
紫杉醇关键酶基因TwDBAT在平菇中的遗传转化及表达
2017-02-05 20:22康俊
江苏农业科学 2016年10期

doi:10.15889/j.issn.1002-1302.2016.10.023
摘要:通过构建含平菇内源gpd启动子、红豆杉中TwDBAT基因的真核表达载体pgFvs-TwDBAT,建立高效的平菇原生质体制备与再生体系;采用PEG介导法,将真核表达载体pgFvs-TwDBAT、pgFvs-hph共转化高温型平菇菌株F7E原生质体,获得可以异源表达TwDBAT基因的平菇菌株。PCR和Southern杂交鉴定结果表明,TwDBAT基因整合进平菇基因组中;经继代培养,TwDBAT基因在转化子中可稳定遗传。RT-PCR结果表明,转化子在mRNA水平上转录了TwDBAT基因。HPLC结果显示,TwDBAT酶可以催化底物10-去乙酰巴卡亭Ⅲ(10-DAB)。
关键词:平菇;紫杉醇;紫杉醇前体物质;关键酶基因;TwDBAT基因;遗传转化;基因表达
中图分类号: S188文献标志码: A文章编号:1002-1302(2016)10-0098-05
收稿日期:2016-03-31
基金项目:内蒙古自治区高等学校科学研究项目(编号:NJZC14383)。
作者简介:康俊(1982—),女,内蒙古乌兰察布人,硕士,讲师,主要从事植物病理及遗传育种的教学研究工作。E-mail:kang_jun@163.com。紫杉醇是从红豆杉属植物中分离出的1种二萜类生物碱,因具有广谱抗癌活性和独特抗癌机制而受到人们青睐[1-3]。自紫杉醇的抗癌功效和作用机制被确定以来,特别是获批准用于临床治疗卵巢癌、乳腺癌,以及紫杉醇生物合成过程的某些关键酶的发现、克隆和高效表达,使得实现紫杉醇高效、稳定生产成为研究热点[3-4]。
中国特有濒危植物南方红豆杉中的10-去乙酰巴卡亭Ⅲ-10β-乙酰转移酶(10-deacetylbaccatin Ⅲ-10β-O-acetyl transferase,DBAT)是红豆杉属植物紫杉醇生物合成途径中的一个重要酶类[5-6],其作用是将10-去乙酰巴卡亭Ⅲ转换成巴卡亭Ⅲ,而巴卡亭Ⅲ不仅是生物合成紫杉醇的直接前体,也是紫杉醇生物合成途径中的最后1个二萜类中间体,更是目前化学半合成紫杉醇的重要前体化合物,在缓解紫杉醇资源短缺问题中起着不可替代的作用。本研究在优化制备高浓度的平菇菌丝原生质体的基础上,采用聚乙二醇(PEG)介导的方法将含有平菇内源性gpd-fvs启动子和TwDBAT基因(南方红豆杉中的DBAT基因)的真核表达载体遗传转化高温型平菇突变菌株F7E原生质体,以获得能够稳定表达TwDBAT基因的转基因平菇菌株,这将为紫杉醇生物合成途径中其他重要酶基因遗传转化平菇提供技术路线和依据,并为平菇组合表达紫杉醇或紫杉醇前体物质研究奠定基础。
1材料与方法
1.1菌株和质粒
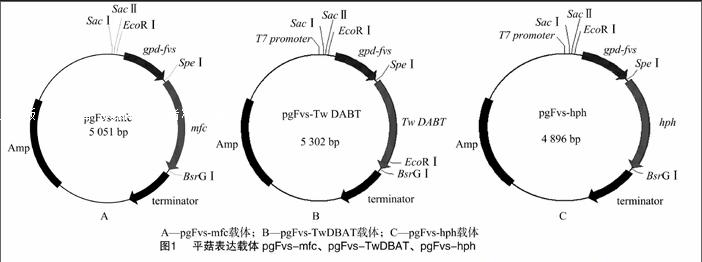
高温型平菇突变菌株F7E,其菌丝体可在高温下正常生长,并能够在20 ℃以上正常发育形成子实体、生物学效率较高的平菇突变体,为笔者所在实验室诱变筛选出的突变株[7];真核表达载体pgFvs-mfc[8]、克隆载体pGEM-TwDBAT[9]均为笔者所在实验室构建保存;表达载体pgFvs-TwDBAT(含有平菇内源性gpd-fvs启动子和TwDBAT基因)、表达载体 pgFvs-hph(含有平菇内源性gpd-fvs启动子和hph基因)为本研究构建。
1.2酶与试剂
ExTaq酶、dNTP、Taq DNA聚合酶、PrimeScriptTM RT-PCR Kit、EcoRⅠ、HindⅢ限制性内切酶,购自宝生物工程(大连)有限公司;DNA凝胶回收试剂盒,购自天根生化科技(北京)有限公司;pGEM-T Easy载体,购自Promega公司;Plant RNAout提取试剂盒,购自北京天恩泽基因科技有限公司;溶壁酶,购自广东省微生物研究所;地高辛试剂盒,购自Roche公司;PCR引物由生工生物工程(上海)股份有限公司合成;DNA测序由华大基因完成;甘露醇(MMC) buffer(50 mL):25 mL 1 mol/L 甘露醇溶液,12.5 mL 0.2 mol/L Maleate buffer,1.25 mL 1 mol/L CaCl2,11.25 mL无菌去离子水;0.2 mol/L Maleate buffer(马来酸缓冲液):将0.2 mol/L马来酸、0.2 mol/L 马来酸钠按1 ∶2比例混合,调节pH值至5.5,121 ℃ 灭菌15 min;1 mol/L CaCl2:称取7.35 g CaCl2,用ddH2O定容至50 mL,过滤除菌;山梨醇(100 mL):称取 18.2 g 山梨醇,分别加入1 mL 1 mol/L Tris-Cl(pH值7.5)、2.5 mL 1 mol/L CaCl2,加ddH2O定容至100 mL,121 ℃灭菌15 min;PEG buffer(20 mL):5.0g PEG 4000,0.2 mL 1 mol/L Tris-Cl (pH值为7.5),0.5 mL 1 mol/L CaCl2,加ddH2O至20 mL,加热至60 ℃过滤除菌,用小管分装后于-40 ℃备用;其他试剂均为国产分析纯。
1.3培养基
PDA固体培养基:200 g去皮马铃薯,20 g葡萄糖,10 g琼脂粉,加水至1 L,常规方法配制。PDB液体培养基:在PDA固体培养基配方的基础上去掉琼脂粉。平菇原生质体再生培养基:在PDA培养基的基础上分别加入蔗糖、甘露醇、氯化钠、氯化钾,使其终浓度均为0.6 mol/L。
1.4引物
根据TwDBAT基因序列,设计2对PCR特异引物,用来鉴定阳性转化子基因组DNA中是否存在TwDBAT基因,引物由上海捷瑞生物工程有限公司合成。鉴定引物对:13F1(5′-TTAGAGAGAGTGATGGTGGCT-3′)、13R1(5′-TTCTTGGGAGGTCGTATGA-3′),预计扩增产物为1 184 bp;13F2(5′-ACATCCATCCTCTGGTGGTTCA-3′)、13R2(5′-CTTCATCAAATCCCAATCGCCT-3′),预计扩增产物为699 bp。
1.5试验方法
1.5.1平菇表达载体pgFvs-TwDBAT、pgFvs-hph的构建质粒抽提、限制酶酶切、DNA片段回收及连接按试剂说明书进行。用SpeⅠ、BsrGⅠ分别双酶切质粒pGEM-TwDBAT、pgFvs-mfc、pBgGI-hph,然后经琼脂糖凝胶电泳后割胶,分别回收大片段、小片段,并用T4DNA连接酶分别连接大小片段,将反应产物转化DH5α 感受态细胞,选择阳性克隆菌进行菌落PCR、双酶切及测序。
1.5.2平菇原生质体制备与再生收集液体PDB培养基中的平菇菌丝,分别用无菌水、稳渗剂冲洗3次,平菇原生质体制备与再生参照Kuo等的方法[10],溶壁酶酶液浓度为15%,所得原生质体最后用0.6 mol/L KCl保存。
1.5.3培养方式对原生质体再生的影响将原生质体按以下3种方式接种培养,以分析培养方式对原生质体再生的影响,本试验平行进行3次。(1)直接涂布平板培养法:用液体再生培养基将新制备的原生质体悬液稀释到106个/mL左右,取0.1 mL直接涂布在含2%琼脂的再生培养基上,25 ℃培养7 d。(2)预培养法:用液体再生培养基将新制备的原生质体悬液稀释到106个/mL左右,25 ℃预培养24 h后,取 0.1 mL 涂布于含2%琼脂的再生培养基上,25 ℃培养7 d。(3)双层平板培养法:先在培养皿中倒1层含2%琼脂的再生培养基,冷凝后,将0.1 mL原生质体悬液(106个/mL)与预热的含1%琼脂的半固体再生培养基于40 ℃混匀,倾注于固体再生培养基平板上,25 ℃培养7 d。
1.5.4平菇菌丝体对潮霉素的最低敏感浓度将活化后的供试菌株接种于含有不同终浓度潮霉素的PDA平板(0、25、50、75、100、125、150 μg/mL)中,于25 ℃恒温培养,7 d后观察结果,测量菌落直径,选择完全抑制供试菌生长的最低潮霉素浓度。
1.5.5平菇原生质体对潮霉素的最低敏感浓度将平菇原生质体稀释到105~106个/mL左右,取0.1 mL直接涂布于含不同终浓度潮霉素(0、10、20、30、40、50、60 μg/mL)的再生培养基平板上,于25 ℃恒温培养,7 d后观察结果,测量菌落直径,选择完全抑制供试菌原生质体再生的最低潮霉素浓度。
1.5.6PEG法转化平菇菌丝原生质体将新鲜平菇原生质体悬浮于MMC buffer中,使其终浓度为1×108个/mL;在分装有50 μL原生质体的管中,分别加入5 μg质粒pgFvs-TwDBAT、pgFvs-hph,另设1管不加质粒作为对照;每管中各加入12.5 μL PEG buffer,冰浴20 min;加入0.5 mL PEG buffer,混匀后于28 ℃温育15 min;加入1.0 mL STC buffer;小心混匀后将其转入10 mL液体再生培养基中,25 ℃预培养 48 h;预培养后于4 000 r/min离心10 min,弃上清,用1.0 mL液体再生培养基将原生质体重新悬浮。按200~250 μL/皿的量涂布在PDSA培养基上(含潮霉素HmB,浓度为 40 μg/mL),25 ℃培养7 d;待原生质体萌发成再生菌落后,在其上覆盖1层含0.8%琼脂的固体PDSA(含潮霉素HmB,浓度为75 μg/mL);待夹层培养基上长出再生菌落,即为阳性转化子。
1.5.7阳性转化子的挑选固体平板培养初筛:用无菌牙签挑取转化平板上菌落的小部分,转接于HmB浓度为 75 μg/mL 的PDSA平板上,培养10 d以上观察各阳性转化子的生长情况,同时设1管阴性对照(未转化平菇原生质体)。选取接种后10 d内可以连续生长的菌丝体,作为阳性转化子,待做PCR鉴定。
液体摇瓶培养初筛:将转化平板上长出的阳性转化子的剩余部分接种于3 mL HmB浓度为75 μg/mL的PDSB试管中,同时设1管阴性对照(未转化平菇原生质体),180 r/min、25 ℃摇床培养5 d。丢弃肉眼不可见菌体的试管,收集有菌试管内菌体(阳性转化子),待做PCR鉴定。
1.5.8阳性转化子的PCR和RT-PCR鉴定以阳性转化子的菌丝基因组DNA、cDNA为模板,用2对引物(13F1、13R1与13F2、13R2)进行PCR、RT-PCR扩增TwDBAT基因(1 184 bp)及其片段(699 bp)。以表达载体pgFvs-TwDBAT为阳性对照,以未转化平菇基因组DNA、cDNA为阴性对照。PCR扩增程序:94 ℃ 4 min;94 ℃ 60 s,52 ℃ 60 s,72 ℃ 2 min,35个循环;72 ℃ 10 min。用TaKaRa凝胶回收试剂盒回收DNA片段,并由华大基因公司进行测序。将测序结果与NCBI数据库中公布的DBAT基因序列进行对比分析。
1.5.9转化子的Southern Blot分析用CTAB法提取PCR、RT-PCR鉴定呈阳性的菌丝基因组DNA。每个菌株样品取30~50 μg基因组DNA,选用限制性内切酶Hind Ⅲ酶切过夜。酶切完全后,用0.8%琼脂糖凝胶电泳使各片段按照分子大小依次分开,利用向下毛细管印迹法将酶切片段由凝胶转移到尼龙膜上。Southern杂交中的探针标记、杂交等按照Roche公司生产的地高辛标记检测试剂盒操作方法进行。
1.5.10转化子菌丝体遗传稳定性分析取Southern杂交鉴定正确的转化子菌丝体,转接于无潮霉素选择压力的PDA固体培养基上继代培养5次后,再次转入含有75 μg/mL潮霉素的PDA平板上,25 ℃培养10 d。挑选具有抗潮霉素抗性、可以正常生长的转化子,提取菌丝体基因组DNA,进行PCR鉴定。
1.5.11转化子菌丝体TwDBAT酶活性研究按Patel等的方法[11-12],并略加修改。将转化子菌丝体用液氮研磨后,用TE缓冲液于4 ℃浸提60 min;6 000 r/min离心10 min,上清液中加入硫酸铵,使硫酸铵饱和度为90%,充分搅拌后,4 ℃放置过夜;12 000 r/min离心20 min,将沉淀物转入透析袋中,放入20 mmol/L Tris-HCl缓冲液(pH值7.5)中透析过夜,其间多次更换透析液。透析后的蛋白酶液与5 mmol/L MgCl2、10-DAB (400 μmol/L)、acetyl CoA (400 μmol/L)混合,31 ℃反应3 h。将反应液用50 mL乙酸乙酯在30 ℃恒温下振摇提取30 min,反复提取3次。合并乙酸乙酯,减压蒸干,用1 mL流动相溶解残留物,微孔滤膜(0.45 μm)滤过,待测定。HPLC条件:色谱柱Hypersil ODS C18(250 mm×4.6 mm,5 μm),流动相水 ∶甲醇 ∶乙腈=70 ∶20 ∶10,流速1.5 mL/min,平衡时间5.0 min,UV检测波长227 nm,进样量 20 μL。
2结果与分析
2.1平菇表达载体pgFvs-TwDBAT、pgFvs-hph的构建
将PCR鉴定和SpeⅠ、BsrGⅠ双酶切鉴定与预期结果一致的阳性克隆菌进行测序。经BLAST比对,测序结果与公布的DBAT、hph基因的同源性为99.9%,说明表达载体构建成功,分别命名为pgFvs-TwDBAT、pgFvs-hph(图1)。
2.2平菇原生质体制备与再生率
综合不同因素对平菇原生质体制备的影响,选取各因素中的最优参数,优化平菇原生质体制备条件。研究发现:液体培养5 d的平菇菌丝,以0.6 mol/L KCl作为稳渗剂,加入15%溶壁酶,在25 ℃下酶解平菇菌丝3 h时,分离得到的原生质体效果最佳,原生质体产量为5×107~8×108个。
分别采用预培养法、直接涂抹法、双层培养法进行试验。结果表明:双层培养法通过将原生质体包埋于半固体培养基中,倾倒于再生培养基上,机械损伤较小,有利于原生质体的再生,但操作比较繁琐,且半固体培养基的温度难以控制,稍低会使培养基凝固,过高容易使原生质体失活;直接涂抹法操作简单方便,但会带来机械损伤,破坏原生质体;预培养法兼具直接涂抹法、双层培养法的优点,采用预培养法效果比较好,原生质体再生率可达6.32%。
2.3平菇菌丝体、原生质体对潮霉素的最低敏感浓度
将高温型平菇突变菌株F7E接种在含不同浓度潮霉素的PDA平板上培养。试验结果表明,随着潮霉素浓度的增加,菌丝生长速度、生长势逐渐降低;当潮霉素浓度为 50 μg/mL 时,菌丝在平板上可以微弱生长;当潮霉素浓度为75 μg/mL时,菌丝无法生长。因此可以确定,平菇突变菌株F7E菌丝对潮霉素的最低敏感浓度为75 μg/mL。
制备高温型平菇突变菌株F7E的原生质体,将原生质体稀释到106个/mL,取0.1 mL原生质体直接涂布于含不同终浓度潮霉素再生培养基平板上培养。试验结果表明,随着潮霉素浓度的增加,原生质体的再生率降低;当潮霉素浓度为20 μg/mL时,原生质体的再生率为0.02%;当潮霉素浓度为30 μg/mL时,原生质体的再生率为0。因此,确定平菇突变菌株F7E原生质体对潮霉素的最低敏感浓度为30 μg/mL。
2.4PEG法转化平菇菌丝原生质体
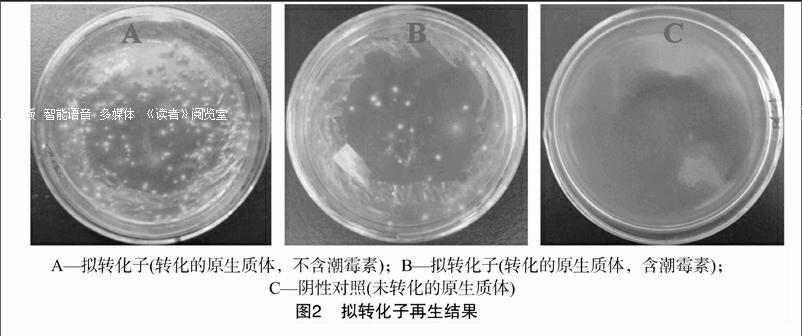
利用PEG介导方法,将质粒pgFvs-TwDBAT、pgFvs-hph共转化平菇原生质体,经潮霉素抗性平板筛选后,在培养基上长出白色菌落;转化后,在含有潮霉素的再生筛选培养基上,只有成功转入质粒pgFvs-TwDBAT、pgFvs-hph的共转化子或只转入质粒pgFvs-hph的单转化子才能正常生长(图2)。
2.5Southern杂交
从筛选培养基上挑出拟转化子,经PCR鉴定筛选转化子。通过CTAB法提取转化子基因组DNA。酶切后的转化子基因组DNA经转膜,与地高辛标记的TwDBAT探针杂交后,经分析可知:6个转化子基因组DNA和阳性质粒都可与TwDBAT探针杂交在膜上显出特异的条带,而泳道7、空白对照非转基因平菇菌株则没有出现与TwDBAT探针杂交的信号带(图3)。这表明TwDBAT基因已经整合到6个转化子(t2、t4、t5、t7、t11、t12)的染色体基因组上,而转化子t20则没有被整合,推断可能是由于PCR的假阳性造成的。对杂交条带的数量、信号强度、出现位置的进行进一步分析可知,除转化子t7(泳道4)仅有1条微弱的杂交信号外,其余转化子一般都有2~3条杂交信号的条带,其信号强度和出现的位置也各不相同,其中转化子t5、t11、t12(泳道3、5、6)杂交信号条带位置和信号强度几乎一致,它们可能为同一株转化子。因此,根据PCR、RT-PCR结果和Southern Blot分析结果,可以得出TwDBAT基因是成功地以多拷贝随机插入到转化子染色体DNA中的。
2.6转化子的RT-PCR鉴定结果
将PCR鉴定和Southern Blot分析正确的阳性转化子(t2、t4、t5、t7、t11、t12)菌丝体提取RNA,进行RT-PCR。由图4可知,在对照CK-未扩增出目的条带的情况下,各转化子均有与阳性对照大小一致的扩增条带出现。这表明在转基因平菇转化子中,TwDBAT基因已在mRNA转录水平上得到表达。
2.7转化子菌丝体遗传稳定性分析
将PCR鉴定和Southern Blot鉴定正确的阳性转化子(t2、t4、t5、t7、t11、t12)菌丝体分别转接10次,进行转化子菌丝体遗传稳定性研究。由图5可见,转化子的S10代菌丝体仍对潮霉素具有一定抗性,可以在潮霉素抗性平板上正常生长。对S10代的转化子进行PCR鉴定可知:6个转化子中都扩增出与表达载体pgFvs-TwDBAT相同的目的基因TwDBAT特异条带,大小约为1 184 bp,与预期理论目的基因PCR扩增产物大小相符。由此初步推断,平菇转化子以转接菌丝方式,经过10次菌丝转接繁殖后,其基因组DNA仍含有外源TwDBAT基因。
2.8转化子TwDBAT酶活性检测
将提取的转化子蛋白酶提取物与底物10-DAB混合反应后,利用HPCL方法对乙酸乙酯提取物进行检测分析。
由图6可知,当添加底物10-DAB时,对照非转化子和转化子提取物反应液HPLC色谱图存在一定的差异,转化子提取物在7.65 min时出现了1个明显的分离小峰(图 6-B),而该峰与产物巴卡亭Ⅲ标准样品的保留时间一致;而在同等条件下,对照非转化子并无此峰出现(图6-A),这种差异可能是转化子表达外源TwDBAT酶蛋白催化底物 10-DAB 而生成产物巴卡亭Ⅲ所引起的。
3讨论
随着紫杉醇代谢途径的逐步阐明和相关酶基因的成功克隆[12-14],通过代谢工程,在微生物中组合表达紫杉醇生物合成重要酶基因,利用“微生物工厂”来直接生产紫杉醇,或者生产中间物质,进而再通过化学半合成的方法生产紫杉醇将是具有广阔前景的、较为可行的途径。目前,已在大肠杆菌和酵母中进行紫杉醇组合表达的研究[4,6,15-16],并生成了紫杉烷类化合物,实现了由非紫杉烷类产生菌获得紫杉烷类化合物的突出研究成果,使人们看到了利用生物技术生产紫杉醇前体的广阔前景。
PEG介导的原生质体转化法,操作简单,是食用菌遗传转化中应用最多的一种方法,已在双孢蘑菇、糙皮侧耳、云芝、灵芝、草菇、灰盖鬼伞、平菇[17-18]等食用菌中成功应用。本研究成功采用PEG介导法,首次将含平菇内源gpd启动子和红豆杉中紫杉醇生物合成途径中的TwDBAT基因的真核表达载体pgFvs-TwDBAT转入高温型平菇菌株F7E原生质体中,其转化结果与Kuo等的PEG原生质体介导法[10,19]一致,都表明PEG原生质体介导法可成功应用于平菇遗传转化研究。此外,本试验中采用的内源性gpd启动子遗传转化平菇结果与Cheng等研究结果[8,10,20]一致,说明内源性gpd启动子有利于受体细胞调控因子的识别,可以减少甲基化,促进外源基因整合到染色体上;平菇内源性gpd启动子是平菇遗传转化、有效控制外源功能基因正确表达的一个重要控制元件。
遗传转化成功后,外源基因能否在转基因植株的后代中稳定遗传和表达是基因工程技术研究中备受关注的问题,外源基因整合到受体基因组后产生的遗传效应是多样的,在受体植株中的遗传行为也是非常复杂的。杨飞芸等在进行转基因草菇菌丝继代遗传稳定性检测时发现,转基因草菇菌丝中afp基因已经丢失[21-22],可能因为草菇菌丝是多核的,在多次继代接种过程中接种了不含目的基因的菌丝,造成细胞中外源基因的丢失。在本研究中,对平菇转化子进行遗传稳定性试验发现,该平菇转化子可稳定遗传外源基因TwDBAT, 这与
李巍等的bFGF基因遗传转化平菇的研究结果[23]一致。
本研究首次成功建立的平菇异源表达红豆杉中紫杉醇生物合成途径上的TwDBAT基因遗传转化及表达研究,将为紫杉醇生物合成途径中的其他重要酶基因(例如GGPP合酶、紫杉烯合酶、紫杉烯5α-羟基化酶、紫杉烯醇5α-乙酰氧化基转移酶、紫杉烷10β-羟基化酶、紫杉烷7β-羟基化酶基因等)遗传转化平菇提供技术路线和依据,并为平菇组合表达紫杉醇或紫杉醇前体物质研究奠定基础。
参考文献:
[1]沈征武,吴莲芬. 紫杉醇研究进展[J]. 化学进展,1997,9(1):1-13.
[2]Stierle A,Strobel G,Stierle D. Taxol and taxane production by Taxomyces andreanae,an endophytic fungus of Pacific yew[J]. Science,1993,260(5105):214-216.
[3]Stumm S,Meyer A,Lindner M,et al. Paclitaxel treatment of breast cancer cell lines modulates Fas/Fasl ligand expression and induces apoptosis which can be inhibited through the CD40 receptor[J]. Oncology,2004,66(2):101-111.
[4]Stone R. Surprise!A fungus factory for taxol?[J]. Science,1993,260(515):154-155.
[5]Danishefsky S J,Master J J,Young W B,et al. Total synthesis of Baccatin Ⅲ and taxol[J]. Journal of the American Chemical Society,1996,118(12):2843-2859.
[6]Walker K,Croteau R. Molecular cloning of a 10-deacetylbaccatin Ⅲ-10-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli[J]. Proceedings of the National Academy of Sciences of the United States of America,2000,97(2):583-587.
[7] 韩飞. 高温节能型平菇选育及其表达外源基因研究[D]. 广州:华南农业大学,2011.
[8]Cheng S,Yang P,Guo L,et al. Expression of multi-functional cellulase gene mfc in Coprinus cinereus under control of different basidiomycete promoters[J]. Bioresource Technology,2009,100(19):4475-4480.
[9]程抒劼,黄仕杰,郭丽琼,等. 南方红豆杉10-去乙酰巴卡亭Ⅲ-10-乙酰转移酶基因的克隆与生物信息学分析[J]. 生物技术通报,2011(1):107-112.
[10]Kuo C Y,Chou S Y,Huang C T. Cloning of glyceraldehyde-3-phosphate dehydrogenase gene and use of the gpd promoter for transformation in Flammulina velutipes[J]. Applied Microbiology and Biotechnology,2004,65(5):593-599.
[11]Patel R N,Banerjee A,Nanduri V. Enzymatic acetylation of 10-deacetylbaccatin Ⅲ to baccatin Ⅲ by C-10 deacetylase from Nocardioides luteus SC 13913[J]. Enzyme and Microbial Technology,2000,27(6):371-375.
[12]Guo B,Kai G,Gong Y,et al. Molecular cloning and heterologous expression of a 10-deacetylbaccatin Ⅲ-10-O-acetyl transferase cDNA from Taxus x media[J]. Molecular Biology Reports,2007,34(2):89-95.
[13]韩飞,康林芝,郭丽琼,等. 南方红豆杉GGPP合酶基因的克隆与生物信息学分析[J]. 食品与生物技术学报,2011,30(5):728-733.
[14]Walker K,Long R,Croteau R. The final acylation step in taxol biosynthesis:cloning of the taxoid C13-side-chain N-benzoyltransferase from Taxus[J]. Proceedings of the National Academy of Sciences of the United States of America,2002,99(14):9166-9171.
[15]Zhang L H,Wang X H,Qiu D Y,et al. Microbial transformation of 10-deacetyl-7-epitaxol and 1-β-hydroxybaccatin Ⅰ by fungi from the inner bark of Taxus yunnanensis[J]. Journal of Natural Products,1998,61(4):497-500.
[16] Wang W,Kong J Q,Meng C,et al. Studies on combinatorial biosynthesis of taxadiene in Escherichia coli[J]. Journal of Chinese Pharmaceutical Sciences,2005,40(18):1428-1431.
[17]郭丽琼,林俊芳,熊盛,等. 抗冷冻蛋白基因遗传转化草菇的研究[J]. 微生物学报,2005,45(1):39-43.
[18]Han F,Liu Y,Guo L Q,et al. Heterologous expression of the immunomodulatory protein gene from Ganoderma sinense in the basidiomycete Coprinopsis cinerea[J]. Journal of Applied Microbiology,2010,109(5):1838-1844.
[19]Maehara T,Yoshida M,Ito Y,et al. Development of a gene transfer system for the mycelia of Flammulina velutipes Fv-1 strain[J]. Bioscience Biotechnology and Biochemistry,2010,74(5):1126-1128.
[20] Punt P J,Dingemanse M A,Kuyvenhoven A,et al. Function elements in the promoter region of the Aspergillus nidulans gpd A gene encoding glyceraldehydes-3-phosphate dehydrogenase[J]. Gene,1990,93(1),101-109.
[21] 杨飞芸. 转基因草菇的生物学特性及其后代遗传稳定性研究[D]. 广州:华南农业大学,2005.
[22] 赵风云. 转多功能纤维素酶基因(mfc)草菇的稳定性分析和高产菌株的选育[D]. 广州:华南农业大学,2010.
[23]李巍,刘秀明,李洪志,等. 农杆菌介导的碱性成纤维细胞生长因子(bFGF)转化平菇的研究[J]. 菌物学报,2011,30(1):46-53.兰英,柳敏,严铸云,等. 不同地理种源丹参组培快繁及再生苗性状差异比较[J]. 江苏农业科学,2016,44(10):103-107.
猜你喜欢
小学生(看图说画)(2020年10期)2020-10-29
实用口腔医学杂志(2017年6期)2017-09-19
中国实用医药(2016年30期)2016-12-28
农家科技中旬版(2016年12期)2016-04-16
哈尔滨医药(2015年2期)2015-12-01
安徽医药(2014年4期)2014-03-20
新疆农垦科技(2014年12期)2014-02-28
