
蛹虫草交配系统与营养体亲和性研究
2016-12-24 06:13焦子伟努尔买买提伊力亚斯杨晓绒
新疆农业科学 2016年9期
焦子伟,努尔买买提·伊力亚斯,梁 月,杨晓绒
(1.伊犁师范学院化学与生物科学学院,新疆伊宁 835000;2.沈阳农业大学植物保护学院,沈阳 110866)
蛹虫草交配系统与营养体亲和性研究
焦子伟1,努尔买买提·伊力亚斯1,梁 月2,杨晓绒1
(1.伊犁师范学院化学与生物科学学院,新疆伊宁 835000;2.沈阳农业大学植物保护学院,沈阳 110866)
【目的】蛹虫草是一种药(食)用真菌,属典型的子囊菌异宗交配系统,研究蛹虫草交配系统和营养体亲和性。【方法】从蛹虫草子实体进行单孢子分离,获得不同的单子囊孢子群体。选取单子囊孢子菌株进行培养并提取菌丝体DNA,设计特异性引物对其交配型目的基因进行PCR扩增,明确不同群体内部各单子囊孢子菌株的交配型类型并分析其分离比例。随机选取不同交配型代表菌种采用对峙培养方法对相同和不同的交配型菌株的营养体亲和性进行检测。【结果】所有检测的菌株均具有两种交配型 MAT1-1或 MAT1-2之一。检测的不同群体菌株中两种交配型的分离比例,卡方检验均符合1∶1的理论比值。随机选取不同交配型的菌株进行营养体亲和性对峙培养试验,所选菌株被初步分为4个不同的营养亲和群,蛹虫草存在潜在的遗传分化。相同交配型菌株之间的营养亲和性较高(90%);相反不同交配型菌种之间的营养亲和性比例较低(60%)。【结论】所有供试单子囊菌株只具有一种交配型且未出现偏分离现象,其交配系统与营养体亲和性之间存在一定的相关性,相同交配型菌株组合比不同交配型菌株组合的营养亲和性更普遍,因此利用不同交配型且菌丝体营养亲和性较高的菌种组合,则更有利于发育产生原基,形成子实体。
蛹虫草;单子囊孢子;交配型;亲和性;营养亲和群
0 引 言
【研究意义】蛹虫草(Cordyceps militaris)最早源于中国,是一种珍贵的药(食)用真菌,寄生于鳞翅目昆虫蛹体后能形成真菌与昆虫复合体,故又名北冬虫夏草、北虫草、蛹草、蛹草菌等[1]。现代医学研究表明蛹虫草具有多种药用成分,具有滋肺益气、增精益肾、止血化痰、抑制肿瘤、延缓衰老、提高机体免疫力等作用,在医疗保健等领域具有广阔的市场[2,3]。由于蛹虫草生活环境限制,自然虫草资源逐年减少,导致蛹虫草子实体价格较高。人工培养蛹虫草子实体的技术日趋成熟,由于缺乏科学的理论基础和指导,在生产中经常出现菌种退化和子实体产量不高品质低下等瓶颈问题[4]。因此加强研究蛹虫草基础生物学,特别是遗传学领域的真菌交配系统与亲和性的相互关联,以期获得优质的稳定、高产蛹虫草菌种及提升人工栽培技术水平十分必要。【前人研究进展】近年来,国内外研究人员在人工培养、菌种选育、活性成分以及药理作用等方面对蛹虫草作了较为深入的研究[5]。特别是蛹虫草菌株育种方法有人工驯化、单孢子分离、组织分离、诱变、杂交等,并获得相对稳定的遗传菌株[6],但对蛹虫草的生物学和遗传学等研究方面仍较薄弱[7]。蛹虫草是典型的异宗配合的子囊菌,已证明菌丝融合能够比较客观地反映真菌种内菌株在遗传、形态、生理和生态等各方面的差异,营养亲和的个体亲缘关系近,不能亲和的个体亲缘关系远[8-10]。蛹虫草含有相互不同源的交配型基因MAT-1-1-1和 MAT1-1-2[11-12]。蛹虫草交配型基因 MAT1-1序列中α-Box(Genbank ID:AB124614)和MAT1-2序列中 HMG-Box(Genbank ID:AB124626)具有高度保守性,常被作为交配型的分子标记 用 于鉴 定 蛹虫 草 交 配型 的 不同[11,13]。通过交配型基因的特异引物,确定交配型基因的类型,进而鉴定蛹虫草的核相类型[13-14]。【本研究切入点】主要通过生物学及分子生物学方法,进行单子囊孢子分离,利用特异性引物扩增以明确菌种的交配型种类,通过统计学方法检测两种交配型分离比例,以获得不同群体内及整合群体的偏态分布情况,并进一步采用对峙培养方法,对相同和不同交配型菌株的营养亲和性进行检验和分析。【拟解决的关键问题】明确蛹虫草的交配系统与菌丝营养亲和性之间联系,为研究蛹虫草遗传分化性的分子机制提供理论基础,为蛹虫草栽培菌种的筛选与驯化提供新的技术参考。
1 材料与方法
1.1 材 料
1.1.1 供试菌 株
供试的新鲜人工栽培蛹虫草子实体取自学院蛹虫草人工栽培室,其亲本菌株源自新疆天山区域。
1.1.2 供试药品和培养基
改良马铃薯(PDA)固体培养基[15]:马铃薯200 g,葡萄糖20 g,蛋白胨3 g,琼脂16 g,磷酸二氢钾(KH2PO4)2 g,七水硫酸镁(MgSO4·7H2O)1 g,复合VB 25 mg,用水补至1 000mL,pH 6.5~7.0。
改良PDA液体培养基:马铃薯 200 g,葡萄糖 20 g,蛋白胨3 g,磷酸二氢钾(KH2PO4)2 g,七水硫酸镁(MgSO4·7H2O)1 g,复合VB 25 mg,用水补至1 000mL,pH 6.5~7.0。
1.0 %水琼脂:琼脂粉10.0 g,用水补足至1 000mL。
改良的DNA提取法[13]:抽提缓冲液(50 mM Tri-HCl,150 mM NaCl,100 mM EDTA),20% SDS,5 M NaCl,CTAB/NaCl溶液(10%CTAB,0.7 M NaCl),Tris饱和酚/氯仿/异戊醇(v/v/v,25∶24∶1),氯仿/异戊醇(24∶1),异丙醇,75%乙醇,TE(10 mM Tris-HCl pH 8.0,1 mM EDTA)和RNase A(100g/mL)。
1.2 方 法
1.2.1 单孢子分离
将蛹虫草子实体悬于 1.0%水琼脂培养基上方,使子实体上密布的子囊壳内的子囊所含的子囊孢子自然弹射,约 1~2 h。在体式解剖镜下用无菌挑针挑取只有单个子囊孢子的水琼脂小块,置于改良的固体 PDA培养基上,23℃下光照培养箱中培养,以获得分离的单子囊孢子蛹虫草菌株。随机选取4个不同来源的蛹虫草子实体分别进行单孢子分离,每个子实体至少分离 5个单子囊孢子菌株,源于同一子实体的菌种作为同一群体的代表菌株,每个单子囊孢子菌株命名为群体字母加数字组成。图1

图1 单子囊孢子分离Fig.1 Isolation of single-ascospores of Cordyceps militaris
1.2.2PCR扩增
DNA提取参照[13]的方法,根据交配型 MAT1-1利用引物(MAT1F,5′-CCATCTCATCGC GGATG-3′;MAT1R,5′-GGGCAAAACGACCATTG-3′),PCR反应体系20 μL,反应条件为:94℃,3 min;94℃变性 30 s,53℃退火 30 s,72℃延伸60 s,35个循环;72℃下延伸 10 min。根据交配型 MAT1-2采用引物(MAT2F,5′-ACATACGCTTGTCAAGA -3′,MAT2R 5′-AGGAGAGCCTTCT TGAT-3′),PCR反应体系20 μL,反应条件为:94℃,3 min;94℃变性30 s,48℃退火30 s,72℃延伸60 s,35个循环;72℃下延伸10 min。PCR产物通过 1.6%琼脂糖凝胶电泳检测,根据电泳结果确定其交配型的类型。
1.2.3 交配型分离
将不同群体的单子囊孢子菌株进行DNA提取和PCR检测,对不同来源的群体内的交配型分离比率进行分析,以获得交配型分离特点,并通过卡方检验群体是否符合理论上的交配型分离比率。
1.2.4 营养亲和性试验
选取不同交配型代表菌株,接种至改良 PDA固体培养基上,进行两两组合配对的对峙培养,每皿放置两个菌株,以观察不同和相同交配型菌株两两组合的亲和性,以同一菌株的自身配对为对照组。接种后培养物,放在23℃黑暗培养4 d,第5 d开始光照/黑暗(16 h/8 h)交替培养。观察记录菌落颜色、生长状态、有无气生菌丝;第14 d特别关注菌落结合区域菌丝亲和性,原基形成和子实体发育等。持续 28 d观察菌株间组合配对结果有无变化,参考李沛利等[16]的划分方法,对菌株间配对的结果进行归类。
2 结果与分析
2.1 交配型PCR鉴定
所有供试菌株均能够扩增出特异性片段,其中代表菌株 A2和 B3能够通过PCR扩增出交配型基因MAT1-1的α-Box片段,扩增特异性产物大小为233 bp。而菌株A9和B8能够通过PCR扩增出交配型基因MAT1-2的 HMG-Box片段,扩增特异性产物长度为191 bp。同时,不同交配性菌株 A2、B3和 A9、B8只能扩增出交配型MAT1-1或MAT1-2之一,说明代表菌株只具有一个交配型基因,没有 MAT1-1和 MAT1-2两种交配型共同存在的情况,是典型的二级异宗交配系统。图2

图2 单子囊孢子菌株交配型PCR鉴定Fig.2 Identification of mating type of single-ascospore strains by PCR
2.2 交配型分离
根据PCR结果,统计不同群体内单子囊孢子菌株的交配型分布。供试的四个不同群体交配型分布为:群体A,交配型MAT ratio(MAT1-1∶MAT1-2)=7∶3;群体 B,交配型MAT ratio=5∶3;群体C,交配型 MAT ratio=7∶7;群体 D,交配型MAT ratio=3∶2。37株蛹虫草单孢分离菌株有22株含有交配型基因MAT1-1,占所有供试菌株的59.5%;有15株含有交配型基因 MAT1-2,占所有供试菌株的40.5%。所有供试单孢分离菌株中交配型MAT ratio=22∶15。图3

图3 不同群体单子囊孢子菌株的交配型分布Fig.3 Distribution of Mating type of different single-ascospore strains
通过统计分析,进行卡方检测分析发现,4个独立的单子囊孢子群体内部交配型比例 χ2值均小于3.841(P<0.05),水平差异不显著,说明不同群体子囊孢子的交配型 MAT1-1和 MAT1-2分离比例符合1∶1;同时将4个群体进行整合,以作为地域代表群体进行分析表明,交配型MAT1-1和 MAT1-2分离比例符合 1∶1,即说明新疆地区人工栽培条件下所采集的单子囊孢子群体不存在偏态分布。表1
2.3 营养亲和性
随机选取两种不同交配型的菌株(MAT1-1:A2,A5,B3,B9,B11和MAT1-2:A1,A9,B1,B4,B8)在改良 PDA固体培养基上两两对峙培养,观察营养体亲和性情况,结果可以初步分为两大类(亲和与不亲和)及四种表型,包括 I型,亲和,菌落间无空白且交互处呈线状隆起并长出原基或子实体;II型,亲和,菌落间无空白地带,无隆起也未长出原基;III型,不亲和,一侧菌落边缘呈线状隆起并有原基,一侧边缘处有空白,两菌落未长平;IV型,不亲和,两菌落接触,一侧菌落中间边缘呈线状隆起及原基,一侧没有 。图4
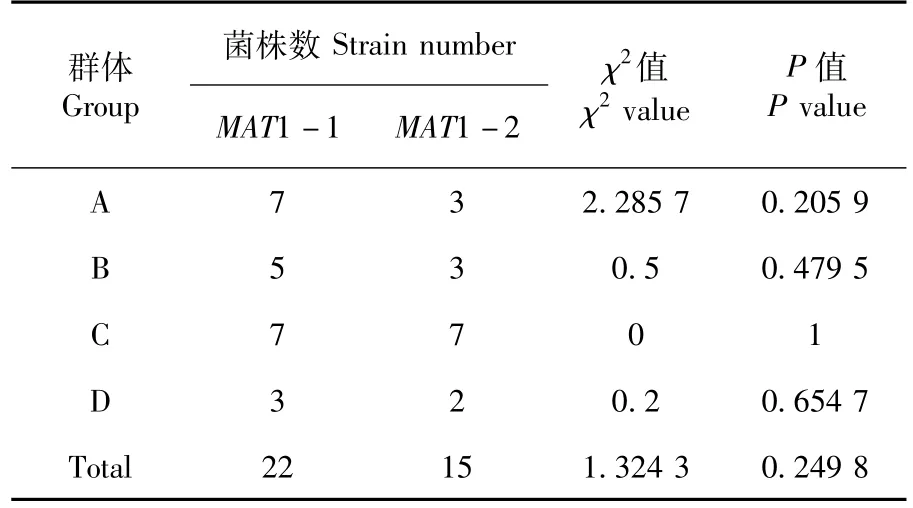
表1 不同群体交配型分离Table 1 Segregation of mating types in different groups
四种表现型在不同对峙培养组合之间的分布各不相同。具有相同交配型MAT1-1的菌株组合两两对峙培养时,配对菌落生长状态出现了三种表型,I型和 II型分别各有7对,III型为 1对,并未出现 IV型。具有相同交配型 MAT1-2的菌株组合两两对峙培养时,仅出现了13对 II型和2对 IV型,其中II型占到了将近 90%。另外,不同交配型MAT1-1与 MAT1-2的菌株对峙培养时,四种表现型均可观察到;其中,9对组合表现出I型,6对组合表现出II型,3对组合表现出 III型,7对组合表现出 IV型,出现频率从高到低为:I型≥IV型≥II型≥III型。表2

图4 菌株在改良 PDA培养基上对峙培养四种表型Fig.4 Four phenotypes of paired strains by crossing culture on modified PDA

表2 不同代表菌株在改良 PDA培养基上对峙培养表型分布Table 2 Distribution of phenotypes of paired strains by crossing culture on modified PDA
对所有供试单子囊孢子菌株营养亲性分析发现群体中,亲和性比例为 76.4%,不亲和比例23.6%。研究表明,不同交配型(MAT1-1与MAT1-2)对峙培养时,营养体亲和比例为 60.0 %,不亲和比例为40.0%。相同交配型(MAT1-1及 MAT1-2)对峙培养时,营养体亲和比例为90%,不亲和比例为10%;其中,交配型MAT1-1对峙培养时,营养体亲和比例为93.3%,不亲和比例为6.7%;交配型 MAT1-2对峙培养时,营养体亲和比例为86.7%,不亲和比例为13.3%营养亲和性结果表明,同种交配型之间配对组合表现出较好的亲和性,但是原基发育的很少;而不同交配型 MAT1-1和MAT1-2之间杂交组合的营养亲和比例相对较低,亲和性良好的组合均发育成原基,说明具有不同交配型且营养亲和性良好的蛹虫草菌株,更容易发育原基,形成子实体。表3,图5

表3 不同菌株间营养体亲和性Table 3 Vegetative compatibility of different strains
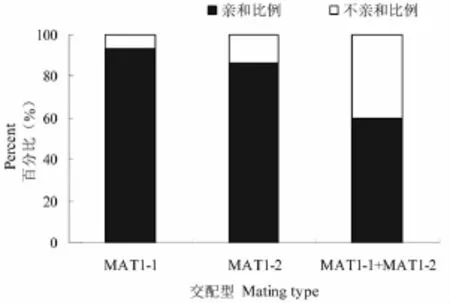
图5 交配型组合的亲和性Fig.5 Compatibility of combination between mating types
3 讨 论
蛹虫草为典型的二级异宗交配系统,由两个不同交配型的发生亲和的同核体配合后形成异核体,异核体发育形成子实体,而缺失了交配型基因的同核体不能完成有性生活史,理论上不能形成正常的蛹虫草子实体[10]。由此推断大部分蛹虫草子实体应该是异核体而非同核体,形成异核体时两种交配型基因随机组合,蛹虫草交配型基因是互不同源的,两种交配型比例应为1∶1。研究检测结果表明所有供试菌株中只含有一种交配型(MAT1-1或MAT1-2),没有两种交配型共存的情况,并通过卡方检验表明不同群体子囊孢子交配型分离比例符合 1∶1,说明供试的蛹虫草群体中交配型均没有出现分离现象。
真菌营养体亲和性鉴定通过抗衡区的产生、隐性的营养缺陷型或色素标记的互补作用以及菌系的生长特性来观察测定[7]。相同交配型单子囊孢子菌株间产生子实体或菌株自身产生子实体的没有不同交配型菌株配对形成子实体的产量高,且相同交配型菌株的配对组合及单子囊孢子菌株自交产生的子实体通常情况下不能产生子囊和子囊孢子[13]。研究通过试验表明交配系统与营养亲和性之间具有一定的潜在联系,同种交配型菌株之间表现亲和性高于不同交配型菌株之间;同种交配型菌株之间也有营养不亲和现象,这可能是菌株交配型相同但来源不同,亲缘关系较远导致。另外,同种交配型之间对峙培养表现出较好的亲和性,但是由于缺乏遗传因子(如交配型),从而导致鲜有原基发育;值得注意的是不同交配型之间其营养亲和性比例较低;但具有营养亲和性的不同交配型菌种之间,大多可以发育原基并形成子实体。
4 结 论
所有供试群体中,单子囊菌株只具有一种交配型(MAT1-1或MAT1-2),交配型 MAT1-1和 MAT1-2分离比例符合 1∶1,且交配型未出现偏分离现象。蛹虫草交配系统与营养亲和性之间具有一定的潜在相关性,相同交配型菌株之间的营养亲和性为90%,较高;不同交配型菌种之间的营养亲和性比例60%,较低。相同交配型菌株比不同交配型菌株营养体之间更普遍存在营养亲和性;但如果不同交配型菌株具有营养亲和性,则更有利于原基发育及子实体形成。
(
)
[1]Zhou,X.,Gong,Z.,Su,Y.,Lin,J.,&Tang,K.(2009). Cordyceps fungi:natural products,pharmacological functions and developmental products.Journal of Pharmacy&Pharmacology,61(3):279-291.
[2]孟泽彬,陈林会,朝近雨,等.蛹虫草化学活性成分的研究[J].分子植物育种,2015,(9):2 147-2 154. MENG Ze-bin,CHEN Li-hui,,CHAO Jin-yu,et al.(2015).Research advances of the chemical active constituents of Cordyceps militaris[J].Molecular Plant Breeding,(9):2,147-2,154.(in Chinese)
[3]张姝,张永杰,SHRESTHA Bhushan,等.冬虫夏草菌和蛹虫草菌的研究现状、问题及展望[J].菌物学报,2013,32(4):577-597. ZHANG Shu,ZHANG Yong-jie,Shrestha B,et al.Ophiocordyceps sinensis and Cordyceps militaris:research advances,issues and perspectives[J].Mycosystema,32(4):577-597.(in Chinese)
[4]胡秀彩,吕爱军.蛹虫草菌退化的探讨[J].中国食用菌,2015,34(1):1-3. HU Xiu-cai,Lü Ai-jun.(2015).Discussion of the Degenerate Strains of Cordyceps militaris[J].Edible Fungi of China,34(1):1-3.(in Chinese)
[5]郑壮丽,黄春花,梅彩英,等.韩日畴蛹虫草国内外研究的新进展环境[J].昆虫学报,2011,33(2):225-233. ZHENG Zhuang-li,HUANG Chun-hua,MEI Cai-ying,et al.(2011).Current advance on the research of Cordyceps militaris[J].Journal of Environmental Entomology,33(2):225-233.(in Chinese)
[6]翟景波,滕利荣,黄宝玺.高产蛹虫草菌株的选育[J].中国生物制品学杂志,2011,24(10):1 230-1 232. ZAI Jing-bo,TENG Li-rong,HUANG Bao-xi.(2011). The breeding of high-yield Cordyceps militaris strains[J].Chinese Journal of Biologicals,24(10):1,230-1,232.(in Chinese)
[7]张星耀,陈海燕,梁军,等.金黄壳囊孢菌的培养性状和营养体亲和性[J].西北农林科技大学学报,2007,35(3):99-105. ZHANG Xing-yao,CHEN Hai-yan,LIANG Jun,et al.(2007).Cultural morphology and vegetative compatibility of Cytospora chrysosperma isolates[J].Journal of Northwest A&F U-niversity,35(3):99-105.(in Chinese)
[8]李美娜,吴埘军,李春燕,等.人工栽培蛹虫草退化现象的分子分析[J].菌物系统,2003,22(2):277-282. LI Mei-na,WU Shi-jun,LI Chun-yan,et al.(2003). Molecular analysis of degeneration of artificial planted Cordyceps militaris[J].Mycosystema,22(2):277-282.(in Chinese)
[9]Shrestha,B.,Kim,H.K.,Sung,G.H.,Spatafora,J.W.,&Sung,J.M.(2004).Bipolar heterothallism,a principal mating system ofcordyceps militaris in vitro.Biotechnology&Bioprocess Engineering,9(6):440-446.
[10]高新华.蛹虫草(Cordyceps militaris)的交配型研究[J].食用菌学报,2008,15(1):6-10. GAO Xin-hua.(2008).Mating System of Cordyceps militaris[J].Acta Edulis Fungi,15(1):6-10.(in Chinese)
[11]Yokoyama,E.,Yamagishi,K.,&Hara,A.(2003).Structures of the mating-type loci of cordyceps takaomontana.Applied&Environmental Microbiology,69(8):5,019-5,022.
[12]Peng,Z.,Xia,Y.,Xiao,G.,Xiong,C.,Xiao,H.,&Zhang,S.,et al.(2011).Genome sequence of the insect pathogenic fungus Cordyceps militaris,a valued traditional chinese medicine.Genome Biology,12(11):287-302.
[13]Zhang,G.,&Liang,Y.(2013).Improvement of fruiting body production in cordyceps militaris by molecular assessment. Archives of Microbiology,195(8):579-585.
[14]谭琦,蔡涛,汪虹,等.蛹虫草无性孢子的交配型基因类型的分子鉴定[J].上海农业学报,2011,27(3):5-8. TAN Qi,CAI Tao,WANG Hong,et al.(2011).Molecular identification of mating-type genes of asexual spores from Cordyceps militaris[J].Acta Agriculturae Shanghai,27(3):5-8.(in Chinese)
[15]梁月,张国珍,安沫平,等.蛹虫草子囊孢子萌发及其后代群体培养性状观察[J].菌物学报,2005,24(4):525-532. LIANG Yue,ZHANG Guo-zhen,AN Mo-ping,et al.(2005).Cordyceps militaris:ascospore germination and cultural characteristics of progeny population[J].Mycosystema,24(4):525-532.(in Chinese)
[16]李沛利,秦芸,严吉明,等.四川省核盘菌的营养体亲和性[J].四川农业大学学报,2010,28(3):324-327.LI Pei-li,QIN Yun,YAN Ji-ming,et al.(2010).Vegetative Compatibility of Sclerotinia sclerotiorum from Sichuan Province[J].Journal of Sichuan Agricultural University,28(3):324-327.(in Chinese)
Preliminary Study on Mating System and Vegetative Compatibility in Cordyceps militaris
JIAO Zi-wei1,Nuermaimaiti1,LIANG Yue2,YANG Xiao-rong1
(1.College of Chemistry and Biological Sciences,Yili Normal University,Yining Xinjiang 835000,China;2.College of Plant Protection,Shenyang Agricultural University,Shenyang 110866,China)
【Objective】Cordyceps militaris is a kind of medical and edible fungus,which is a typically heterothallic ascomycetous fungus.This project aims to conduct a preliminary investigation of the correlation of mating system and vegetative compatibility.【Method】In this paper,different single-spore groups were obtained through single ascospore isolation from the different fruiting bodies of C.militaris.The isolated strains were cultured and mycelial DNA was extracted.Design specific primers were designed for PCR amplification of mating type gene to identify the mating types and segregation ratio.Some strains were randomly selected to detect vegetative compatibility between the same and/or the different mating type strains using the confrontation training method between paired cultures.【Result】The results showed that all detected strains were either mating type MAT 1-1 or MAT 1-2.Difference sampling groups were determined and thereafter the segregation ratio was analyzed by Chi-square,which indicated that the ratio of two mating types was consistent with a theoretical segregation ratio of 1∶1.The strains with the different mating type were randomly chosen to investigate the vegetative compatibility using crossing culture between paired strains.The results indicated that the strains were divided into four vegetative compatibility groups,which indicated the potentially genetic differentiation.Vegetative compatibility between the strains between the same mating type(s)was higher(90%)than those between the different mating types(60%).【Conclusion】All the tested single ascospores only showed one of mating types and the population sample did not present segregation distortion.The correlation of mating system and vegetative compatibility in C.militaris was possible.The vegetative compatibility of strain in combination with the same mating type was more widespread than that of different mating types.Thereafter,the combination of the strains with different mating types and vegetative compatibility suggested utilization for development of stroma and formation of fruiting body.
Cordyceps militaris;Single ascospore;mating type;compatibility;vegetative compatibility group
S188+.2
A
1001-4330(2016)09-1708-07
10.6048/j.issn.1001-4330.2016.09.019
2016-02-23
新疆维吾尔自治区科技支疆项目(201491153);辽宁省高等学校优秀科技人才支持计划(LR2015058)
焦子伟(1973-),男,河南人,副教授,博士,研究方向为微生物生态,(E-mail)741285332@qq.com
(Cotresponding author):努尔买买提·伊力亚斯(1971-),男,新疆伊宁人,副教授,博士研究生,研究方向为植物分类,(E-mail)2696232271@qq.com梁月(1978-),男,辽宁人,教授,博士,研究方向为真菌生物学及植物真菌病害,(E-mail)yliang@syau.edu.cn
Fund project:Supported by The Xinjiang Uygur Autonomous Region science and technology support project(201491153)and Liaoning Province university outstanding scientific and technical personnel support program(LR2015058).
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2021年12期)2021-11-30
四川蚕业(2021年1期)2021-02-12
昆虫学报(2020年6期)2020-08-06
昆虫学报(2020年1期)2020-03-03
世界热带农业信息(2019年9期)2019-01-05
阿来研究(2018年1期)2018-07-10
山东农业科学(2017年5期)2017-06-05
上海农业学报(2017年3期)2017-04-10
数字技术与应用(2016年10期)2017-04-01
