso FC, Pacifico RN, Mortara RA, et al. Human antibody responses of patients living in endemic areas for schistosomiasis to the tegumental protein Sm29 identified through genomic studies[J]. Clin Exp Immunol, 2006, 144(3): 382-391. DOI: 10.1111/j.1365-2249.2006.03081.x
[16]Tu Y, Fan G, Chen Z, et al. Protective effects of membrane-anchored and secreted DNA vaccines encoding fatty acid-binding protein and glutathione S-transferase againstSchistosomajaponicum[J]. PLoS One, 2014, 9(1): e86575. DOI: 10.1371/journal.pone.0086575
[17]Dai Y, Wang X, Zhao S, et al. Construction and evaluation of replication-defective recombinant optimized triosephosphate isomerase adenoviral vaccination inSchistosomajaponicumchallenged mice[J]. Vaccine, 2014, 32(7): 771-778. DOI: 10.1016/j.vaccine.2013.12.059
[18]Wang X, Dai Y, Zhao S, et al. Pamam-Lys, a novel vaccine delivery vector, enhances the protective effects of theSjC23 DNA vaccine againstSchistosomajaponicuminfection[J]. PLoS One, 2014, 9(1): e86578. DOI 10.1371/journal.pone.0086578
[19]Lee EF, Young ND, Lim NTY, et al. Apoptosis in schistosomes: toward novel targets for the treatment of schistosomiasis[J]. Trends Parasitol, 2014, 30(2): 75-84. DOI: 10.1016/j.pt.2013.12.005
[20]Chen L, Rao KV, He YX, et al. Skin-stage schistosomula ofSchistosomamansoniproduce an apoptosis-inducing factor that can cause apoptosis of T cells[J]. J Biol Chem, 2002, 277(37): 34329-34335. DOI: 10.1074/jbc.M201344200
Li Wen-gui, Email: cqliwengui@163.com
Dynamic study on immune responses of BALB/c mice induced by the recombinant Bb (pGEX-Sj32) vaccine againstSchistosomajaponicum
TAN Jian-rong1,LI Wen-gui2,QIN Ting1
(1.DigestivesystemdepartmentBofXiangyanghospitalaffiliatedtoHubeiuniversitymedicine,Xiangyang,441000; 2.InstituteofInfectiousandParasiticDisease,theFirstAffi1iatedHospital,ChongqingMedicalUniversity,Chongqing400016,China)
To observe the dynamic changes of immune responses in mice immunized with recombinant vaccineBifidobacteriumbifidum(pGEX-Sj32) ofSchistosomajaponicumand study the immune mechanisms of the recombinant vaccine, each mouse in this study was immunized with recombinant Bb (pGEX-Sj32) vaccine by per os (PO group) and intranasally (IN group) with recombinant vaccine. During week 0-20 after immunization, splenocytes and eye blood sera of 4 mice from each group were separated at 2-week interval. Enzyme-linked immunosorbent assay (ELISA) was performed to detect the levels of antibodies IgG, IgG1, IgG2a, IgG2b, IgG3, IgE, IgA and cytokine interleukin(IL)-10, IL-12 and tumor necrosis factor(TNF)-a. Splenocyte proliferation was investigated by MTT colorimetric assay, subsets of CD4+and CD8+T cells and apoptosis of splenocytes by FACsort flow cytometry, respectively. The maximum proliferation efficiencies of both PO and IN group got on week 6 after immunization, while the apoptotic rates appears on 6thweek without any stimulation as well as on 8thweek with ConA. Rates of CD4+T and CD8+T cells subsets reached to the peak on 6thand 8thweek in PO group after immunization while on 8thand 12thweek in IN group, respectively. The level of TNF-α, IL-10 and IL-12 in splenocyte supernatant peaked on the 8th, 10thand 6thweek in group PO, while got the maximum on the 8th, 10thand 8thweek in group IN, respectively. Except for IgE reached peak on the 14th week, all mice developed significant peaks of specific IgG, IgG1, IgG2a, IgG2b, IgG3 and IgA on the 8th week in PO group and those appeared on the 12thweek (IgG and IgA peaked on week 8) in group IN, respectively, compared to 0 week (P<0.05 orP<0.01). The level of IL-10, IL-12 and TNF-αwhich detected in serum peaked on the 14th, 6thand 8thweek respectively in group PO, and got the maximum on the 8th, 6thand 8thweek in group IN, respectively. The rBb (pGEX-Sj32) vaccine may induce mice to produce immune response effectively.
Schistosomajaponicum; rBb (pGEX-Sj32) vaccine; immune
10.3969/j.issn.1002-2694.2015.09.005
重庆市科委地方病重大专项基金资助(2008AB5055;2008AB5008;2008AB5054)
李文桂,Email:cqliwengui@163.com
1.湖北医药学院附属襄阳医院消化内科B病区,襄阳 441000; 2.重庆医科大学附属第一医院传染病寄生虫病研究所,重庆 400016
Supported by the Major Special Found of Endemic Disease of Chongqing (Nos. 2008AB5005, 2008AB5008 and 2008AB5054)
R383.2
A
1002-2694(2015)09-0805-07
2014-11-11;
2015-02-02
猜你喜欢
中国医药科学(2022年5期)2022-05-05
健康之家(2021年19期)2021-05-23
中华养生保健(2020年2期)2020-11-16
中国感染与化疗杂志(2020年1期)2020-01-12
中国动物检疫(2018年7期)2018-07-04
家庭用药(2018年3期)2018-03-23
医药前沿(2018年2期)2018-01-11
中成药(2017年9期)2017-12-19
腹腔镜外科杂志(2016年9期)2016-06-01
中国当代医药(2015年33期)2015-03-01
