
四川境内变电站RTV涂料绝缘子上藻类的分子系统学研究
2017-05-30 04:42周兵兵白欢李亚伟张志霞陈洪波张星海兰利琼卿人韦
安徽农业科学 2017年5期
周兵兵 白欢 李亚伟 张志霞 陈洪波 张星海 兰利琼 卿人韦
摘要[目的]对四川省境内变电站RTV涂料绝缘子表面藻类的生长情况进行了调研,并对其进行分子系统学分析。[方法]选取4个藻类生长比较典型的地点,对4地RTV涂料绝缘子上生长的藻类进行实地取样,用光学显微镜对其形态进行初步鉴定,并进一步使用18S rDNA基因进行系统进化分析。[结果]通过显微观察发现,4地优势藻类均为单细胞球状绿藻;系统进化分析发现,雅安藻种属于渡边球藻分支,与Heterochlorella luteoviridis亲缘关系较近,在MP、NJ和BI树中的支持率分别为86%、79%、1.0,将其初步鉴定为Heterochlorella sp.SCYA1;广安藻种属于Elliptochloris,与Elliptochloris subsphaerica的18S rDNA序列碱基没有差异,在MP、NJ和BI树中的支持率分别为97%、99% 和1.0,故确认其为Elliptochloris subsphaerica SCGA8藻种;巴中和南充藻种都属于栅藻属,两者亲缘关系很近且序列只有2个碱基差异,MP、NJ和BI树中的支持率比较高,分别为93%、 98% 和1.0,将其初步鉴定为Scenedesmus sp.SCBZ5和Scenedesmus sp.SCNC7。[结论]4地变电站RTV涂料绝缘子上的优势藻种不全是同一种藻,生长于四川省境内RTV涂料绝缘子上的藻类具有多样性。
关键词RTV涂料;藻种纯化;18S rDNA;系统进化分析
中图分类号S188文献标识码A文章编号0517-6611(2017)05-0135-05
Abstract[Objective]The algaes growth on the surface of the insulators coated with RTV (room temperature vulcanized silicone rubber)at Sichuan Province was investigated ,the molecular systematics were analyzed.[Method]Several typical substations were selected.Algaes collected from the surface of the insulators located in those areas were used to explore species diversity.Those dominant algaes were separately purified by spread plate method.The morphology was identified by optical microscopy and the phylogenetic analysis of four monoclonal algaes were carried out based on 18S rRNA sequences.[Result]Those algaes were singlecelled globular algaes examined by light microscopy.Phylogenetic analysis indicated that the Ya′an dominant algae was identified as Heterochlorella sp.SCYA1,which was closely related to Heterochlorella luteoviridis.The support rates of the two species were 86%,79%,1.0 in MP,NJ and BI trees,respectively.The 18S rDNA sequence of Guang′an dominant algae was not different from that of Elliptochloris subsphaerica ,the support rates of the two species in the MP,NJ and BI trees were 97%,99% and 10,respectively,it was identified as Elliptochloris subsphaerica SCGA8.Both Bazhong and Nanchong dominant algaes belonged to Scenedesmus,which had close genetic relationship,they were identified as Scenedesmus sp.SCBZ5 and Scenedesmus sp.SCNC7.The 18S rDNA sequence of two species only had two base differences,their support rates in the MP,NJ and BI trees were 93%,98% and 1.0.[Conclusion]Results indicate that four dominant algaes are not the same kind,algaes growing on the surface of RTV coated insulators at Sichuan Province have species diversity.
Key wordsRTV coating;Purification of algae species;18S rDNA;Phylogenetic analysis
基金項目四川省电力科学研究院项目(13H1138);四川省科技厅项目(2014JY0171)。
作者简介周兵兵(1990—),男,甘肃天水人,硕士研究生,研究方向:藻类分类及资源利用。*通讯作者,副教授,博士,从事藻类学及能源微藻油脂代谢研究。
收稿日期2016-12-19
室温硫化硅橡胶(Room Temperature Vulcanized Silicon Rubber,RTV)具有非常好的热稳定性、电绝缘性、憎水性和憎水迁移性[1],作为防污闪材料普遍应用于电力系统。在潮湿、污染比较严重的情况下,硅橡胶能够有效地防止污闪的发生[2]。
虽然硅橡胶作为防污闪材料有许多优点,但是近年来,很多地区都出现了生物生长在硅橡胶绝缘子表面的现象。藻类是硅橡胶绝缘子上生长的主要微生物,尤其是热带地区[3]。Fernando等[4]报道了斯里兰卡和坦桑尼亚33 kV输变电设施RTV绝缘子上藻类的生长,这些藻主要生长在绝缘子表面。从斯里兰卡硅橡胶绝缘子上分离得到的藻种经鉴定均为佐夫小球藻(Chlorella zonfingensis Dnz),这种藻主要分布在亚洲、澳洲以及欧洲,经常生长在树干、岩石以及建筑物墙壁上。从坦桑尼亚硅橡胶绝缘子上分离得到的绿藻被鉴定为Apatococcus constipatus,属于Chaetophraceae科,Leptosiroideae亚科,这种藻仅仅存在于非洲[5]。安置于新奥尔良西部沼泽的不同类型绝缘子上也发现有藻类和苔藓等生物的生长[6]。在德国,Kunde等[7]观察到使用7年之久的硅橡胶绝缘子上有生物的生长,这些生物由藻类和真菌组成,主要分布在绝缘子上较低的部位和绝缘子大小伞上表面边沿。日本Yonezawa试验点的聚合硅橡胶绝缘子经5年运行后,其上也发现有藻类的生长,这些藻存在于绝缘子相间间隔棒和套管上[8-9]。Sakata等[10]将绝缘子表面生长的藻鉴定为虚幻球藻(Apatococcus lobatus)。Dernfalk[11]在瑞典一个运行12年之久的11 kV配电系统的老化硅橡胶绝缘子上,也发现有严重的生物污染。这些硅橡胶绝缘子表面一部分被绿藻包被,一部分由地衣覆盖。被生物污染的绝缘子表面相对比较粗糙,并且藻类和地衣在绝缘子上大多呈片状分布[12]。目前,国外已经对绝缘子上生长的藻类做了很多研究工作,发现藻类的生长能影响绝缘子特性[13-16],可以导致污闪事故发生,进而影响电路的正常运行。
我国地域辽阔,自然环境丰富多样,但是藻类在我国输变电设施硅橡胶绝缘子上的生长情况鲜有报道。通过对四川省境内进行调研,发现藻类在硅橡胶绝缘子上生长具有普遍性。选取4个比较典型的地区:雅安市、广安市、巴中市和南充市,对4地变电站RTV涂料绝缘子、上生长的藻类进行采集、分离纯化,并对4地藻种的18S rDNA基因进行克隆和分子系统学研究,初步确立这4地藻種的分类地位并为后续的研究提供理论基础。
1材料与方法
1.1藻种及培养条件
2014年,藻种分别采自雅安市草坝220 kV变电站RTV涂料绝缘子、广安市铜堡220 kV变电站RTV涂料绝缘子、巴中市110 kV信义变电站RTV绝缘子和南充市营山县110 kV变电站RTV涂料绝缘子表面,采样时,选取4 cm×4 cm的样方,用药勺刮取藻层,收集在无菌离心管中,将从各地采集的藻样在灭菌后的BG11培养基[17]中培养,培养条件:温度22 ℃,光照强度3 000 lx,光暗周期12 h〖DK〗∶12 h。
1.2形态观察分别取30 μL各地藻液,制成水封片,在Olympus BH2显微镜下观察并用测微尺测量细胞大小,Toup View拍照。
1.3藻种的分离纯化取适量培养14 d左右的各地藻液,在血球计数板下计算细胞密度后将其稀释成不同梯度,用涂布平板法[18]对各地优势藻种进行纯化。
1.44地藻种18S rDNA 基因的克隆
4地藻种总DNA的提取采用CTAB法[19],用TU-1810紫外可见分光光度计(北京普析通用仪器有限责任公司)检测DNA浓度,-20 ℃保存待用。
用已知绿藻的18S rDNA基因序列,ClustalX 1.83[20]软件进行多序列比对(Multiple Alignment),选择保守区域,并用Primer Premier 5.0[21]软件设计引物,得到1对引物:18S5F(5-AGGCCTACCATGGTGGTAACGG-3); 18S7R(5-CCAGAACATCTAAGGGCATCAC-3)。对4地藻种18S rDNA进行扩增,PCR反应体系为25.00 μL,包含2.50 μL 10×Buffer,0.15 μL Taq DNA聚合酶(Vazyme,南京),2.50 μL dNTP(2.5 mmol/L),上下游引物各1.00 μL,2.00 μL DNA模板约50 ng,15.85 μL双蒸水。PCR反应程序为94 ℃预变性5 min;之后进行34个循环,每个循环包括94 ℃变性30 s,49 ℃退火30 s,72 ℃延伸90 s;最后72 ℃总延伸10 min。PCR产物用SanPrep柱式胶回收试剂盒(Sangon,上海)纯化后,连接到pMD19-T载体(TaKaRa,大连有限公司)。连接产物转化到E.coli DH5α(Invitrogen,USA)感受态细胞中,经菌落PCR鉴定阳性克隆,所获阳性克隆送华大基因技术有限公司测序。
1.54地藻种18S rDNA基因的分子系统学分析
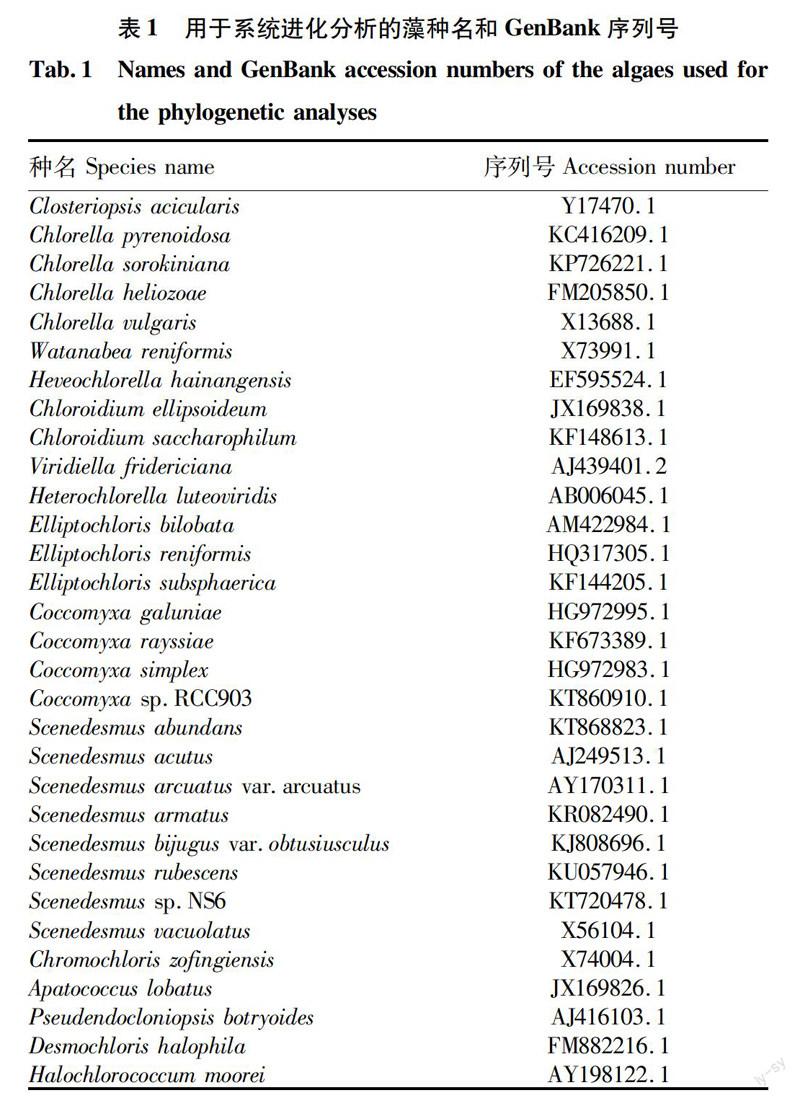
在GenBank数据库中下载一系列相关绿藻的18S rDNA基因序列(表1)。将从GenBank数据库中获得的已知绿藻的18S rDNA基因序列与克隆得到的4地藻种的18S rDNA基因序列一起构建分析数据库,用ClustalX 1.83[20]对所构建的基因数据中的序列进行对位排列和多序列比对,在此基础上进行相关的分子系统学分析。
45卷5期周兵兵等四川境内变电站RTV涂料绝缘子上藻类的分子系统学研究
1.5.1贝叶斯 (Bayesian Inference,BI)系统分析法。
运用 MrBayes v3.1.2[22]软件,首先对数据进行模型和参数估计,以赤池信息量准则(Akaike Infomation Criterion,AIC )[23]选择出最佳碱基替代模型。2个并行程序分别以4条链(Markov Chain Monte Carlo,MCMC )运行20 000 000代,每100代保留1棵树。所得的样本树每1 000代的前20%作为老化样本舍弃,剩余样本树采用50%多数一致性原则构建一致树。最后通过后验概率(Posterior Probability,PP)评估进化树拓扑结构的可靠性。
1.5.2邻位相接 (Neighbor-Joining,NJ) 系统分析法。
运用MEGA v.6.0[24]软件,采用Kimura-2-parameter参数遗传距离模型[25],对系统发育树的拓扑结构通过自展法(Bootstrap,BS)进行置信度检测,自检数据集为1 000。
1.5.3最大简约 (Maximum Parsimony,MP) 系统分析法。
分析使用 PAUP* version 4.0 beta 10软件[26],以树二重连接(Tree Bisectionreconnection)进行枝长变换;启发式搜索(Heuristic Searches)进行1 000次随机添加(Random Addition)重复;众树选择(MulTrees Option) 获取分支树。同样,通过自展法对进化树拓扑结构进行置信度检测,自检数据集为1 000。
以上方法均以Pseudendocloniopsis botryoides、Desmochloris halophila和Halochlorococcum moorei作为外类群。
2结果与分析
2.14地RTV涂料绝缘子上藻类的生长情况
从图1可看出,在4地各变电站RTV涂料绝缘子大、小伞的上表面均有藻类生长,藻类主要生长在未被小伞遮挡的大伞外表面,除雨水径流过的地方(图1B,箭头所指)外,藻类的分布都较均匀;绝缘子小伞上藻类的生长较少,主要分布于小伞上表面外边沿,且分布不均,呈零星片状分布。在背光,水和尘埃不易附着的绝缘子大、小伞下表面均无藻类生长。从上往下,藻类在绝缘子表面的生长量逐渐增多。
2.24地RTV绝缘子上藻类的显微观察
该试验分离的4地优势藻种依采样点分别命名为SCYA1(雅安藻种)、SCGA8(广安藻种)、SCBZ5(巴中藻种)和SCNC7(南充藻种)。
SCYA1(图2A)为单细胞球状绿藻,细胞大小為4.0~90 μm; 成熟细胞叶绿体杯状,1或2个,幼体细胞叶绿体大多纺锤形,无明显蛋白核;无鞭毛,液体环境下不能运动。SCYA1形态学特征与异小球藻属(Heterocholorella)较为相似。
SCGA8(图2B)为单细胞,有时许多细胞暂时集聚在一起,细胞球形,细胞大小为4.5~13.0 μm;叶绿体轴生、星芒状,充满整个细胞,无明显蛋白核;无鞭毛,液体环境下不能运动;营养繁殖为细胞分裂,分裂面多少与其细胞长轴垂直。SCGA8形态学特征与共球藻纲的Elliptochloris藻种类似。
SCBZ5(图2C)和SCNC7(图2D)均为单细胞球形,细胞大小为5.5~11.0 μm和5.0~11.0 μm;叶绿体片状、周生,1个,具有1个蛋白核,细胞壁平滑、无刺;无鞭毛,液体环境下不能运动;在无性生殖过程中每个细胞产生2个椭球状似亲孢子,待母细胞壁破裂后子细胞被释放出来,没有观察到有性生殖。SCBZ5、SCNC7形态学特征与栅藻属(Scenedesmus)中球形藻种较为相似。
由于在光学显微镜下难以观察到更细致的特征,因此仅依据形态学特征不能判断以上藻种在分类学上的属种位置。
2.3序列分析
通过PCR扩增获得4地藻种18S rDNA基因序列长度为1 078 bp。与表1中各藻种18S rDNA基因多序列比对后,最终比对序列总长度为1 031 bp。在18S rDNA基因序列矩阵中,包含213个变异位点,163个简约信息位点,分别占总位点的20.66%和15.81%。碱基含量差异不大,碱基T、C、A和G的平均含量分别为25.1%、20.6%、25.1%和29.2%。G+C 的含量(49.8%)和A+T 的含量(50.2%)基本相当。所测序藻种SCYA1的18S rDNA基因序列与黄绿异小球藻(Heterochlorella luteoviridis)、喜糖椭圆球藻(Chloroidium saccharophilum)、椭圆球藻(Chloroidium ellipsoideum)、肾形渡边球藻(Watanabea reniformis)和弗雷德鲜绿球藻(Viridiella fridericiana)相同基因序列之间的相似性分别达96.29%、94.81%、94.63%、94.25% 和93.80%;藻种SCGA8的18S rDNA基因序列与Elliptochloris subsphaerica、Elliptochloris reniformis和Elliptochloris bilobata相同基因序列之间的相似性分别达100%、 99.32% 和99.16%;藻种SCBZ5和SCNC7的18S rDNA基因序列之间相似度达99.81%,只有2个碱基的差异,与Scenedesmus vacuolatus、Scenedesmus bijugus var.obtusiusculus、Scenedesmus rubescens和Scenedesmus acutus相同基因序列之间的相似性分别达99.63% 和9944%、99.07%和98.88%、98.88%和98.70%、98.70%和98.51%。这一高度的相似性进一步支持了形态学鉴定的结果。
2.4系统发育分析
用MP、NJ和BI法构建的系统树的拓扑结构基本一致,如图3所示。雅安变电站RTV绝缘子上的藻种在系统树中位于渡边球藻属(Watanabea)分支,与黄绿异小球藻(Heterochlorella luteoviridis)聚为一簇,MP、NJ和BI树中的支持率较高,分别为86%、79%、1.0,可以确定其为异小球藻属的藻种,并初步鉴定为Heterochlorella sp.SCYA1;广安RTV绝缘子上的藻种位于Elliptochloris属分支, 与Elliptochloris subsphaerica碱基没有差异,MP、NJ和BI树中的支持率分别为97%、99% 和1.0,可以确认其为Elliptochloris subsphaerica SCGA8藻种。巴中和南充变电站RTV涂料绝缘子上藻在系统树中都位于栅藻属分支,且两者亲缘关系很近,MP、NJ和BI树中的支持率比较高,分别为93%、98%和1.0。可以将这2种藻确认为栅藻属藻种,将其分别鉴定为Scenedesmus sp.SCBZ5和Scenedesmus sp.SCNC7。
3结论与讨论
该研究先采用传统方法进行分离鉴定,再用现代分子生物学方法对4个藻种进行分析,这较大程度地保证了结果的准确性和客观性。通过显微观察发现,雅安市草坝220 kV变电站、广安市铜堡220 kV变电站、南充市营山县110 kV变电站和巴中市110 kV信义变电站RTV涂料绝缘子上的优势藻种形态特征虽有不同,但是在光学显微镜下很难确定这几种藻分类学上的种属位置。通过18S rDNA基因构建的系统发育树发现,这几种藻并非同一种藻,分属不同类别。Heterochlorella sp.SCYA1为共球藻纲、渡边球藻分支、异小球藻属藻种;Elliptochloris subsphaerica SCGA8为共球藻纲Microthamniales目Elliptochloris属藻种,而Scenedesmus sp.SCBZ5和Scenedesmus sp.SCNC7均属于绿藻纲、环藻目、栅藻属。
国外报道生长在硅橡胶绝缘子上的藻种有Chlorella zonfingensis Dnz、Apatococcus constipatus、Apatococcus lobatus,除了Apatococcus constipatus藻种的18S rDNA基因序列没办法获取外,将其他2种藻的18S rDNA基因序列同样用于系统进化树的构建并判断这2种藻是否与该研究中的藻种分类地位一致。经发现Chlorella zonfingensis Dnz藻种并非与Gubanski等[5]科学家鉴定的那样,是小球藻科成员,而是该藻位于绿藻纲,与栅藻属亲缘关系相近(BI、NJ和MP树中支持率分别为1.0、99%和99%)。近来年,Fucíkov等[27]根据该藻的分子和形态两方面特征,重新定义了该藻,将其归为环藻目、色球藻属。这与该研究构建的系统进化树中该藻种的分类地位相吻合。Sakata等[10]将Apatococcus lobatus鉴定为共球藻纲、小球藻科、虚幻球藻属成员。在构建的系统发育树中发现,该藻是共球藻纲成员,但并非小球藻科成员,独立于小球藻科外,自成分支。由此可以看出,能够在硅橡胶绝缘子上生长的藻并非一种,而是不同地方藻的种类有所不同,在我国四川境内各变电站绝缘子上生长的藻也不尽相同,这说明硅橡胶绝缘子上生长的藻种具有物种多样性。
藻类是光能自养型生物,能够利用CO2作为碳源合成有机物供藻细胞生长与繁殖。同时,光照、温度、盐度和水分等也是藻类生长和繁殖过程中不可缺少的生态因子。从已报道的藻类生长在硅橡胶绝缘子表面的地区来看,绝大多数处于热带亚热带地区[3-12],而调查的雅安、广安、南充和巴中4地都处于亚热带季风性湿润气候区,全年气候适宜且极端天气较少[28-31]。这几个地方的光照和温度都能满足藻类在野生环境下生长,而空气湿度成了绝缘子上藻类生长的主要限制因素。有机硅橡胶的表面张力非常小,水滴在材料表面形成的静态接触角较大[32-34],喷淋试验显示:水在RTV硅橡胶涂料表面形成分离的水珠,而不是连续的水膜,这使得水在RTV涂料表面形成点状聚集,减少了水分蒸发的面积,可以为藻类的生长提供一个潮湿环境,并为其提供生长所需的无机盐类。与陶瓷或玻璃材料相比,RTV硅橡胶导热系数更低,具有更好的绝热性[35-37]。外界温度的变化和太阳光持续的照射都不易使RTV硅橡胶表面温度发生急剧变化,这可减缓绝缘子表面水分的蒸发,并为藻类提供一个相对适宜的生长环境。从藻类在4地RTV涂料绝缘子上的生长及分布情况可以看出,在背光、水分不易聚集的绝缘子大、小伞下表面没有藻类的着生,被小伞遮挡的大伞上表面几乎也没有藻类的生长,这说明光合作用是藻类生长的基础,而藻类受到光照强弱和接触水分的多少对其光合作用效率影响很大。除此之外,藻类生命力非常强,可在逆境环境下通过自身代谢或者产生孢子等度过不良环境,而在条件适宜时可恢复生长,并快速繁殖形成新的藻落。综合以上因素可以推断出,藻类生长在野外RTV绝缘子表面是一种必然结果,且藻类在其上面的生长情况要比陶瓷或玻璃絕缘子上更好。藻类生长对绝缘子的使用寿命、电化学性质会产生影响,其产生的机理和影响的程度还需进一步探索。
综上,该研究初步确定了四川境内雅安、广安、巴中和南充变电站RTV涂料绝缘子上藻类的分类地位,这丰富了硅橡胶表面生长藻类的物种多样性,也为探究我国其他地方绝缘子上藻类的生长与硅橡胶和藻类的生长关系提供一定的理论基础。
参考文献
[1] 陈洪波,钟聪,何松.浅析RTV防污闪涂料在电力系统的应用[J].四川电力技术,2003,26(1):39-40.
[2] KUMAGAI S.Influence of algal fouling on hydrophobicity and leakage current on silicone rubber[J].IEEE Transactions on Dielectrics & Electrical Insulation,2007,14(5):1201-1206.
[3] CALLOW M E,CALLOW J A.Enhanced adhesion and chemoattraction of zoospores of the fouling alga enteromorpha to some foulrelease silicone elastomers[J].Biofouling,1998,13(2):157-172.
[4] FERNANDO M A R M,GUBANSKI S M.Performance of nonceramic insulators under tropical field conditions[J].IEEE Transactions on Power Delivery,2000,15(1):355-360.
[5] GUBANSKI S M,FERNANDO M A R M,PIETR S J,et al.Effects of biological contamination on insulator performance[C]∥Properties and applications of dielectric materials,2000.Proceedings of the 6th international conference on.Goteborg:Chalmers Univ.of Technol.,2000:797-801.
[6] MCAFEE R D,HEATON R D,KING J M,et al.A study of biological contaminants on high voltage porcelain insulators,I[J].Electric power systems research,1997,42(1):35-39.
[7] KUNDE K,HENNING S R,KUHL M R,et al.New experience with composite insulators[R].Presented at Cigre,1998.
[8] JOSHO M,MIYAKAWA H,SAKATA M,et al.Long term field test of polymer insulators at the Yonezawa Test Site[C]∥IEEJ Transactions on HV.[s.l]:[s.n],2001.
[9] Japanese utility studies composite insulators at longterm test station[R].In Insulator News and Market Report,2004:26-32.〖ZK)〗
[10] 〖ZK(#〗SAKATA M,MIYAKAWA H,UTSUMIAND Y,et al.Investigation of algae growth on surface of polymer insulators and hollow core insulators at the Yonezawa test station[C]∥Annual Meeting Record of IEE Japan.Japanese:[s.n.],2001.
[11] DERNFALK A.Diagnostic methods for composite insulators with biological growth[M].Gteborg:Chalmers University of Technology,2004.
[12] WALLSTR M S,KARLSSON S.Biofilms on silicone rubber insulators; microbial composition and diagnostics of removal by use of ESEM/EDS:Composition of biofilms infecting silicone rubber insulators[J].Polymer degradation and stability,2004,85(2):841-846.
[13] FLEMMING H C.Relevance of biofilms for the biodeterioration of surfaces of polymeric materials[J].Polymer degradation & stability,1998,59(1/2/3):309-315.
[14] SUNDARARAJAN R,GRAVES J,MOHAMMED A,et al.Performance evaluation of polymeric insulators aged in lab and field[C]∥Power engineering society general meeting,2003,IEEE.[s.l.]:IEEE,2003.
[15] FERNANDO M A R M,GUBANSKI S M.Performance of nonceramic insulators under tropical field conditions[J].IEEE Transactions on Power Delivery,2000,15(1):355-360.
[16] GAUTAM B K,MIZUNO Y,MATSUBAYASHI G,et al.Effect of wetting conditions on the contamination flashover voltages of polymer insulators[C]∥International conference on properties and applications of Dielectric Materials.[s.l.]:IEEE,2006:538-541.
[17] ALLEN M M,STANIER R Y.Growth and division of some unicellular bluegreen algae[J].Journal of general microbiology,1968,51:199-202.
[18] BUCK J D,CLEVERDON R C.The spread plate as a method for the enumeration of marine bacteria[J].Limnology & oceanography,1960,5(1):78-80.
[19] ROGERS S O,BENDICH A J.Extraction of total cellular DNA from plants,algae and fungi[M].The Netherlands:Kluwer Academic Publishers,1994.
[20] JEANMOUGIN F,THOMPSON J D,GOUY M,et al.Multiple sequence alignment with CLUSTAL X[J].Trends in biochemical sciences,1998,23(10):403-405.
[21] LALITHA S.Primer premier 5[J].Biotech software & internet report,2000,1(6):270-272.
[22] RONQUIST F,HUELSENBECK J P.MrBayes 3:Bayesian phylogenetic inference under mixed models [J].Bioinformatics,2003,19(12):1572-1574.
[23]AKAIKE H.A new look at the statistical model identification[J].Automatic Control IEEE Transactions on,1974,19(6):716-723.
[24] TAMURA K,STECHER G,PETERSON D,et al.MEGA6:Molecular evolutionary genetics analysis version 6.0[J].Molecular biology and evolution,2013,30(12):2725-2729.
[25] SRIVATHSAN A,MEIER R.On the inappropriate use of Kimura2parameter (K2P) divergences in the DNAbarcoding literature[J].Cladistics,2012,28(2):190-194.
[26] SIMONSEN T J,BROWN R L,SPERLING F A H.Tracing an invasion:Phylogeography of Cactoblastis cactorum(Lepidoptera:Pyralidae) in the United States Based on Mitochondrial DNA[J].Annals of the entomological society of America,2008,101(5):899-905.
[27] FUC〖DD(-1〗ˇ〖DD)〗KOV K,LEWIS L A.Intersection of Chlorella,Muriella and Bracteacoccus:Resurrecting the genus Chromochloris KOL et CHODAT(Chlorophyceae,Chlorophyta)[J].Fottea,2012,12(1):83-93.
[28] 吳泽,范广洲,周定文,等.近60年“雅安天漏”变化特征分析[J].成都信息工程学院学报,2012,27(1):63-70.
[29] 黎红,熊华萍,李鹏,等.广安市局地气候对城区空气质量影响研究[J].四川环境,2015,34(4):97-104.
[30] 鲜铁军,翟园.1961~2014年南充市气温变化特征分析[J].安徽农业科学,2015,43(25):213-214,217.
[31] 张雷,肖天贵,燕亚菲,等.四川巴中地区38年来气候变化特征分析[J].成都信息工程学院学报,2010,25(3):293-300.
[32] 苏建军,李杰,王学刚,等.RTV硅橡胶防污闪涂料研究现状[J].山东电力技术,2015,42(1):1-7.
[33] KIM S H,CHERNEY E A,HACKAM R.Suppression mechanism of leakage current on RTV coated porcelain and silicone rubber insulators[J].IEEE Transactions on Power Delivery,1991,6(4):1549-1556.
[34] 吉翠萍,管延锦,张筱,等.室温硫化硅橡胶防污闪涂料的研究进展[J].涂料工业,2015,45(11):81-87.
[35] 杨栋,张炜,姜本正.硅橡胶基绝热材料高温热行为研究[J].固体火箭技术,2012,35(3):396-400.
[36] 潘大海,刘梅,孟岩,等.导热绝缘室温硫化硅橡胶的研制[J].橡胶工业,2004,51(9):534-536.
[37] 韩雄炜.导热硅橡胶的制备及性能[D].杭州:浙江大学,2006.
猜你喜欢
可再生能源(2022年8期)2022-08-17
大自然探索(2019年2期)2019-03-01
中国测试(2018年10期)2018-11-17
生态学报(2018年14期)2018-08-29
农业工程学报(2017年3期)2017-03-04
分析测试学报(2015年9期)2015-12-17
橡胶工业(2015年8期)2015-07-29
橡胶工业(2015年6期)2015-07-29
真空与低温(2015年4期)2015-06-18
山西建筑(2015年17期)2015-06-05
