
巴西橡胶树PPCK基因家族成员的鉴定和表达分析
2016-07-09 18:39肖小虎隋金蕾戚继艳阳江华朱晋恒唐朝荣
热带作物学报 2016年11期
关键词:表达分析
肖小虎 隋金蕾 戚继艳 阳江华 朱晋恒 唐朝荣
摘 要 以已发表的PPCK序列为探针,对橡胶树和其他5种植物(木薯、水稻、杨树、蓖麻和拟南芥)的转录组和基因组进行全面搜索,鉴定得到17个PPCK基因,其中包括3个橡胶树PPCK基因,命名为HbPPCK1-3。序列分析发现,HbPPCK1-3和其他植物一样在结构上非常保守,只有一个内含子和一个蛋白激酶结构域,并且相对位置也比较一致。在进化上,橡胶树、木薯和蓖麻3种大戟科植物的PPCK蛋白具有较近的亲缘关系。HbPPCK3在树皮中表达丰度最高,其次是种子和雄花;HbPPCK2在胶乳和根中表达丰度相对较高,而HbPPCK1在各组织中的表达都比较低。另外,大部分家族成员的表达都受到真菌侵染、低温和干旱胁迫处理的诱导,而乙烯利处理却能抑制HbPPCK2的表达。
关键词 巴西橡胶树;PPCK;基因家族;结构和进化;表达分析
中图分类号 S794.1 文献标识码 A
在植物中,磷酸烯醇式丙酮酸羧化酶(PEPC)以Mg2+或Mn2+为辅助因子, 催化磷酸烯醇式丙酮酸(PEP)和HCO3-生成草酰乙酸(OAA)和无机磷酸的不可逆性反应。PEPC广泛存在于光合生物, 如植物、藻类、蓝细菌和光合细菌中, 还存在于很多非光合细菌和原生动物中。PEPC催化的反应为细胞各种组分的生物合成提供四碳二羧酸, 参与维持柠檬酸循环,在初级代谢中有重要的补给作用,在C4植物和CAM植物的光合作用中催化大气中CO2固定的第一步反应,是C4光合作用途径中最重要的酶之一[1]。PEPC的活性主要是通过磷酸烯醇式丙酮酸羧化酶激酶(phosphoenolpyruvate carboxylase kinases,PPCKs)来调节[2]。PPCKs是钙调控型蛋白激酶的一员,但却缺少钙依赖型蛋白激酶所特有的自抑区(auto-inhibitory)和EF-hands结构域,所以PPCK是一种非钙依赖型蛋白激酶[2-3]。在景天酸代谢植物玉吊钟(Kalanchoe fedtschenkoi)、C4植物玉米已有研究和C3植物拟南芥中已有相关研究[3-5],研究认为PPCK的表达主要通过转录水平进行调控。在橡胶树中,橡胶的生产通过割胶来完成,割胶对橡胶树来说是一种机械伤害,而作为一种伤害信号却推动了橡胶的生物合成。人工使用乙烯利进行刺激可以显著提高胶乳的产量,正是应用了橡胶树树皮对激素刺激的应答能力,进而使橡胶树光合作用产物流向橡胶树的生物合成[6-7]。本研究以已发表PPCK蛋白氨基酸序列为探针[3-5],对橡胶树、木薯、蓖麻、拟南芥、杨树和水稻6种植物的全基因组数据进行全面搜索,鉴定得到17个PPCK基因家族成员,其中橡胶树中有3个。笔者利用相关软件和数据库,对橡胶树PPCK基因家族成员的基因结构、进化和表达水平进行了初步分析。研究结果将有助于笔者深入了解橡胶树PPCK基因在橡胶树生长发育和逆境胁迫应答过程中的功能。
1 材料与方法
1.1 材料
本实验室Solexa测序所用的巴西橡胶树(Hevea brasiliensis)热研7-33-97材料,除了根来自于组培苗外,其他组织(包括胶乳、树皮、树叶、种子、雌花和雄花)均来自中国热带农业科学院试验场三队的正常割胶橡胶树(开割2 a以上),根来自于中国热带农业科学院橡胶研究所种质资源圃华玉伟老师赠送的热研7-33-9组培苗;不同发育时期的橡胶树叶片来自于中国热带农业科学院橡胶所种质资源圃,为1年生的热研7-33-97嫁接苗;乙烯利处理的材料主要来自于海南省儋州市中国热带农业科学院试验场三队,品系为热研7-33-9,正常开割树(3天1刀,不涂乙烯利刺激),用1.5%乙烯利在不同时间处理橡胶树,相应4个时间点分别为0、3、12、24 h,具体实验参照相关文献中的方法进行[8]。
网上测序数据来自NCBI SRA数据库,包括不同组织(PRJNA201084),真菌侵染(Corynespora cassiicola tolerance, PRJNA179126,叶片)、高低温干旱胁迫(PRJNA182078,叶片)和乙烯利处理(PRJNA182079,胶乳),详细信息参见数据库中相关说明(http://www.ncbi.nlm.nih.gov/sra)。
1.2 方法
1.2.1 基因家族成员的鉴定与序列分析 为了全面鉴定橡胶树、拟南芥、杨树、水稻、木薯和蓖麻的PPCK基因家族成员,笔者从NCBI(http://www.ncbi.nlm.nih.gov/)和phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)数据库下载了6種植物的基因组和转录组数据。以已发表的PPCK序列为探针[3-5],对6种植物的基因组和转录组进行搜索,得到候选PPCK基因家族成员,利用InterProScan(http://www.ebi.ac.uk/Tools/pfa/iprscan/)对候选基因的结构域进行分析。再利用EMBOSS数据库的在线软件Pepstats(http://www.ebi.ac.uk/Tools/seqstats/emboss_pepstats/)对基因的等电点和分子量进行分析。
1.2.2 基因结构与进化分析 利用在线软件GSDS2.0(http://gsds.cbi.pku.edu.cn/)对橡胶树和其他5种植物PPCK基因家族成员的外显子/内含子组织结构进行分析。利用MEGA 6.0软件对橡胶树和其他5种植物的PPCK氨基酸序列一起构建系统发育树,采用Neighbor-Joining方法进行分子系统学分析,进行1 000次bootstrap统计学检验。
1.2.3 基因的表达模式分析 利用本实验室和NCBI的solexa转录组数据对橡胶树PPCK基因家族成员的表达模式进行分析[9]。首先,将NCBI的SRA数据库(http://www.ncbi.nlm.nih.gov/sra/)中橡胶树相关的solexa转录组数据下载到本地服务器,去除低质量序列,利用程序RSEM进行表达分析[10]。
2 结果与分析
2.1 PPCK基因家族成员的鉴定与序列分析
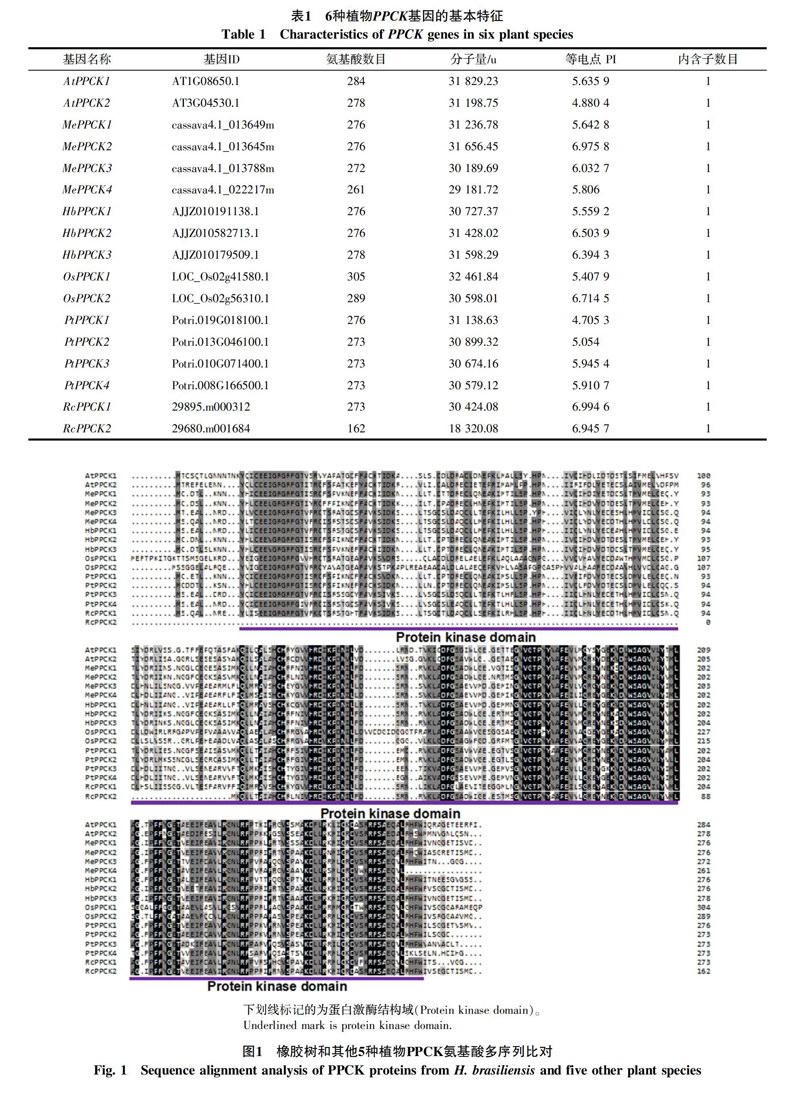
利用tblastn搜索橡胶树、木薯、蓖麻、拟南芥、杨树和水稻6种植物的转录组和基因组数据,鉴定得到17个基因家族(表1),其中包括3个橡胶树基因家族,命名为HbPPCK1-3。通过序列分析发现,橡胶树PPCK蛋白为276~278个氨基酸,分子量为31~32 ku,等电点在5.6~6.5之间,内含子数目均为1。氨基酸序列多序列比对发现,橡胶树PPCK蛋白都含有蛋白激酶保守结构域(图1)。
2.2 基因结构和进化分析
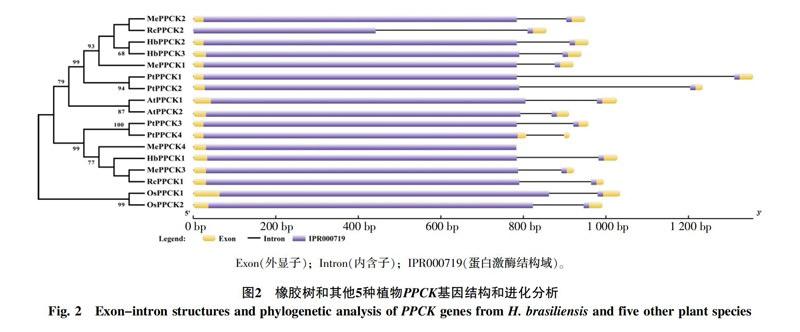
基因结构和进化分析发现(图2),6种植物基因家族成员在内含子数目和相对位置,以及结構域的相对位置上都非常一致,均仅有1个内含子和蛋白激酶结构域。在进化上,单子叶植物和双子叶植物分别聚在一起,而双子叶植物又分出2个分支,其中HbPPCK1位于一个分支,HbPPCK2和HbPPCK3位于一个分支。有部分物种存在同源蛋白对,如HbPPCK2和HbPPCK3、AtPPCK1和AtPPCK2、PtPPCK3和 PtPPCK4等,说明这些同源对具有较近的亲缘关系。橡胶树基因家族成员主要和木薯、蓖麻2种大戟科植物PPCK聚在一起,这3种植物具有较近的亲缘关系一致。
2.3 橡胶树PPCK基因家族成员的表达分析
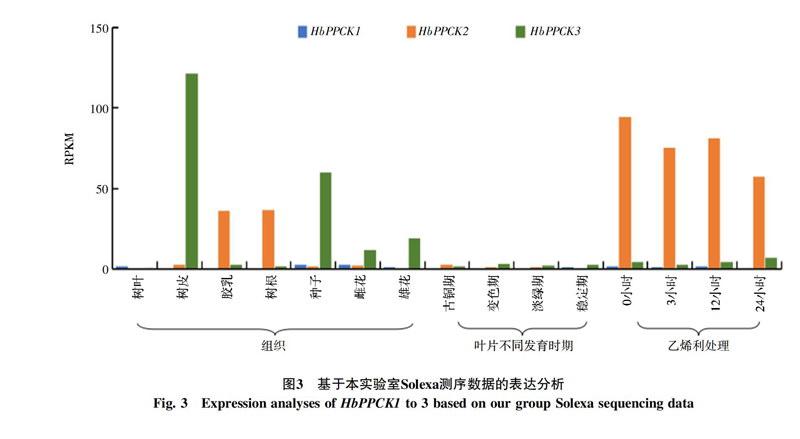
利用本实验室现有的Solexa测序数据对3个橡胶树PPCK基因在不同组织、不同叶片发育时期和乙烯利处理下的表达模式进行分析。结果表明:HbPPCK3在树皮中表达丰度最高,其次是种子和雄花;HbPPCK2主要在胶乳和根中表达,在其他组织中的表达偏低,随乙烯利刺激时间加长,表达呈下降趋势。HbPPCK1在所检测的所有组织中表达都很低(图3)。另外,各基因家族成员在叶片中的表达丰度相对其他组织都非常低,在叶片发育过程中也没有明显变化趋势。
笔者还利用NCBI的SRA数据库中的转录组数据对橡胶树PPCK的表达情况进行了分析。分析结果表明,HbPPCK3同样在树皮中表达丰度最高,C. cassiicola侵染、干旱、低温和乙烯利刺激都会不同程度的诱导HbPPCK3表达;HbPPCK2主要在胶乳中表达,低温处理后其表达丰度都会升高,而乙烯利刺激后表达丰度降低与本实验室数据结果一致;HbPPCK1在各组织中表达都很低,C. cassiicola侵染和低温处理会诱导其表达。
从以上两部分实验数据的结果来看,一个明显的特征就是HbPPCK3在树皮中高丰度表达,并且表达丰度要远远高于其他基因或其他组织,并且各种胁迫处理都会不同程度的诱导该基因的表达。这说明HbPPCK3可能在橡胶树响应逆境胁迫应答方面具有重要功能。另外,NCBI和本实验室数据结果的一致性,说明了本实验分析结果的准确性。
3 讨论
PEPC是C4光合作用途径中最重要的酶之一,PPCK通过磷酸化调节PEPC活性。本研究充分利用网上数据库资源,从橡胶树、木薯、水稻、杨树、蓖麻和拟南芥6种植物的全基因组数据中鉴定出17个PPCK基因家族成员,并从基因结构、进化和表达水平等方面进行了系统分析。在基因结构方面,和CDPK-SnRK超家族的其他成员相比,PPCK较为简单,仅有一个蛋白酶结构域(图2),除去激酶结构域以外的编码区很短。相对CDPK等其他CDPK-SnRK成员,PPCK缺少钙依赖型蛋白激酶所特有的自抑区(auto-inhibitory)和EF-hands 结构域,所以PPCK是一种非钙依赖型蛋白激酶[2-3]。在进化方面,PPCK主要分为2个较大的分支,单子叶植物和双子叶植物分别聚在一起,双子叶植物又分出2个分支,其中HbPPCK2和 HbPPCK3位于一个分支,HbPPCK1位于一个分支,结合表达分析的结果可以看出,HbPPCK1在各组织中表达丰度都比较低,而HbPPCK2和HbPPCK3分别在不同组中高丰度表达,另外拟南芥的2个PPCK成员也和HbPPCK2、HbPPCK3位于同一分支,因而可以初步推测HbPPCK2和HbPPCK3所处的分支为PPCK进化的主要方向。已有研究认为,PPCK基因的表达主要通过转录水平进行调控[3-5],在本研究中HbPPCK1在各组织中表达丰度都比较低或不表达,HbPPCK2和HbPPCK3的表达具有一定的组织特异性,并且受不同胁迫处理的调控,如HbPPCK3主要在树皮和种子中表达,真菌侵染、干旱、低温和乙烯利刺激都会不同程度的诱导其表达,HbPPCK2主要在胶乳中表达,低温处理可以诱导其在叶片中的表达,而乙烯利处理会降低其在胶乳中的表达,由此推测逆境条件能诱导PPCK基因的表达,进而对磷酸烯醇式丙酮酸羧化酶的活性进行调节。本实验室的高通量数据已成功应用于橡胶树基因组数据分析以及橡胶树蔗糖合成酶基因家族相关分析,相关结果已发表在Nature Plants[11]和 Febs Journal[9],这进一步证实了本文数据结果的可靠性。本研究利用相关软件和数据库,对橡胶树和其他植物PPCK基因家族成员的结构、进化和表达进行了初步分析,研究结果将为进一步研究PPCK基因在橡胶树光合作用和逆境胁迫应答等方面的功能奠定基础。
参考文献
[1] Hatch M D. C 4 photosynthesis: a unique elend of modified biochemistry, anatomy and ultrastructure[J]. Biochimica et Biophysica Acta(BBA)-Reviews on Bioenergetics, 1987, 895(2): 81-106.
[2] Hrabak E M, Chan C W M, Michael G, et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases.[J]. Plant Physiology, 2003, 132(2): 666-680.
[3] Hartwell J, Gill A, Nimmo G A, et al. Phosphoenolpyruvate carboxylase kinase is a novel protein kinase regulated at the level of expression[J]. Plant Journal for Cell & Molecular Biology, 1999, 20(3): 333-42.
[4] Shenton M. Distinct patterns of control and expression amongst members of the PEP carboxylase kinase gene family in C 4 plants[J]. Plant Journal, 2006, 48(1): 45-53.
[5] Fontaine V, Hartwell J, Jenkins G I, et al. Arabidopsis thaliana contains two phosphoenolpyruvate carboxylase kinase genes with different expression patterns[J]. Plant Cell & Environment, 2002, 25(1): 115-122.
[6] d'Auzac J, Jacob J L, Prévot J C, et al. The regulation of cis-polyisoprene production(natural rubber)from Hevea brasiliensis, In Recent research developments in plant physiology(ed S.G. Pandalai)[R]. Research Singpost, Trivandrum, India. 1997: 273-332.
[7] Hao B Z, Wu J L. Laticifer Differentiation in Hevea brasiliensis: Induction by Exogenous Jasmonic Acid and Linolenic Acid[J]. Annals of Botany, 2000, 85(1): 37-43.
[8] 肖小虎. 巴西橡膠树蔗糖代谢相关基因家族的克隆、 结构进化和表达分析[D]. 海南大学, 2013.
[9] Xiao X H, Tang C R, Fang Y, et al. Structure and expression profile of the sucrose synthase gene family in the rubber tree: indicative of roles in stress response and sucrose utilization in the laticifers[J]. Febs Journal, 2014, 281(1): 291-305.
[10] Li B, Dewey C N. Li B, etal. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome [J]. BMC Bioinformatics, 2011, 12(1): 93-99.
[11] Tang C, Meng Y, Fang Y, et al. The rubber tree genome reveals new insights into rubber production and species adaptation[J]. Nature Plants, 2016, 2: 16073.
猜你喜欢
热带农业科学(2016年12期)2017-03-31
中国中药杂志(2017年4期)2017-03-28
江苏农业科学(2016年11期)2017-03-21
中国中药杂志(2017年1期)2017-03-06
中国中药杂志(2016年20期)2016-11-19
江苏农业科学(2015年11期)2016-01-27
热带农业科学(2015年12期)2016-01-15
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年5期)2015-10-20
江苏农业科学(2015年9期)2015-10-20
