
林地生物量遥感估算模型进展及在竹林碳汇移植前景
2015-12-30 04:10蒋小凡张肖清顾志康李荣华岳言龙
浙江林业科技 2015年2期
蒋小凡,李 东,邵 飞,张肖清,孟 晖,顾志康*,李荣华,岳言龙
(1. 浙江省湖州市林业科学研究所,浙江 湖州 313000;2. 山东省国有林场管理站,山东 济南 250014;3. 浙江省湖州市湖州木材检查站,浙江 湖州 313000;4. 浙江省湖州市太湖旅游度假区仁皇山街道,浙江 湖州 313100;5. 浙江省湖州市林业局,浙江 湖州 313000)
林地生物量遥感估算模型进展及在竹林碳汇移植前景
蒋小凡1,李 东1,邵 飞2,张肖清3,孟 晖4,顾志康1*,李荣华5,岳言龙5
(1. 浙江省湖州市林业科学研究所,浙江 湖州 313000;2. 山东省国有林场管理站,山东 济南 250014;3. 浙江省湖州市湖州木材检查站,浙江 湖州 313000;4. 浙江省湖州市太湖旅游度假区仁皇山街道,浙江 湖州 313100;5. 浙江省湖州市林业局,浙江 湖州 313000)
从林地生物量的估算方法、遥感估算模型的分类、遥感经验估算模型和遥感经验估算模型的可移植性方面概述了森林碳汇研究中的生物量估算研究进展,分析了在竹林碳汇研究中移植前景。
遥感估算;生物量;竹林;碳汇
近年来由于全球气候变化,使不少专家学者的研究集中在植被对全球碳循环和平衡作用方面。通过植被生物量来探讨植被生态系统的碳“汇”功能,更是重要的研究内容[1]。森林生物量约占全球陆地植被生物量90%,不仅是森林固碳能力的重要标志,也是评估森林碳收支的重要参数,因此,森林生态系统总生物量和生产力的研究,是判断森林生态系统中大气CO2的源和汇的重要标志[2]。
毛竹(Phyllostachys heterocycla cv. pubescens)林生态系统特有的结构与功能及人为经营方式使其有别于其他类型的森林生态系统,是我国竹林中分布最广、面积最大的一种竹林[3],在全球森林面积急剧下降的今天,竹林面积却以3%的速度在递增,这也就是意味着,竹林是一个不断增大的碳汇[4]。在自然界中竹类的营养生长和无性繁殖能力较强,竹林生态系统的实际贮碳能力不容忽视。目前,许多学者已对世界范围内不同地区、不同类型森林的生物量和碳储量进行了估算。
为此,笔者对林地生物量的估算方法、遥感估算模型的分类、遥感经验估算模型和遥感经验估算模型的可移植性4个方面的研究做如下概述。
1 林地生物量常规估算方法
传统的生物量研究一般采用以实测数据为基础进行宏观拓展估算或相关分析的方法:先选取样区,利用收获法实地称量林木的干重、湿重、树干重量、叶重、地下部分重量等数据,或以收获法为基础,利用每木调查、树干解析、材积转换等方法进行各部分生物量及总生物量的测量,而后利用这些数据进行宏观估算,以获知整个研究区域的生物量状况;或者对样区内生物量及其影响因素进行分析,建立相关模型并推而广之。传统生物量的研究方法有皆伐法、平均生物量法、生物量回归模型估计法、材积源生物量法。生物量的研究方法比较成熟,国内外都取得了一定的研究成果[5]。
2 遥感估算模型的分类
生物量的遥感估算多利用红波段和近红外波段的组合即植被指数(Vegetation Indices)和叶面积指数(LAI)及植被覆盖度等的关系,推断出植被指数与生物量之间的关系进而求得生物量。在森林生物量的研究中,常用的遥感数据源主要包括光学遥感、热红外遥感、微波遥感和高光谱遥感等[6]。
在可见光至红外光波谱段,植物的反射光谱曲线具有显著的特征,但是不同类型的植物之间反射光谱特性曲线存在着一定的差异,即使是同一种植物,也由于其生长状态的不同,使其在各个波段的反射率发生变化[7],因此可以利用植物的这一光谱特性,监测植被状况及其变化动态。卫星遥感数据中属于这一类的主要有Landsat TM数据、MSS数据、SPOT数据以及NOAA AVHRR数据等。最初,人们利用Landsat MSS数据来监测植被的叶面积指数和活体生物量[8]。后来,更多的是利用Landsat TM和NOAA AVHRR数据来监测植被生长和生物量[9]。Lefsky等[10]利用地面调查数据和TM数据对美国EasMaryland落叶林的地上部分生物量进行了估算。Boyd 等[11]对喀麦隆的热带森林生物量的遥感研究表明,中红外植被指数与热带森林植被的总生物量具有较好的相关性,其相关系数为0.47。在国内,郭志华等[12]通过样方调查获取森林材积,借助于全球定位系统技术为调查样方准确定位。根据Landsat TM数据7个波段信息及其线形与非线形组合,应用逐步回归技术分别建立估算针叶林和阔叶林材积的最优光谱模型,进而确定了粤西及附近地区的森林生物量。马泽清等[13]利用TM数据对江西千烟洲地区的湿地松林生物量估算,建立了生物量多元回归模型。Hame等[13]结合地面调查数据、TM 数据和AVHRR数据,对数百万平方公里欧洲森林生物量进行了准确的估算。
合成孔径雷达是在真实孔径侧视雷达的基础上发展起来的,克服了真实孔径雷达受到天线长度限制而无法较大幅度地提高分辨率的困难[14]。研究表明,不同波段的雷达数据对森林生物量的敏感度不同,雷达后向散射对森林生物量的敏感性或相关性随着波长的增加而增加,即P-波段与生物量的关系最为密切,而C-波段则较差[15]。Harrll[16]曾指出,合成孔径雷达(SAR)在区域或局部尺度的生物量监测、生物量分布等领域是一种非常有希望、有前途的工具,能获取生物量计算所需的主要参数(密度、胸径、高度和年龄等)。开展了星载SAR对森林蓄积量估算的研究,也取得了一些成果[17]。
传统的多光谱遥感只是在几个离散的波段,以不同的波段宽度(常为100 ~ 200 nm)来获取图像,这样就丢失了对地物识别有用的大量光谱吸收特征信息。而高光谱遥感波段宽度一般小于 10 nm,它可以将光谱波段在某一特定光谱域进行细分,从而获得多波段的详实、连续的光谱信息,这些光谱可以很好地描述植被特征。童庆禧等[18]建立了 LAI及生物量与高光谱遥感图像的植被因子之间的理论模型。张良培等[19]利用高光谱对APAR和生物量进行了估算。Thenkabail等[20]则研究了最适多窄波段反射系数OMNBR(Optimum Multiple Narrow Band Reflectivity)、窄波段NDVI、窄波段TSAVI以及NDVI(宽波段)和作物特征的关系,指出窄波段模型要比宽波段模型更好。
国外许多研究已经证明机载小光斑雷达数据在森林资源调查中的重要性,通过激光扫描数据可以准确地估测林分特征,如树高、胸高断面积以及林分蓄积量。Lefsky等根据植被材积及生物量与高度的关系,发现运用激光雷达信号可以很好地估算森林生物量。Popescu等[21]用小光斑雷达数据和多光谱数据进行小样区(0.017 hm2)水平的材积和生物量的估测,研究首先用局部最大值法得到单株木树高和冠幅,然后用回归模型计算胸径和胸高断面积,根据已有的生物量和材积计算公式分别计算其生物量和材积。在国外已经有了许多此类研究,但是,国内激光雷达在林业应用领域的研究则刚刚起步。
3 遥感经验估算模型
经验模型[2]一般是描述性的,不涉及机理问题,主要是对观测数据进行经验性的统计描述或者是通过相关分析,建立适当的公式用于估算。目前主要的经验模型是将植被指数如NDVI等与生物量进行回归分析,从而得到估算的经验式。这些回归式有线性、幂函数和对数形式,而且系数也各不相同。除了植被指数,还可以直接利用遥感器各波段的线性或非线性组合来建立生物量的估算公式。这些估算模型的优点是简单、便于计算,缺点是形式多种多样,易受植被类型以及非植被因素如土壤背景、大气条件、地形和地表二向性反射特性的影响。而且,这些模型中参数的生态学原理目前尚不清楚。由于遥感象元表达的是植被和其它因素的混合反应,未来的植被指数的研究应以“植被—土壤—大气”系统为一个整体目标,并考虑到遥感器和电磁波的辐射特性。
为了克服经验模型的缺陷,许多学者提出了基于植被二向反射特性的物理模型,如考虑辐射传输的 3D模型、几何光学的间隙率模型。由植被的结构特征和光谱特征计算植被BRDF是遥感的正向问题;由植被BRDF使用相应的计算方法生成植被结构称为反演问题。植被冠层的反演有两种方案:一是用多光谱信息反演即通过光谱变换得到各种植被系数,这类方法存在着许多问题和困难,精度也不够;二是通过多角度遥感信息进行反演。
半经验模型综合了经验模型与物理模型的优点,通常使用的参数很少,但这些参数多少具有一定的物理学意义,例如Ross表层核、Roujoan模型、Verstraote模型、Wannor核驱动模型等,这种折衷模型具有一定的应用前景。
综合模型借助遥感信息和植被信息、气象因子等来建立,由于包含了更多的信息量,可以更加精确地反映植被的生物物理参数。
3.1 线性回归模型
3.1.1 植被指数作为自变量构建模型 李仁东[22]等在应用Landsat ETM数据估算鄱阳湖湿生植被生物量研究中分析采样数据与遥感数据之间的相关关系,结果表明:采样数据与ETM 4波段数据相关性最高,其相关系数达到0.86。采样数据与差值植被指数(DVI,Difference Vegetation Index)、归一化植被指数(NDVI,Normalized Difference Vegetation Index)数据的相关系数分别为0.83、0.80。采样数据与第一主成分数据之间的相关关系不显著,仅0.40,并建立了采样数据与ETM4波段数据的线性相关模型。Lee[23]等发现日本西部Hiroshima地区森林生物量与NDVI及DVI之间存在线性关系,松树与NDVI的相关系数为0.85,日本雪松和落叶阔叶林与DVI的相关系数分别为-0.83和0.8。
3.1.2 原始波段及其派生波段作为自变量构建模型 郭志华[24]等对粤西地区阔叶林和针叶林材积与反射辐射的关系进行研究,发现波段TM1、TM2、TM3和TM6与阔叶林和针叶林材积之间呈极显著负相关(p < 0.01);阔叶林和针叶林材积与NDVI、比值植被指数(RVI,Ratio vegetation index)间呈极显著正相关(p < 0.01)。进一步的研究表明仅以TM3为自变量就可获得估算阔叶林和针叶林材积的光谱模型,模型的常数项和系数也均达到0.001的显著水平,模型相关性较高。
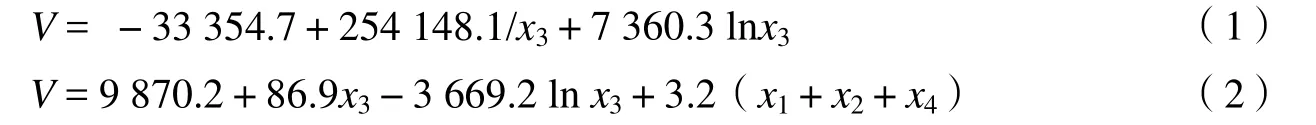
式中,V为材积,x1、x2、x3分别为波段TM1、TM2、TM3,x4为比值植被指数。3.1.3 地形或生物因子参与下的线性模型 国庆喜[25]等以海拔、坡度、土壤厚度、平均年龄、郁闭度、平均直径、平均树高、TM1、TM2、TM3、TM4、TM5、TM6、TM7、共20个变量因子为待选自变量,进行逐步回归筛选出以下13个自变量包括海拔、坡度、土壤厚度、平均年龄、郁闭度、TM2、TM3、TM4、TM6、TM7、TM4、来估测森林的生物量依次编
3号为X1~ X13回归模型为(3)式,回归方程决定系数R2= 0.712 5。
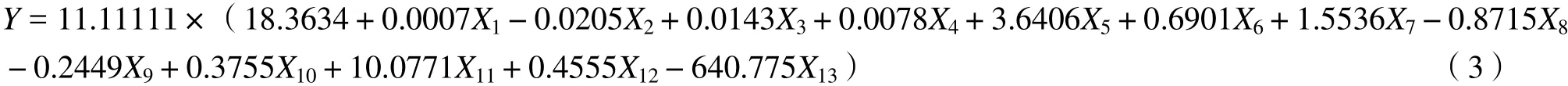
LU[26]等利用TM数据探讨影像纹理(texture)与森林地上生物量相关性。研究结果表明:纹理能够提高成熟林生物量估算精度,尤其是对于林分结构复杂的森林,而对演替林生物量估算所起的作用不明显。将纹理与光谱信号相结合估测生物量比单单用纹理或光谱信号估测精度高。
3.2 非线性回归模型
3.2.1 指数模型 金丽芳[27]等利用Landsat TM数据求得归一化植被指数(NDVI)并以它为自变量与生物量之间建立指数函数模型,决定系数R2达到0.91;Gilabert等[25]利用实验遥感方法对作物冠层的叶面积指数(LAI)、生物量与NDVI的关系进行研究,构建 X =A+ BlnY 模型:

式中,为Y为生物量,X为归一化植被指数,A、B是据研究区状况而调整的参数。
3.2.2 幂函数模型 洪保章[28]利用650、700、750、800 nm波段的RVI(R750/R650、R750/R700、R800/R650、R800/R700)作为自变量,分别与毛豆地上鲜生物量进行对数、指数、幂函数以及多项式回归分析,发现R750/R700构造的RVI与毛豆地上鲜生物量之间存在较好的幂函数关系。
3.2.3 多项式模型 李健[29]等在基于 Landsat_TM 数据建立鄱阳湖湿地植被生物量遥感监测模型中,利用生物量数据与植被指数DVI、RVI、NDVI、SAVI以及变换归一化植被指数(TNDVI)进行了三次多项式的非线性回归分析,结果显示DVI和生物量的三次函数非线性回归分析拟合度较高,其模型为:
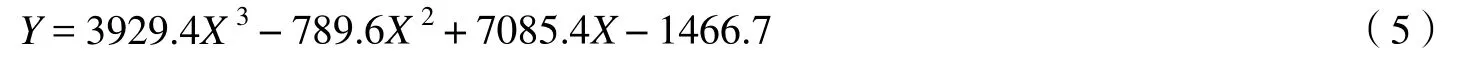
式中,Y为生物量,X为DVI,决定系数R2达到0.859 5。
3.2.4 双曲函数曲线 邢素丽[30]等探讨基于ETM数据的落叶松林生物量的估算方法和模式,开发18个自变量,经过逐步回归分析得出:ETM3数据是落叶松林生物量的最好估计因子,生物量与ETM3数据具有双曲函数关系,与1/ETM3呈极显著正相关,相关系数为0.878。
3.3 人工神经元网络模型
人工神经元网络(Artificial Neural Networks, ANN)是一种较好的非参数模型,其在遥感应用研究中属于一种较新的方法。范文义等[31]采用黑龙江长白山地区的TM影像和133块森林资源一类清查样地的数据,选取地学参数、遥感反演参数等71个自变量分别构建神经网络模型模型,进而估算该地区的森林生物量,结果表明,传统神经网络模型估测森林生物量精度较好。
国庆喜[25]等以13个因子作为输入变量,用BP(后向传播Back propagation)神经网络模型进行模拟,经模型反演得出的估计值与实测值比较得出决定系数R2= 0.906。宋开山等[32]对玉米地上鲜生物量的高光谱遥感估算模型进行研究,表明以光谱的550 nm、660 nm与680 nm以及710 nm处的反射率为输入变量建立BP网络能够准确预测玉米的地上鲜生物量,模型预测结果相对与由550 nm、700 nm两个光谱反射率构成的二元线性模型来说,其精度较高。
人工神经网络模型的精度相对较高,不足之处主要是由于人工神经网络方法的“黑箱”操作,使得它不利于揭示森林生物量形成的内在机理。
4 遥感经验估算模型的可移植性的研究
在经验模型的研究中,许多学者在研究中对模型移植的不确定因素取得了一定程度上的突破。国内 CHEN Liangfu等[33]用中巴地球资源卫星CBERS-02CCD数据提取的NDVI和样地测量数据估算了千烟洲附近马尾松和湿地松的生物量,发现由于植被指数的饱和、重采样时像元大小的改变引起空间和光谱的改变、表面异质性、分辨率的差异等原因的存在,使得该模型有一定的局限性。国外Giles M Foodya等[34]基于TM波段数据,利用植被指数法、多重回归和前馈神经网络三种模型,对热带森林生物量进行估算预测,并比较三种模型的预测效果,结果表明,人工神经网络反映的预测值和实测值关系最强(相关系数扩 > 0.71,置信水平为99%);模型移植后精度均会降低,其中人工神经网络模型的移植性相对较好。研究发现遥感数据在预处理时的处理方法相同与否、地面调查数据测量数据不准确、时空尺度的差异导致遥感和地面数据的不祸合等原因都是模型移植的不确定因素。
近年来,森林生态系统碳储量及其碳汇功能的研究已日益深入,但毛竹林生态系统研究由于起步较晚,在这方面还存在着较大的差距,尤其国内的竹林碳储量估算研究还处于区域性的研究阶段,普适性估算模型的较少,今后应加大毛竹林生态系统研究的科技发展力度,在研究方法上还要不断改进、完善和创新,在研究内容上将日益丰富和深入,综合运用多种方法,使普适性估算模型研究向更高层次发展,为更高层次的竹林碳汇研究奠定基础。
[1] 魏晶,吴钢,邓红兵. 长白山高山冻原植被生物量的分布规律[J]. 应用生态学报,2004,15(11):1999-2004.
[2] 刘炜洋,梁嘉鑫,李志新. 生物量的遥感估算方法及影响因素[J]. 林业科技情报,2010,42(2):1-2.
[3] 王燕,王兵,赵广东,等. 我国毛竹林生态系统碳平衡研究进展[J]. 林业科技开发,2008,22(4):9-12.
[4] 郭起荣,杨光耀,杜天真,等. 中国竹林的碳素特征[J]. 世界竹藤通讯,2005,3(3):25-28.
[5] 王维枫,雷渊才,王雪峰,等. 森林生物量模型综述[J]. 西北林学院学报,2008,23(2):58-63.
[6] 万猛,李志刚,李富海,等. 基于遥感信息的森林生物量估算研究进展[J]. 河南林业科技,2009,29(4):42-45.
[7] 吕国揩. 遥感概论[M]. 北京:高等教育出版社,1994.
[8] Peng S L, Guo Z H, Wang B S. Application of RS and GIS on Terrestrial Vegetation Ecology[J]. Chin J Ecol, 1999, 18(5):52-64.
[9] Hame T, Salli A, Andersson K, et al. A New Methodology for the Estimation of Biomass of Conifer-Dominated Boreal Forest Using NOAA-AVHRR Data[J]. Int J Remote Sens, 1997, 18(15):3 211-3 243.
[10] Lefsky M A, Harding D, Cohen W B, et al. Surface Lidar Remote Sensing of Basal Area and Biomass in Deciduous Forests of Eastern Maryland, USA[J]. Remote Sens Environ, 1999(67):83-98.
[11] Boyd D S, Foody G M, Curran P J. The relationship between the biomass of Cameroonian tropical forests and radiation reflected in middle infrared wavelengths[J]. Int J Remote Sens, 1999, 20(5):1 017-1 023.
[12] 郭志华,彭少麟,王伯苏. 利用TM数据提取粤西地区的森林生物量[J]. 生态学报,2002,22(11):1 832-1 839.
[13] 马泽清,刘琪璟,徐雯佳,等. 江西千烟洲人工林生态系统的碳蓄积特征[J]. 林业科学,2007,43(11):1-7.
[14] 张钧屏. 对地观测与对空监视[M]. 北京:科学出版社,2001.
[15] Toan L, Beaudoin T, Riom A, et al. Relating forestbiomass to SAR data[J]. Geosci Remote Sens, 1992, 30(2):403-411.
[16] Harrell P A, Bourgeau-Chavez L L, Kasischke E S, et al. Sensitivity of ERS-1 and JERS-1 radar data to biomass and stand structure in Alaskan Boreal Forest[J]. Remote Sens Environ, 1995(54):247-260.
[17] 李增元,车学俭.ERS-1SAR影像森林应用研究初探[J]. 林业科学研究,1994,7(6):692-696.
[18] 童庆禧,郑兰芬. 湿地植被成像光谱遥感研究[J]. 遥感学报,1997,1(1):50-57.
[19] 张良培,郑兰芬,童庆禧. 利用高光谱对生物量进行估计[J]. 遥感学报,1997,1(2):111-114.
[20] Thenkabail P S, R B Smith, E De Pauw. Hyperspectral vegetation indices and their relationships with agricultural crop characteristics[J]. Remote Sens Environ, 2000, 7(1):158-182.
[21] Popescu S C, Wynne R H, Scrivani J A. Fusion of small-footprint LiDAR and multispectral data to estimateplot-level volume and biomass in deciduous and pine forests in Virginia, USA[J]. For Sci, 2004, 50(4):551-565.
[22] 李仁东,刘纪远. 应用Landsat ETM数据估算鄱阳湖湿生植被生物量[J]. 地理学报,2001,56(5):532-540.
[23] Lee N J, Nakane K. Forest vegetation classification and biomass estimation based on Landsat TM data in a mountainous region of west Japan[A]. Gholz H L, K Nakane, H Himoda eds. The use of remote sensing in the modeling of forest productivity[C]. Kluwer Academic Publishers, Netherlands, 1997. 159-171.
[24] 郭志华,彭少麟,王伯荪. 利用TM数据提取粤西地区的森林生物量[J]. 生态学报,2002,22(11):832-839.
[25] 国庆喜,张锋. 基于遥感信息估测森林的生物量[J]. 东北林业大学学报,2003,31(2):13-16.
[26] Lu D, Mausel P, Brondízio E, et al. Relationships between Forest Stand Parameters and Landsat Thematic Mapper Spectral Responses in the
Brazilian Amazon Basin[J]. For Ecol Manage, 2004(198):149-167.
[27] 金丽芳,徐希孺,张猛,等. 内蒙古典型草原地带牧草产量估算的光谱模型[J]. 中国草原与牧草杂志,1986,3(2):51-54.
[28] 洪保章. 毛豆遥感光谱与地上鲜生物量的相关性分析[D]. 福州:福建农林大学,2006.
[29] 李健,舒晓波,陈水森. 基于Landsat-TM数据鄱阳湖湿地植被生物量遥感监测模型的建立[J]. 广州大学学报(自然科学版),2005,4 (6):494-498
[30] 邢素丽,张广录,刘慧涛,等. 基于Landsat ETM数据的落叶松林生物量估算模式[J]. 福建林学院学报,2004,24(2):153-156.
[31] 范文义,张海玉,于颖,等. 三种森林生物量估测模型的比较分析[J]. 植物生态学报,2011,35(4):402-410.
[32] 宋开山,张柏,于磊. 玉米地上鲜生物量的高光谱遥感估算模型研究[J] .农业系统科学与综合研究,2005,21(1):65-67.
[33] CHEN L F, GAO Y H, CHENG Y, et a1. Biomass estimation and uncertainty analysis based on CBERS-02 CCD camera data and field
measurement[J]. Sci Chin(Ser E)Eng Mater Sci, 2005, 48(1):116-128.
[34] FOODY G M, BOYD D S. CULTER M E J. Predictive relations of tropical forest biomass from Landsat TM data and their transferability
between regions[J]. Remote Sens Environ, 2003(85):463-474.
Advance of Remote Sensing Estimation Model for Forest Biomass and Transferability for Bamboo Forest Carbon Storage
JIANG Xiao-fan1,LI Dong1,SHAO Fei2,ZHANG Xiao-qing3,MENG Hui4GU Zhi-kang1,LI Rong-hua5,YUE Yan-long5
(1. Huzhou Forestry Institute of Zhejiang, Huzhou 313000, China; 2. Administration of State Forest Farm of Shandong, Ji’nan 250014, China; 3. Huzhou Forestry Bureau of Zhejiang,Huzhou 313000, China; 4. Renhuangshan Street of Taihu Tourism Resort of Zhejiang, Huzhou 313000, China; 5. Huzhou Wood Inspection Station of Zhejiang, Huzhou 313000, China)
Introductions were made on estimation method of forest biomass, classification of remote sensing estimation models, remote sensing experience estimate model method and transferability of remote sensing experience estimate models. Advances were put forward to biomass estimation of forest carbon storage as well as to transferability for bamboo forest.
remote sensing estimation; biomass; bamboo forest; carbon storage;
S718.5
A
1001-3776(2015)02-0089-06
2014-09-11;
2015-01-10
蒋小凡(1973-),男,浙江长兴人,工程师,从事竹林培育与利用;*通讯作者。
猜你喜欢
广西林业科学(2022年3期)2022-07-08
航天返回与遥感(2022年2期)2022-05-12
草业科学(2022年3期)2022-03-26
农业机械学报(2021年8期)2021-08-27
农业机械学报(2019年6期)2019-06-27
江苏农业科学(2018年14期)2018-08-08
电子制作(2018年2期)2018-04-18
制导与引信(2017年3期)2017-11-02
电子制作(2017年8期)2017-06-05
绿色科技(2013年1期)2013-11-17
